
Anti-Listerial Activity of Bacteriocin-like Inhibitory Substance Produced by Enterococcus lactis LBM BT2 Using Alternative Medium with Sugarcane Molasses


 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
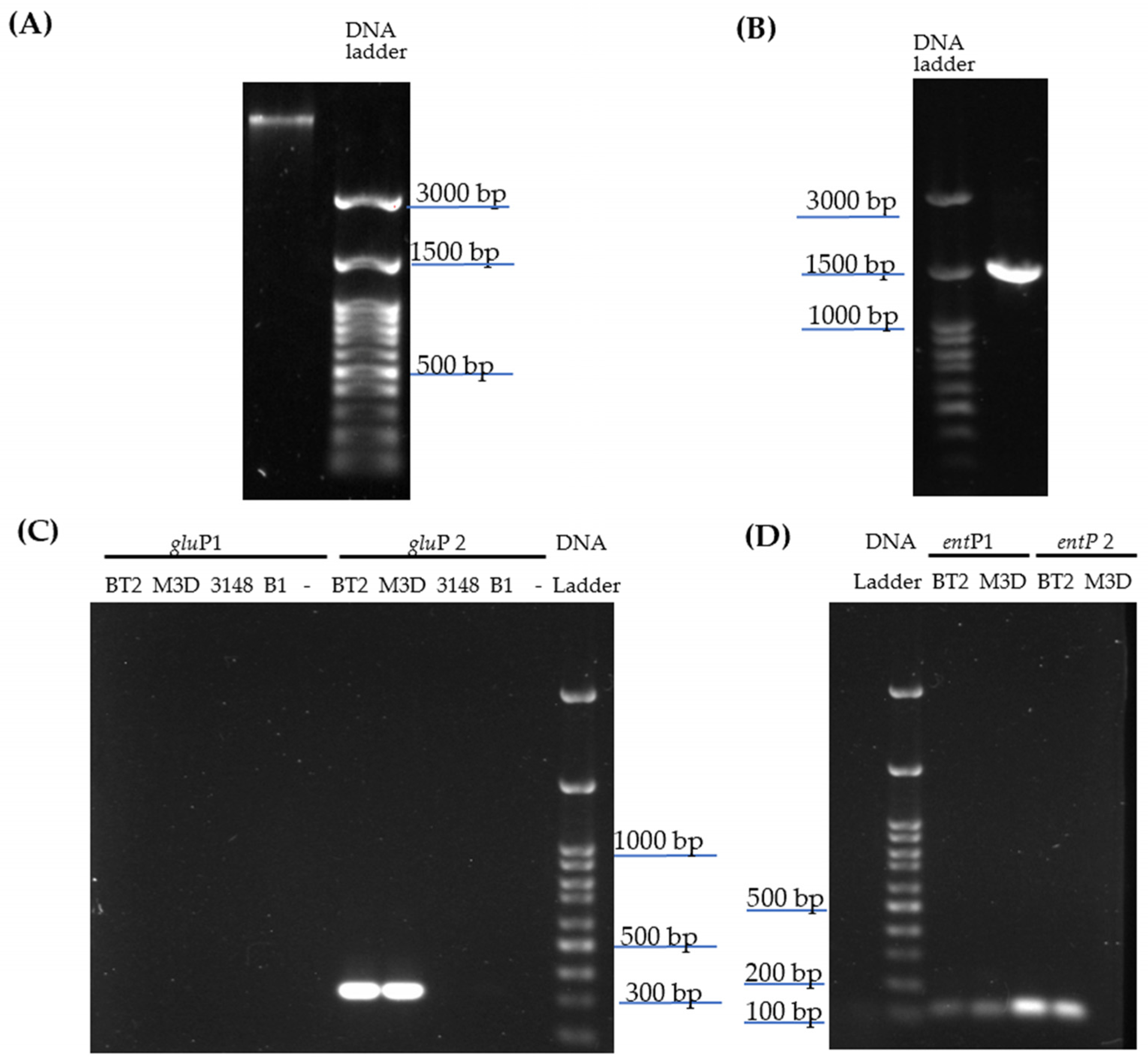
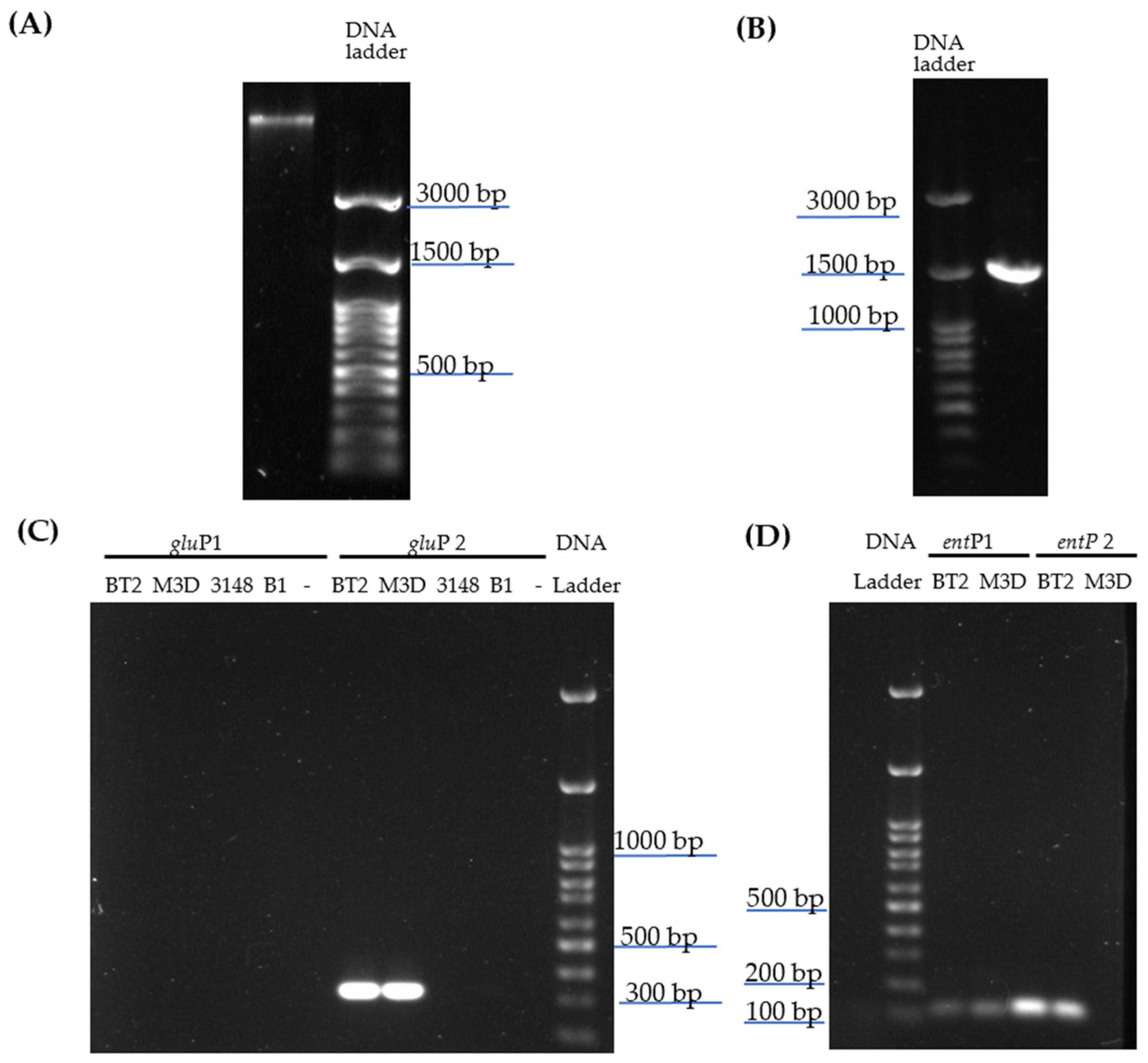
2.1. Bacterial Identification and Screening of Enterocin-Encoding Genes by PCR
2.2. Antibacterial Screening of Bacteriocin-like Inhibitory Substances from E. lactis LBM BT2
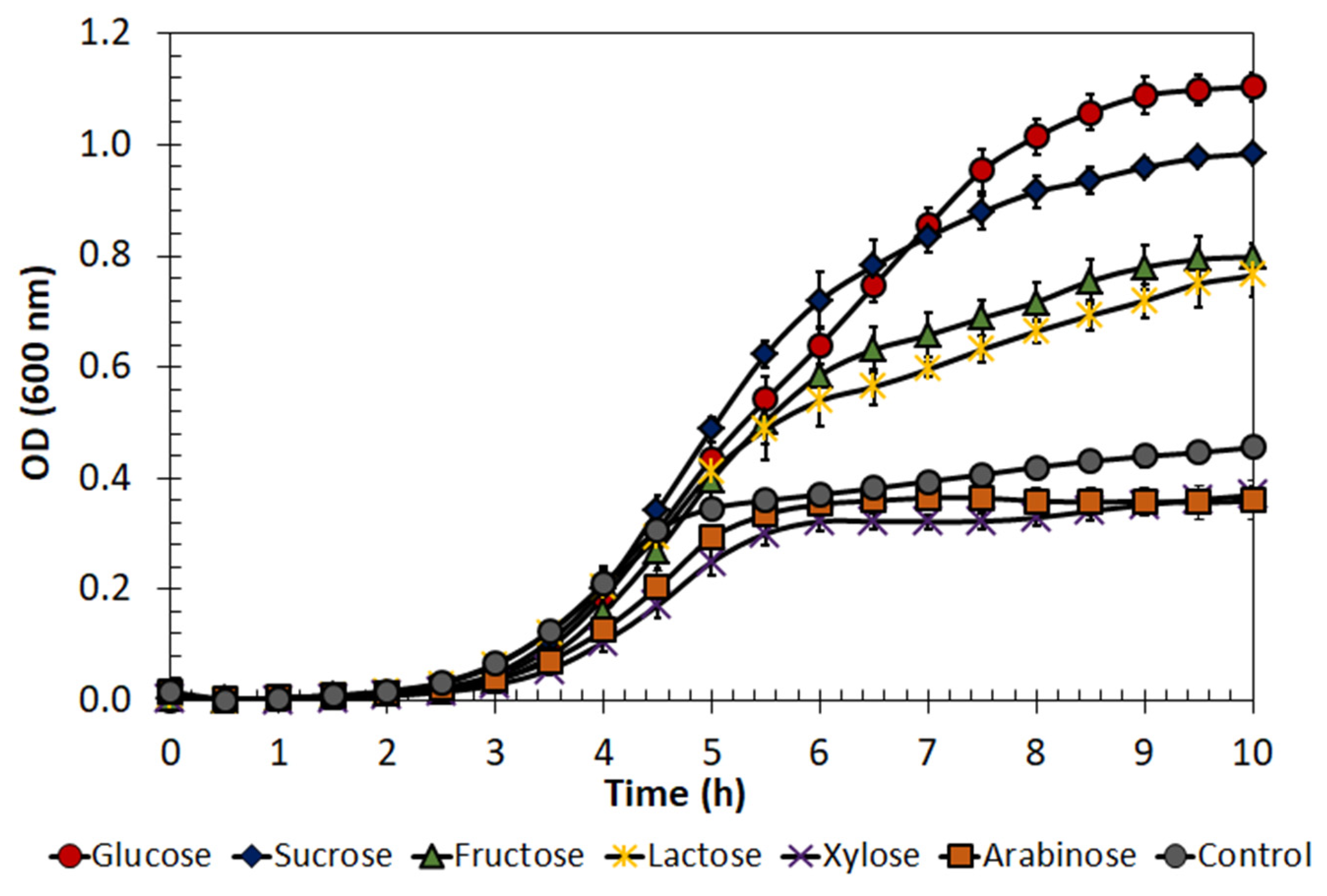
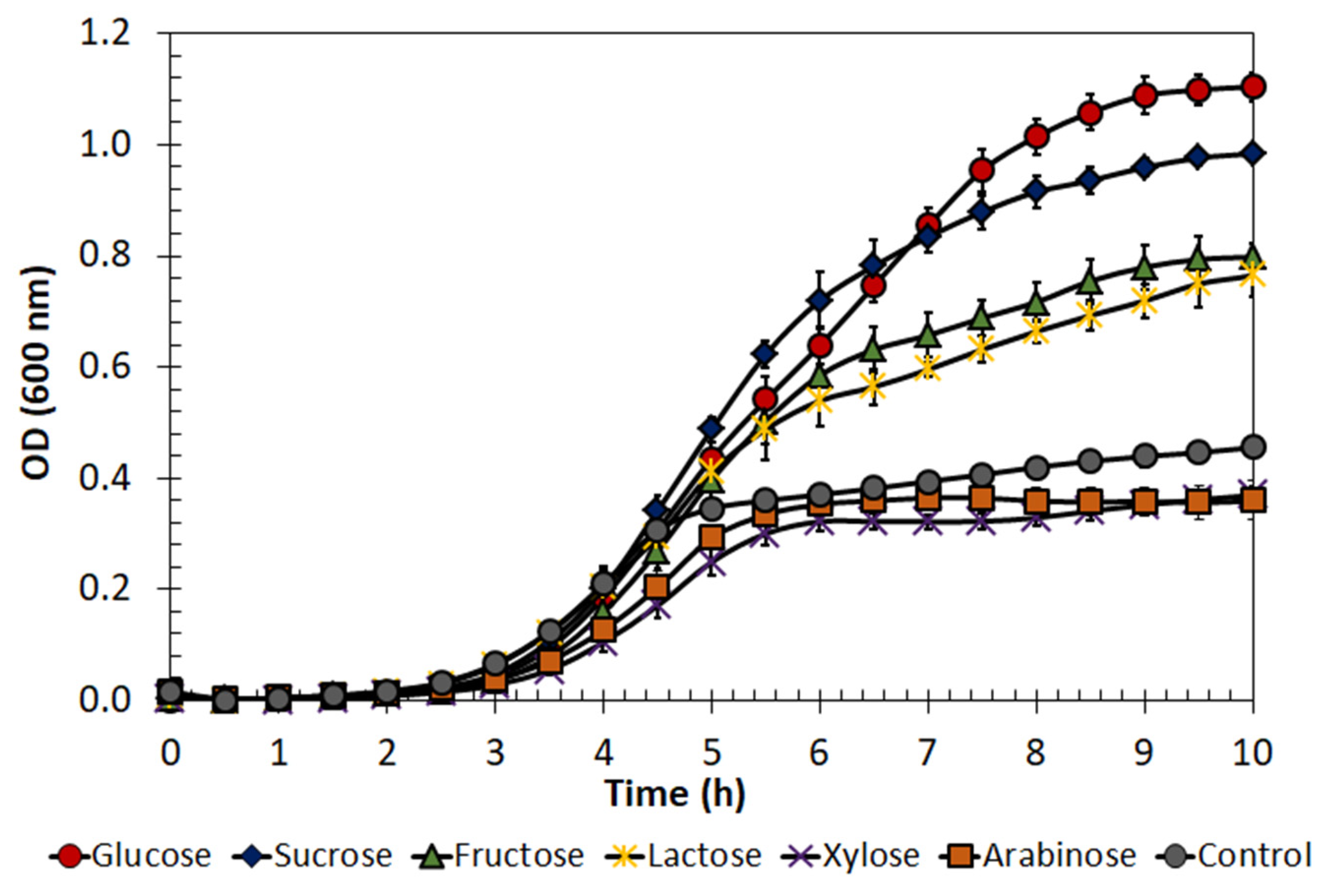
2.3. Enterococcus lactis Growth on Different Substrates
2.4. Molasses Composition
2.5. Effect of Different Molasses Concentration on E. lactis Growth
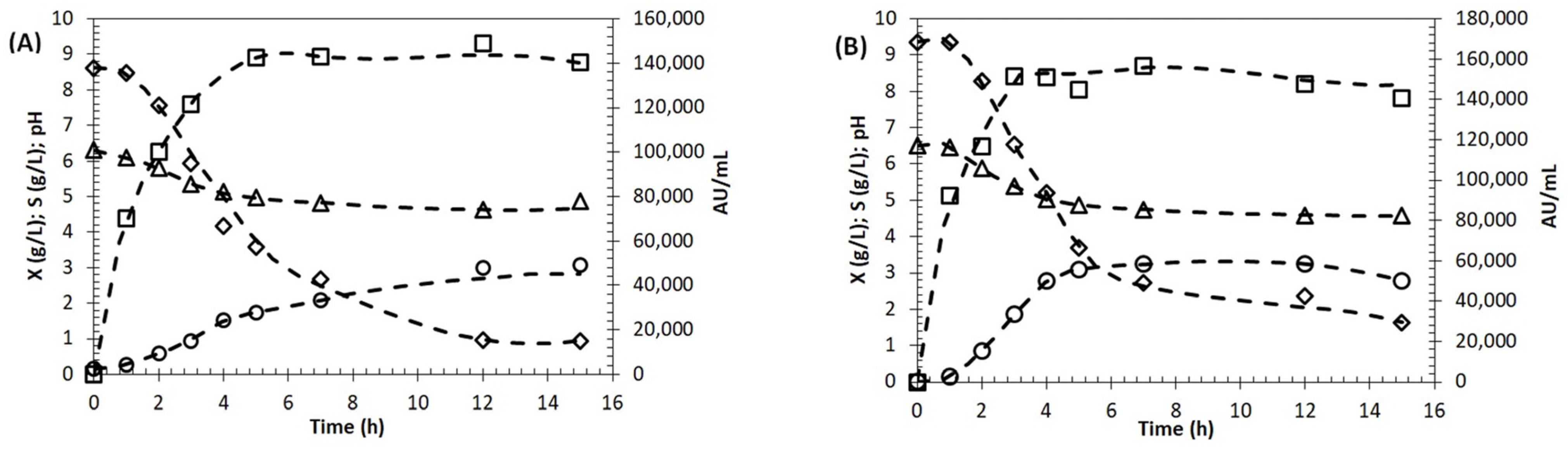
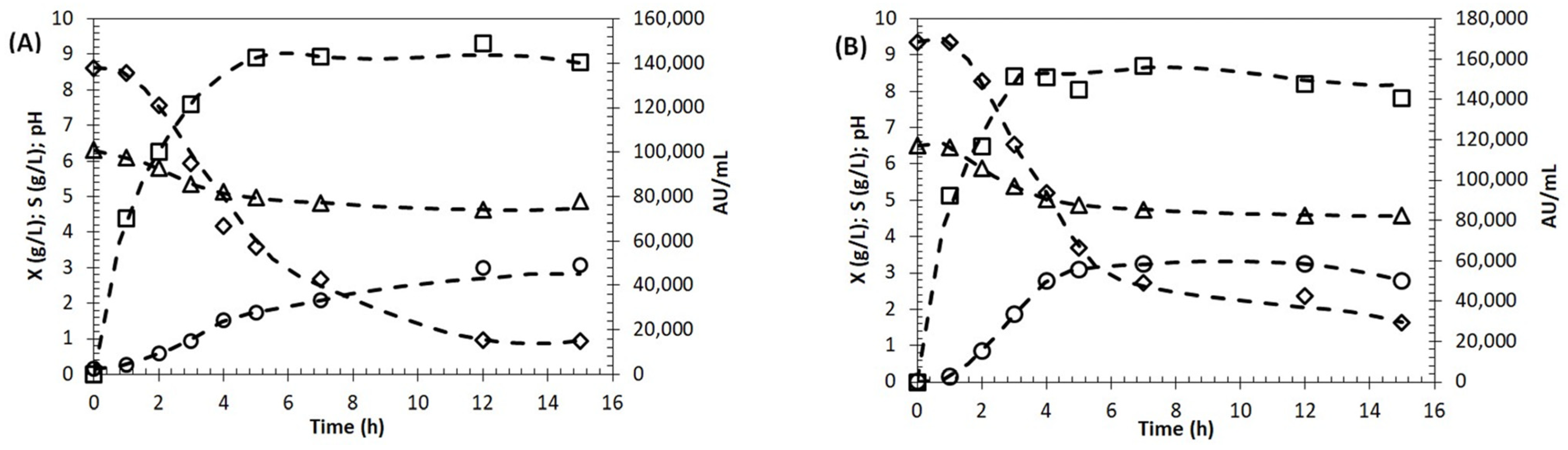
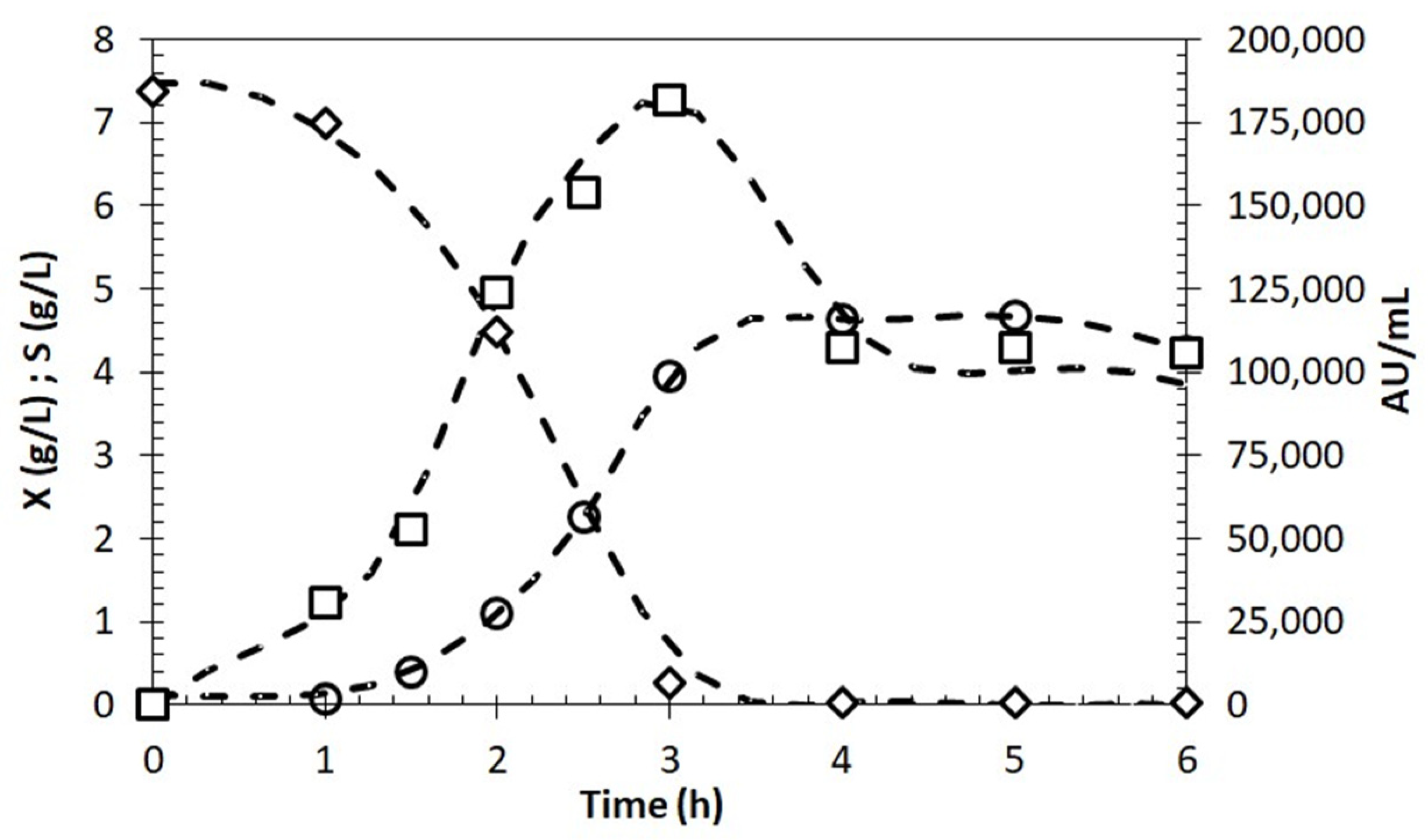
2.6. Growth and BLIS Production by Enterococcus lactis in Molasses as Carbon Source
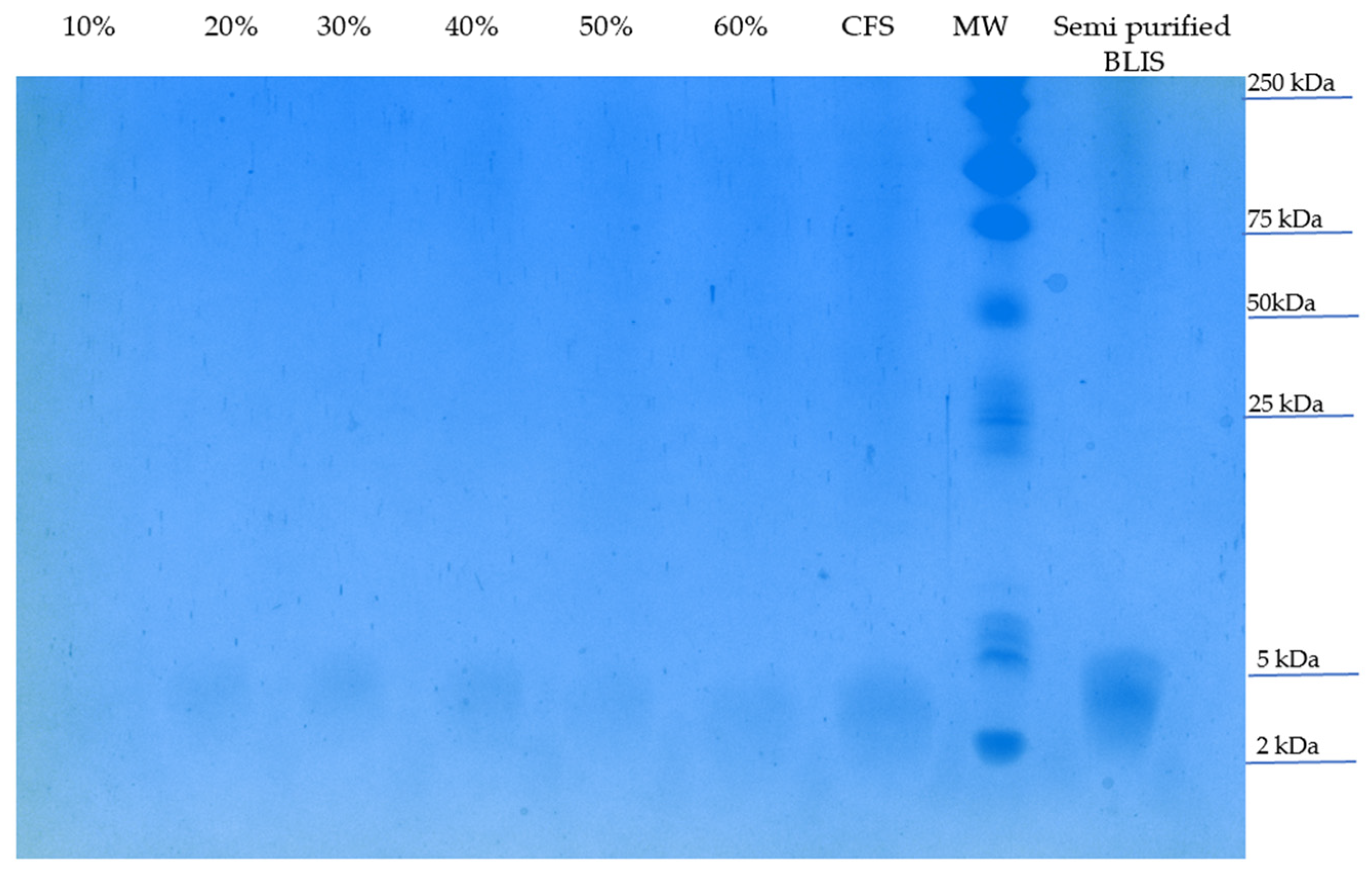
2.7. Partial Purification and Characterization of BLIS
2.8. Stability of Semi-Purified BLIS to pH, Temperature and Enzymatic Hydrolysis
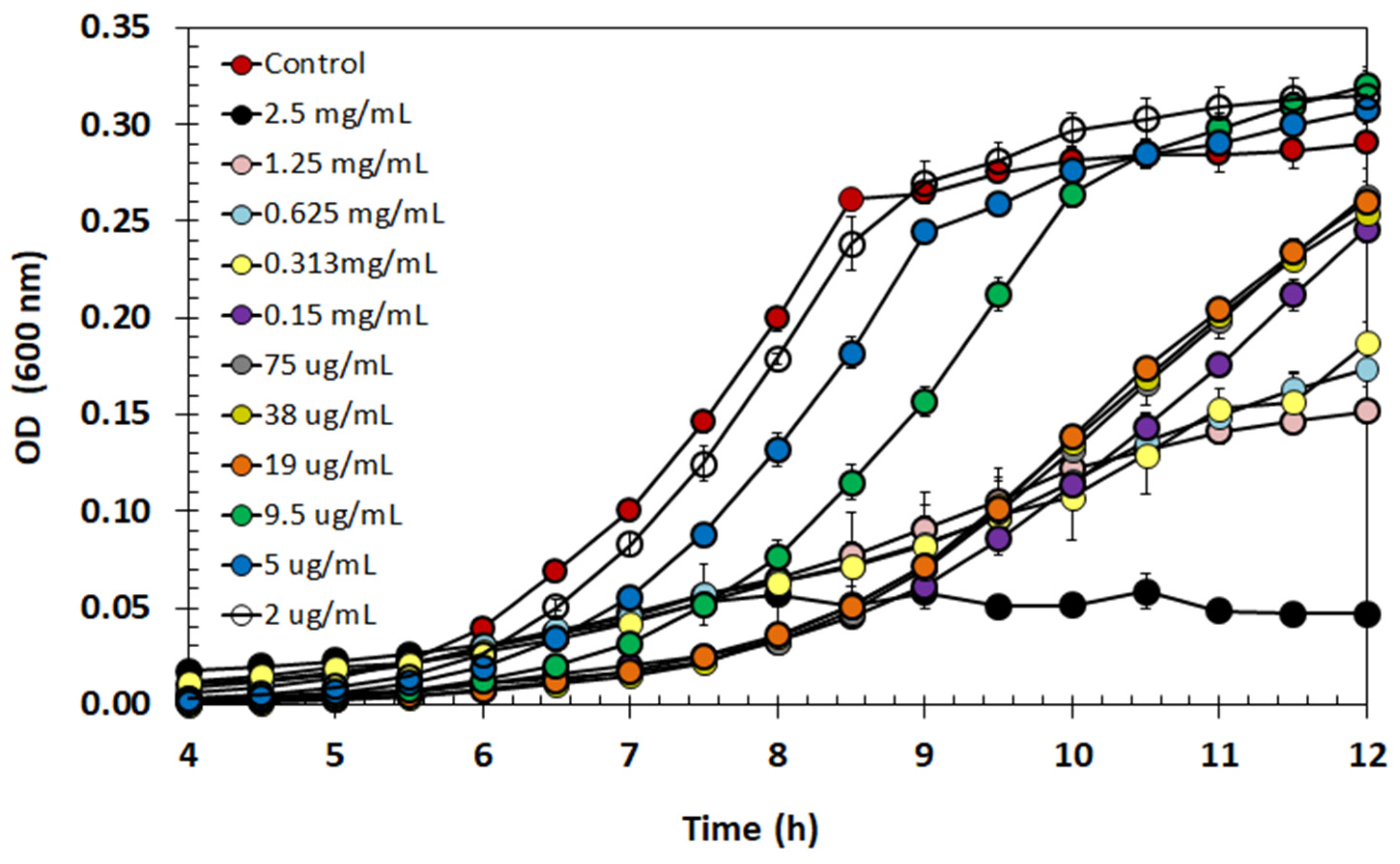
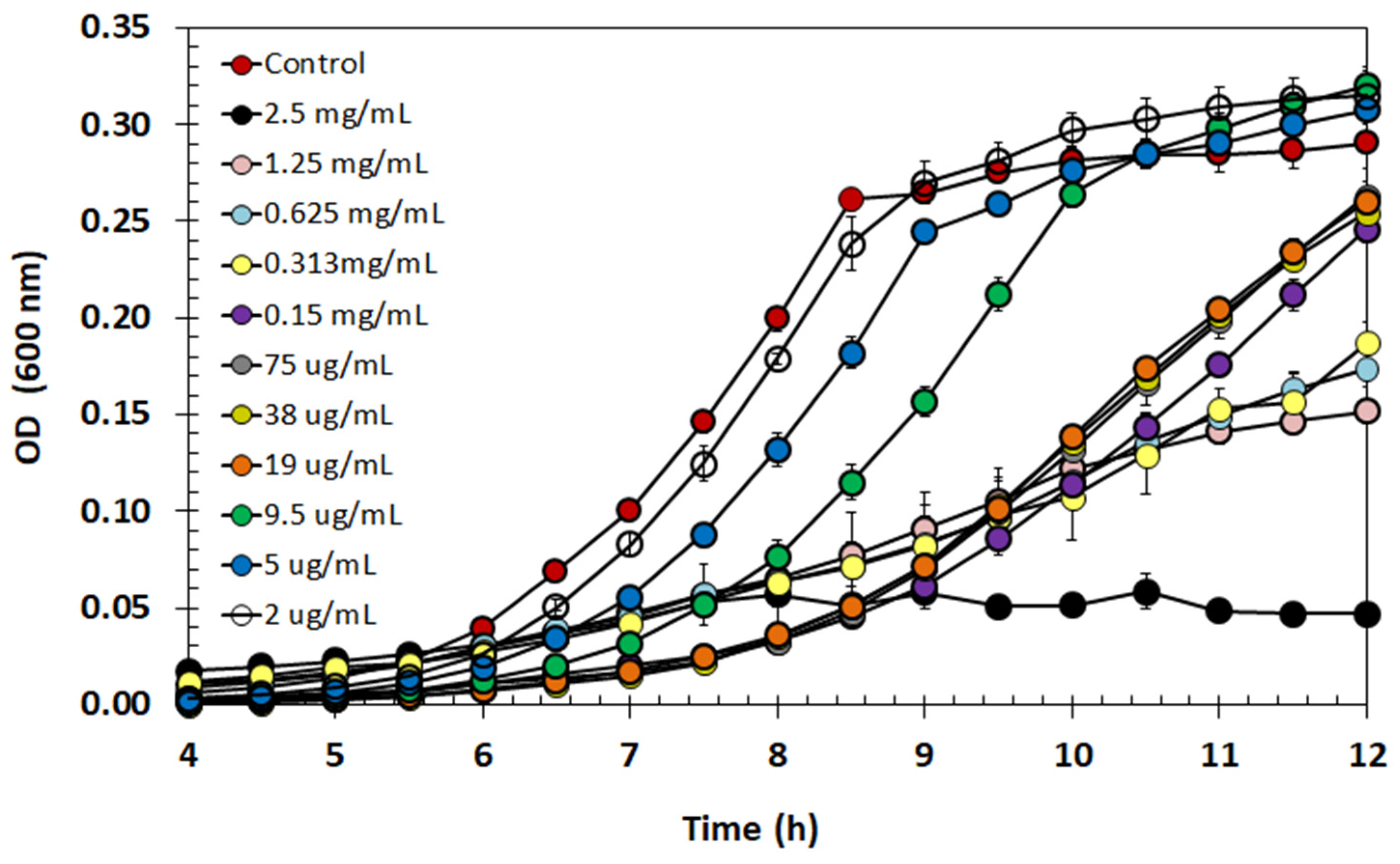
2.9. Effect of Semi-Purified BLIS on Listeria monocytogenes Growth
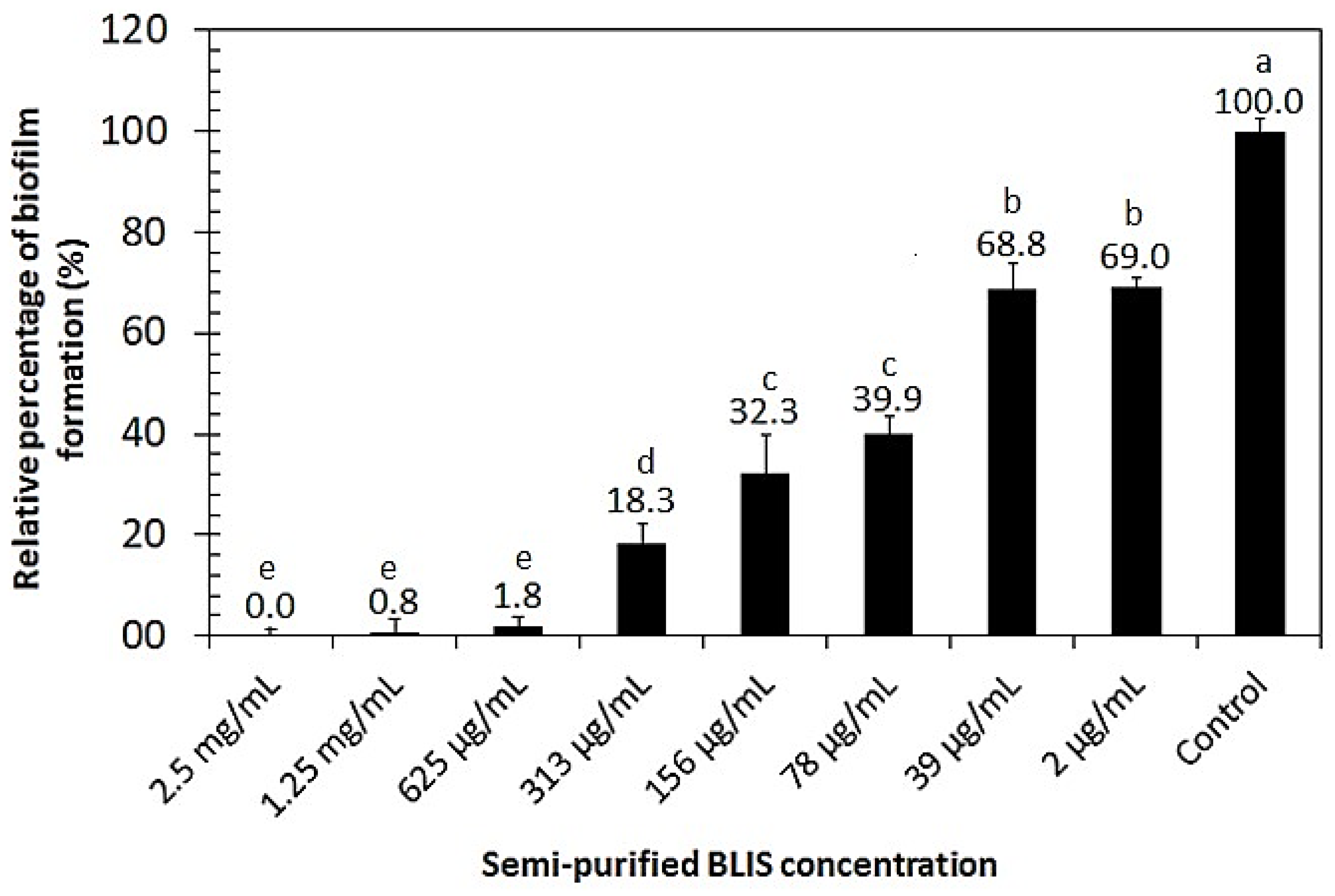
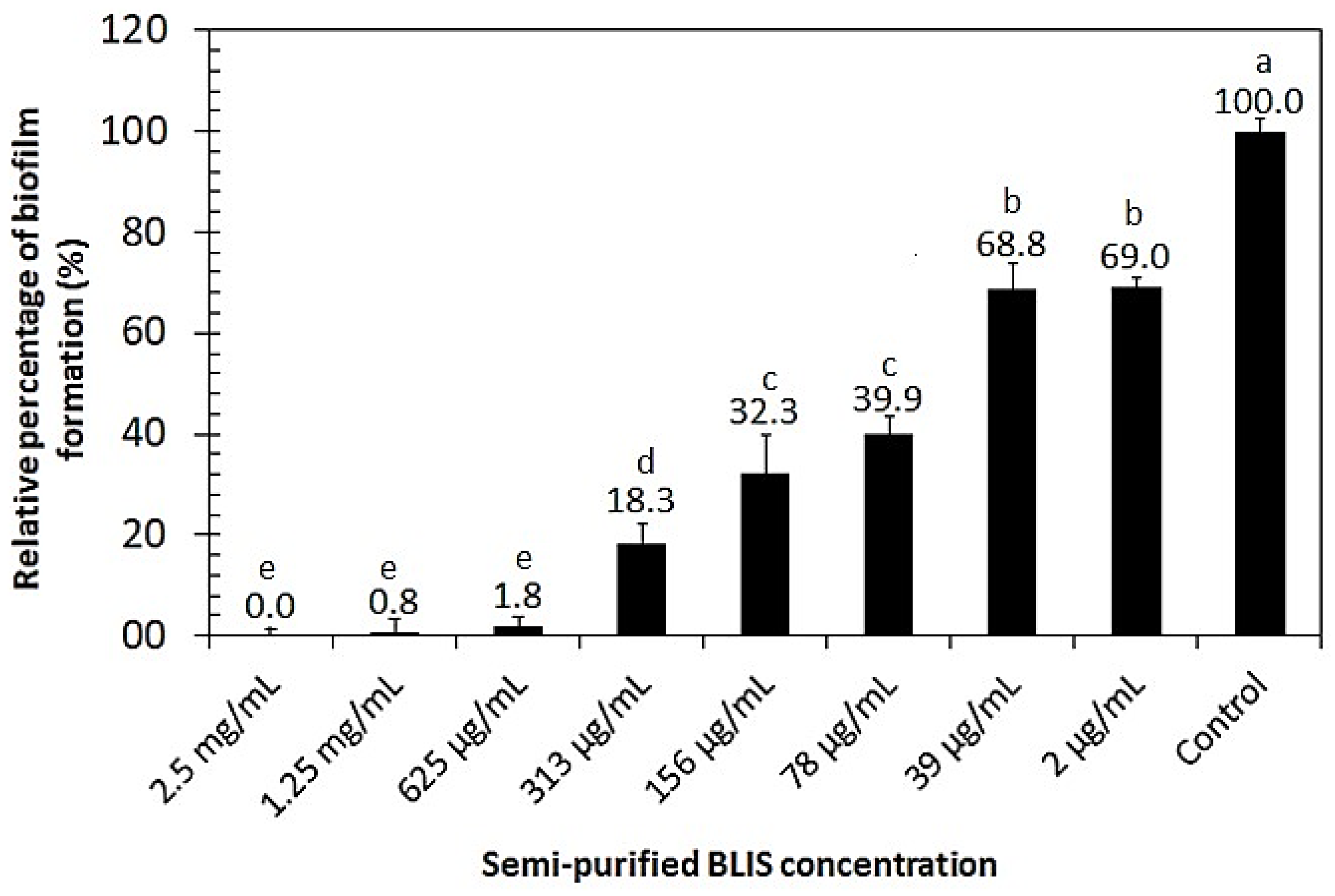
2.10. Biofilm Inhibition Activity of Semi-Purified BLIS
3. Discussion
3.1. Bacterial Identification and BLIS Characterization
3.2. BLIS Production by Enterococcus lactis LBM BT2 in Molasses-Based Medium
3.3. Anti-listerial Activity of BLIS Produced by Enterococcus lactis LBM BT2
4. Materials and Methods
4.1. Bacteria Strains and Maintenance
4.2. Sugarcane Molasses
4.3. 16S Ribosomal RNA Identification and Screening of Enterocin-Encoding Genes by PCR
4.4. Inoculum and Culture Medium for E. lactis BT2 Cultivation
4.5. Evaluation of E. lactis LBM BT2 Growth in Different Carbon Sources
4.6. Impact of Molasses Concentration on E. lactis LBM BT2 Growth
4.7. Kinetics of Growth and Bacteriocin Production in Shake Flasks
4.8. Batch Cultivation in 2 L Bioreactor
4.9. Biomass Determination
4.10. Sugars Analyses
4.11. Antimicrobial Activity Quantification
4.12. Spectrum of Action and Antimicrobial Activity of Cell-Free Supernatant
4.13. Partial Purification of BLIS
4.14. Stability of Semi-Purified BLIS to pH, Temperature and Enzymatic Hydrolysis
4.15. Minimum Inhibitory Concentration of Semi-Purified BLIS
4.16. Inhibitory Effect of Semi-Purified BLIS on Biofilm Formation by Listeria monocytogenes
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vidovic, S.; Paturi, G.; Gupta, S.; Fletcher, G.C. Lifestyle of Listeria monocytogenes and food safety: Emerging listericidal technologies in the food industry. Crit. Rev. Food Sci. Nutr. 2022, in press. [Google Scholar] [CrossRef]
- Klopper, K.B.; Bester, E.; Wolfaardt, G.M. Listeria monocytogenes biofilms are planktonic cell factories despite peracetic acid exposure under continuous flow conditions. Antibiotics 2023, 12, 209. [Google Scholar] [CrossRef]
- Pogreba-Brown, K.; Boyd, K.; Schaefer, K.; Austhof, E.; Armstrong, A.; Owusu-Dommey, A.; Villa-Zapata, L.; Arora, M.; McClelland, J.D.; Hoffman, S. Complications associated with foodborne listeriosis: A scoping review. Foodborne Pathog. Dis. 2022, 19, 725–743. [Google Scholar] [CrossRef]
- Kurpas, M.; Wieczorek, K.; Osek, J. Ready-to-eat meat products as a source of Listeria monocytogenes. J. Vet. Res. 2018, 62, 49–55. [Google Scholar] [CrossRef]
- Niederhäusern, S.; Camellini, S.; Sabia, C.; Iseppi, R.; Bondi, M.; Messi, P. Antilisterial Activity of Bacteriocins Produced by Lactic Bacteria Isolated from Dairy Products. Foods 2020, 9, 1757. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, L.; Ji, S.; Li, Q.; Wang, H.; Zhang, Z.; Mao, P.; Sun, H.; Li, L.; Wang, Y.; et al. Dissecting Listeria monocytogenes persistent contamination in a retail market using whole-genome sequencing. Microbiol. Spectr. 2022, 10, e0018522. [Google Scholar] [CrossRef]
- Kayode, A.J.; Okoh, A.I. Antimicrobial-resistant Listeria monocytogenes in ready-to-eat foods: Implications for food safety and risk assessment. Foods 2023, 12, 1346. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Feng, M.; Anwar, T.M.; Chai, W.; Ed-Dra, A.; Kang, X.; Rantsiou, K.; Kehrenberg, C.; Yue, M.; Li, Y. Change in antimicrobial susceptibility of Listeria spp. in response to stress conditions. Front. Sustain. Food Syst. 2023, 7, 1179835. [Google Scholar] [CrossRef]
- Ladjouzi, R.; Dussert, E.; Teiar, R.; Belguesmia, Y.; Drider, D. A review on enterocin DD14, the leaderless two-peptide bacteriocin with multiple biological functions and unusual transport pathway. Antibiotics 2023, 12, 1188. [Google Scholar] [CrossRef]
- Bermúdez-Puga, S.; Dias, M.; Freire de Oliveira, T.; Mendonça, C.M.N.; Yokomizo de Almeida, S.R.; Rozas, E.E.; do Nascimento, C.A.O.; Mendes, M.A.; Oliveira De Souza de Azevedo, P.; Almeida, J.R.; et al. Dual antibacterial mechanism of [K4K15]CZS-1 against Salmonella Typhimurium: A membrane active and intracellular-targeting antimicrobial peptide. Front. Microbiol. 2023, 14, 1320154. [Google Scholar] [CrossRef]
- Hassan, M.; Kjos, M.; Nes, I.F.; Diep, D.B.; Lotfipour, F. Natural antimicrobial peptides from bacteria: Characteristics and potential applications to fight against antibiotic resistance. J. Appl. Microbiol. 2012, 113, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Song, X.; Chen, M.; Tian, S.; Lu, Z.; Sun, J.; Li, X.; Lu, Y.; Yuk, H.-G. Combating biofilms of foodborne pathogens with bacteriocins by lactic acid bacteria in the food industry. Comp. Rev. Food Sci. Food Safety. 2022, 21, 1657–1676. [Google Scholar] [CrossRef] [PubMed]
- Garmasheva, I.L.; Oleschenko, L.T. Screening of bacteriocin-producing dairy Enterococcus strains using low-cost culture media. Front. Microbiol. 2023, 14, 1168835. [Google Scholar] [CrossRef] [PubMed]
- Abbasiliasi, S.; Tan, J.S.; Tengku Ibrahim, T.A.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A.B. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. RSC Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Metsoviti, M.; Paramithiotis, S.; Drosinos, E.H.; Skandamis, P.N.; Galiotou-Panayotou, M.; Papanikolaou, S. Biotechnological valorization of low-cost sugar-based media for bacteriocin production by Leuconostoc mesenteroides E131. New Biotechnol. 2011, 28, 600–609. [Google Scholar] [CrossRef]
- Conab—Safra Brasileira de Cana-de-Açúcar. Available online: https://www.conab.gov.br/info-agro/safras/cana (accessed on 15 January 2024).
- Tang, H.W.; Abbasiliasi, S.; Ng, Z.J.; Lee, Y.-Y.; Tang, T.-K.; Tan, J.S. Insight into the pilot-scale fed-batch fermentation for production of Enterococcus faecium CW3801 using molasses-based medium. Prep. Biochem. Biotechnol. 2022, 52, 691–700. [Google Scholar] [CrossRef]
- Chin, W.L.; Kee, P.E.; Ng, H.S.; Lan, J.C.-W.; Tan, J.S. Selective screening Lactobacillus spp. against Escherichia coli and Salmonella Typhimurium and optimizing a cane molasses-based medium with Lactobacillus acidophilus as a guiding model. J. Taiwan Inst. Chem. Eng. 2023, 105197, in press. [Google Scholar] [CrossRef]
- Almeida-Santos, A.C.; Novais, C.; Peixe, L.; Freitas, A.R. Enterococcus spp. as a producer and target of bacteriocins: A double-edged sword in the antimicrobial resistance crisis context. Antibiotics 2021, 10, 1215. [Google Scholar] [CrossRef]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns-an update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Belloso Daza, M.V.; Cortimiglia, C.; Bassi, D.; Cocconcelli, P.S. Genome-based studies indicate that the Enterococcus faecium Clade B strains belong to Enterococcus lactis species and lack of the hospital infection associated markers. Int. J. Syst. Evol. Microbiol. 2021, 71, 4948. [Google Scholar] [CrossRef]
- Choi, D.G.; Baek, J.H.; Han, D.M.; Khan, S.A.; Jeon, C.O. Comparative pangenome analysis of Enterococcus faecium and Enterococcus lactis provides new insights into the adaptive evolution by horizontal gene acquisitions. BMC Genomics. 2024, 25, 28. [Google Scholar] [CrossRef]
- Belloso Daza, M.V.; Almeida-Santos, A.C.; Novais, C.; Read, A.; Alves, V.; Cocconcelli, P.S.; Freitas, A.R.; Peixe, L. Distinction between Enterococcus faecium and Enterococcus lactis by a gluP PCR-based assay for accurate identification and diagnostics. Microbiol. Spectr. 2022, 10, e0326822. [Google Scholar] [CrossRef]
- Tedim, A.P.; Almeida-Santos, A.C.; Lanza, V.F.; Novais, C.; Coque, T.M.; Freitas, A.R.; Peixe, L. Bacteriocin distribution patterns in Enterococcus faecium and Enterococcus lactis: Bioinformatic analysis using a tailored genomics framework. BioRxiv 2023, in press. [Google Scholar] [CrossRef]
- Kang, J.H.; Lee, M.S. Characterization of a bacteriocin produced by Enterococcus faecium GM-1 isolated from an infant. J. Appl. Microbiol. 2005, 98, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- du Toit, M.; Franz, C.M.; Dicks, L.M.; Holzapfel, W.H. Preliminary characterization of bacteriocins produced by Enterococcus faecium and Enterococcus faecalis isolated from pig faeces. J. Appl. Microbiol. 2000, 88, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Cintas, L.M.; Casaus, P.; Håvarstein, L.S.; Hernández, P.E.; Nes, I.F. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl. Environ. Microbiol. 1997, 63, 4321–4330. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Fang, L.; Zhang, M.; Deng, H.; Cheng, X.; Yao, M.; Huang, L. Isolation and identification of new source of bacteriocin-producing Lactobacillus plantarum C010 and growth kinetics of its batch fermentation. World J. Microbiol. Biotechnol. 2022, 38, 67. [Google Scholar] [CrossRef] [PubMed]
- Wayah, S.B.; Philip, K. Characterization, yield optimization, scale up and biopreservative potential of fermencin SA715, a novel bacteriocin from Lactobacillus fermentum GA715 of goat milk origin. Microb. Cell Fact. 2018, 17, 125. [Google Scholar] [CrossRef]
- Modiri, S.; Kasra Kermanshahi, R.; Reza Soudi, M.; Dad, N.; Ebadi, M.; Shahbani Zahiri, H.; Akbari Noghabi, K. Growth optimization of Lactobacillus acidophilus for production of antimicrobial peptide Acidocin 4356: Scale up from flask to lab-scale fermenter. Iran. J. Biotech. 2021, 19, e2686. [Google Scholar] [CrossRef]
- Jamir, L.; Kumar, V.; Kaur, J.; Kumar, S.; Singh, H. Composition, valorization and therapeutical potential of molasses: A critical review. Environ. Technol. Rev. 2021, 10, 131–142. [Google Scholar] [CrossRef]
- Xu, W.; Liang, L.; Zhu, M. Determination of Sugars in Molasses by HPLC Following Solid-Phase Extraction. Int. J. Food Prop. 2015, 18, 547–557. [Google Scholar] [CrossRef]
- Mulyani, S.; Jenie, B.S.L.; Kusumaningrum, H.D.; Arief, I.I. Characterisation of crude bacteriocin produced by Pediococcus pentosaceus 2A2 in enriched molasses medium. Int. Food Res. J. 2018, 26, 187–192. [Google Scholar]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of culture media, pH and temperature on growth and bacteriocin production of bacteriocinogenic lactic acid bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Herranz, C.; Martínez, J.M.; Rodríguez, J.M.; Hernández, P.E.; Cintas, L.M. Optimization of enterocin P production by batch fermentation of Enterococcus faecium P13 at constant pH. Appl. Microbiol. Biotechnol. 2001, 56, 378–383. [Google Scholar] [CrossRef]
- Ünlü, G.; Nielsen, B.; Ionita, C. Production of antilisterial bacteriocins from lactic acid bacteria in dairy-based media: A comparative study. Probiotics Antimicrob. Proteins 2015, 7, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Jawan, R.; Abbasiliasi, S.; Tan, J.S.; Mustafa, S.; Halim, M.; Ariff, A.B. Influence of culture conditions and medium compositions on the production of bacteriocin-like inhibitory substances by Lactococcus lactis Gh1. Microorganisms 2020, 8, 1454. [Google Scholar] [CrossRef]
- Shokri, D.; Zaghian, S.; Khodabakhsh, F.; Fazeli, H.; Mobasherizadeh, S.; Ataei, B. Antimicrobial activity of a UV-stable bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium strain DSH20 against vancomycin-resistant Enterococcus (VRE) strains. J. Microbiol. Immunol. Infect. 2014, 47, 371–376. [Google Scholar] [CrossRef]
- El-Gendy, A.O.; Brede, D.A.; Essam, T.M.; Amin, M.A.; Ahmed, S.H.; Holo, H.; Nes, I.F.; Shamikh, Y.I. Purification and characterization of bacteriocins-like inhibitory substances from food isolated Enterococcus faecalis OS13 with activity against nosocomial enterococci. Sci. Rep. 2021, 11, 3795. [Google Scholar] [CrossRef]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef]
- Bali, V.; Panesar, P.S.; Bera, M.B.; Kennedy, J.F. Bacteriocins: Recent trends and potential applications. Crit. Rev. Food Sci. Nutr. 2016, 56, 817–834. [Google Scholar] [CrossRef]
- Todorov, S.D.; Popov, I.; Weeks, R.; Chikindas, M.L. Use of bacteriocins and bacteriocinogenic beneficial organisms in food products: Benefits, challenges, concerns. Foods 2022, 11, 3145. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Criado, R.; Martín, M.; Herranz, C.; Cintas, L.M.; Hernández, P.E. Production of enterocin P, an antilisterial pediocin-like bacteriocin from Enterococcus faecium P13, in Pichia pastoris. Antimicrob. Agents Chemother. 2005, 49, 3004–3008. [Google Scholar] [CrossRef]
- Ben Braïek, O.; Ghomrassi, H.; Cremonesi, P.; Morandi, S.; Fleury, Y.; Le Chevalier, P.; Hani, K.; Bel Hadj, O.; Ghrairi, T. Isolation and characterisation of an enterocin P-producing Enterococcus lactis strain from a fresh shrimp (Penaeus vannamei). Antonie Van Leeuwenhoek 2017, 110, 771–786. [Google Scholar] [CrossRef]
- Verma, D.K.; Thakur, M.; Singh, S.; Tripathy, S.; Gupta, A.K.; Baranwal, D.; Patel, A.R.; Shah, N.; Utama, G.L.; Niamah, A.K.; et al. Bacteriocins as antimicrobial and preservative agents in food: Biosynthesis, separation and application. Food Biosci. 2022, 46, 101594. [Google Scholar] [CrossRef]
- Camargo, A.C.; de Paula, O.A.L.; Todorov, S.D.; Nero, L.A. In vitro evaluation of bacteriocins activity against Listeria monocytogenes biofilm formation. Appl. Biochem. Biotechnol. 2016, 178, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Rocha, K.R.; Perini, H.F.; de Souza, C.M.; Schueler, J.; Tosoni, N.F.; Furlaneto, M.C.; Furlaneto-Maia, L. Inhibitory effect of bacteriocins from enterococci on developing and preformed biofilms of Listeria monocytogenes, Listeria ivanovii and Listeria innocua. World J. Microbiol. Biotechnol. 2019, 35, 96. [Google Scholar] [CrossRef] [PubMed]
- Masebe, R.D.; Thantsha, M.S. Anti-biofilm activity of cell free supernatants of selected lactic acid bacteria against Listeria monocytogenes isolated from avocado and cucumber fruits, and from an avocado processing plant. Foods 2022, 11, 2872. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, H.H.; Song, Y.J.; Park, Y.J.; Lee, N.-K.; Paik, H.-D. Anti-biofilm effect of the cell-free supernatant of probiotic Saccharomyces cerevisiae against Listeria monocytogenes. Food Control 2021, 121, 107667. [Google Scholar] [CrossRef]
- Qiao, Z.; Zhang, L.; Wang, X.; Liu, B.; Shan, Y.; Yi, Y.; Zhou, Y.; Lü, X. Antibiofilm effects of Bacteriocin BMP32r on Listeria monocytogenes. Probiotics Antimicrob. Proteins 2022, 14, 1067–1076. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- da Silva Sabo, S.; Lopes, A.M.; de Carvalho Santos-Ebinuma, V.; de Oliveira Rangel-Yagui, C.; de Souza Oliveira, R.P. Bacteriocin partitioning from a clarified fermentation broth of Lactobacillus plantarum ST16Pa in aqueous two-phase systems with sodium sulfate and choline-based salts as additives. Process Biochem. 2018, 66, 212–221. [Google Scholar] [CrossRef]
- M2-A8; Performance Standards for Antimicrobial Disk Susceptibility Tests—Eighth Edition: Approved Standard M2-A8. NCCLS: Villanova, PA, USA, 2003.
- Pato, U.; Yusmarini, Y.; Fitriani, S.; Jonnaidi, N.N.; Wahyuni, M.S.; Feruni, J.A.; Jaswir, I. Antimicrobial activity of lactic acid bacteria strains isolated from Dadih against Escherichia coli. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 709, p. 012019. [Google Scholar] [CrossRef]
- Schägger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Xu, X.; Fan, P.; Qiao, B.; Xie, M.; Huang, T.; Xiong, T. Identification and characterization of a novel pH and heat stable bacteriocin-like substance lactococcin036019 with food preserving potential. Food Control 2023, 148, 109682. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogenic Strain | Diameter of Inhibition Halo (mm) |
|---|---|
| Listeria monocytogenes CET93 | 21.05 ± 0.88 |
| Listeria innocua CLIST 2052 | 11.51 ± 0.68 |
| Carnobacterium piscicola CECT 4020 | 11.75 ± 0.91 |
| Staphylococcus aureus CECT239 | 0 |
| Escherichia coli ATCC 25922 | 0 |
| Salmonella enterica subsp. enterica serovar Typhimurium | 0 |
| Pseudomonas aeruginosa ATCC 27853 | 0 |
| Molasses Concentration (% w/v) | µx (h−1) |
|---|---|
| 2.5 | 1.077 ± 0.07 a |
| 5.0 | 1.070 ± 0.02 a |
| 10.0 | 1.123 ± 0.02 a |
| 15.0 | 1.110 ± 0.01 a |
| 20.0 | 1.101 ± 0.07 a |
| 30.0 | 0.936 ± 0.02 b |
| 40.0 | 0.738 ± 0.05 c |
| Treatments | Antibacterial Activity | |
|---|---|---|
| pH | 2.0 | + |
| 4.0 | + | |
| 8.0 | + | |
| 10.0 | + | |
| Temperature | 60 °C for 30 min | + |
| 80 °C for 30 min | + | |
| 100 °C for 30 min | + | |
| 121 °C for 30 min | − | |
| Enzyme | Proteinase K | − |
| Trypsin | − | |
| Primer Name | Sequence 5′-3′ | Target/Product Size | Annealing Temperature | References |
|---|---|---|---|---|
| entA-For | AAATATTATGGAGTGTAT | Structural enterocin A gene/126 bp | 56 °C | [26] |
| entA-Rev | GCACTTCCCTGGAATTGCTC | |||
| entP1-For | TATGGTAATGGTGTTTATTGTAAT | Structural enterocin P gene/~130 bp | ||
| entP1-Rev | ATGTCCCATACCTGCCAAAC | |||
| ent L501-For | STGGGAGAATCGCAAAATTAG | Structural enterocin L50A or L50B genes/130 bp | ||
| ent L501-Rev | ATTGCCCATCCTTCTCCAAT | |||
| entP2-For | GCTACGCGTTCATATGGTAATGGTG | Structural enterocin P gene/132 bp | 55 °C | [27] |
| entP2-Rev | ATGTCCCATACCTGCCAAACCAGAAGC | |||
| ent L502-For | GATTGGAGGAGTTATATTATGGG | Structural enterocin L50A or L50B genes/556 bp | ||
| ent L502-Rev | CAAATTATAAAGAAATAATTACCTATCATTAAC | |||
| fD1 | AGAGTTTGATCCTGGCTCAG | 16S rRNA gene/ ~1500 bp | 60 °C | [51] |
| rP1 | ACGGTTACCTTGTTACGACTT | |||
| gluP1-For | GCGTGCATGGTTAAGACGAC | Rhomboid protease gene from E. faecalis/427 bp | 61 °C | |
| gluP1-Rev | CTGCTGGATCGCTGGGTTAT | |||
| gluP2-For | TACGGTCACTGGCGGTTTTT | Rhomboid protease gene from E. lactis/324 bp | 58 °C | [23] |
| gluP2-Rev | TGTCTGCTGTTTCGGTAGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, T.F.d.; Kuniyoshi, T.M.; Frota, E.G.; Bermúdez-Puga, S.; Sakaue, L.N.; Cassiano, L.L.; Tachibana, L.; Piccoli, R.A.M.; Converti, A.; Oliveira, R.P.d.S. Anti-Listerial Activity of Bacteriocin-like Inhibitory Substance Produced by Enterococcus lactis LBM BT2 Using Alternative Medium with Sugarcane Molasses. Antibiotics 2024, 13, 210. https://doi.org/10.3390/antibiotics13030210
Oliveira TFd, Kuniyoshi TM, Frota EG, Bermúdez-Puga S, Sakaue LN, Cassiano LL, Tachibana L, Piccoli RAM, Converti A, Oliveira RPdS. Anti-Listerial Activity of Bacteriocin-like Inhibitory Substance Produced by Enterococcus lactis LBM BT2 Using Alternative Medium with Sugarcane Molasses. Antibiotics. 2024; 13(3):210. https://doi.org/10.3390/antibiotics13030210
Chicago/Turabian StyleOliveira, Taciana Freire de, Taís Mayumi Kuniyoshi, Elionio Galvão Frota, Sebastián Bermúdez-Puga, Letícia Naomy Sakaue, Luara Lucena Cassiano, Leonardo Tachibana, Rosane Aparecida Moniz Piccoli, Attilio Converti, and Ricardo Pinheiro de Souza Oliveira. 2024. "Anti-Listerial Activity of Bacteriocin-like Inhibitory Substance Produced by Enterococcus lactis LBM BT2 Using Alternative Medium with Sugarcane Molasses" Antibiotics 13, no. 3: 210. https://doi.org/10.3390/antibiotics13030210
APA StyleOliveira, T. F. d., Kuniyoshi, T. M., Frota, E. G., Bermúdez-Puga, S., Sakaue, L. N., Cassiano, L. L., Tachibana, L., Piccoli, R. A. M., Converti, A., & Oliveira, R. P. d. S. (2024). Anti-Listerial Activity of Bacteriocin-like Inhibitory Substance Produced by Enterococcus lactis LBM BT2 Using Alternative Medium with Sugarcane Molasses. Antibiotics, 13(3), 210. https://doi.org/10.3390/antibiotics13030210