
Prevalence and Genetic Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Pigs in Japan

Abstract
1. Introduction
2. Results
2.1. Prevalence of MRSA in Slaughterhouses
2.2. Molecular Characterization of MRSA Isolates
2.3. Antimicrobial Susceptibility and the AMR Gene
2.4. Zinc Chloride Resistance, PVL Toxin, and Bacteriophage-Mediated Immune Evasion Genes
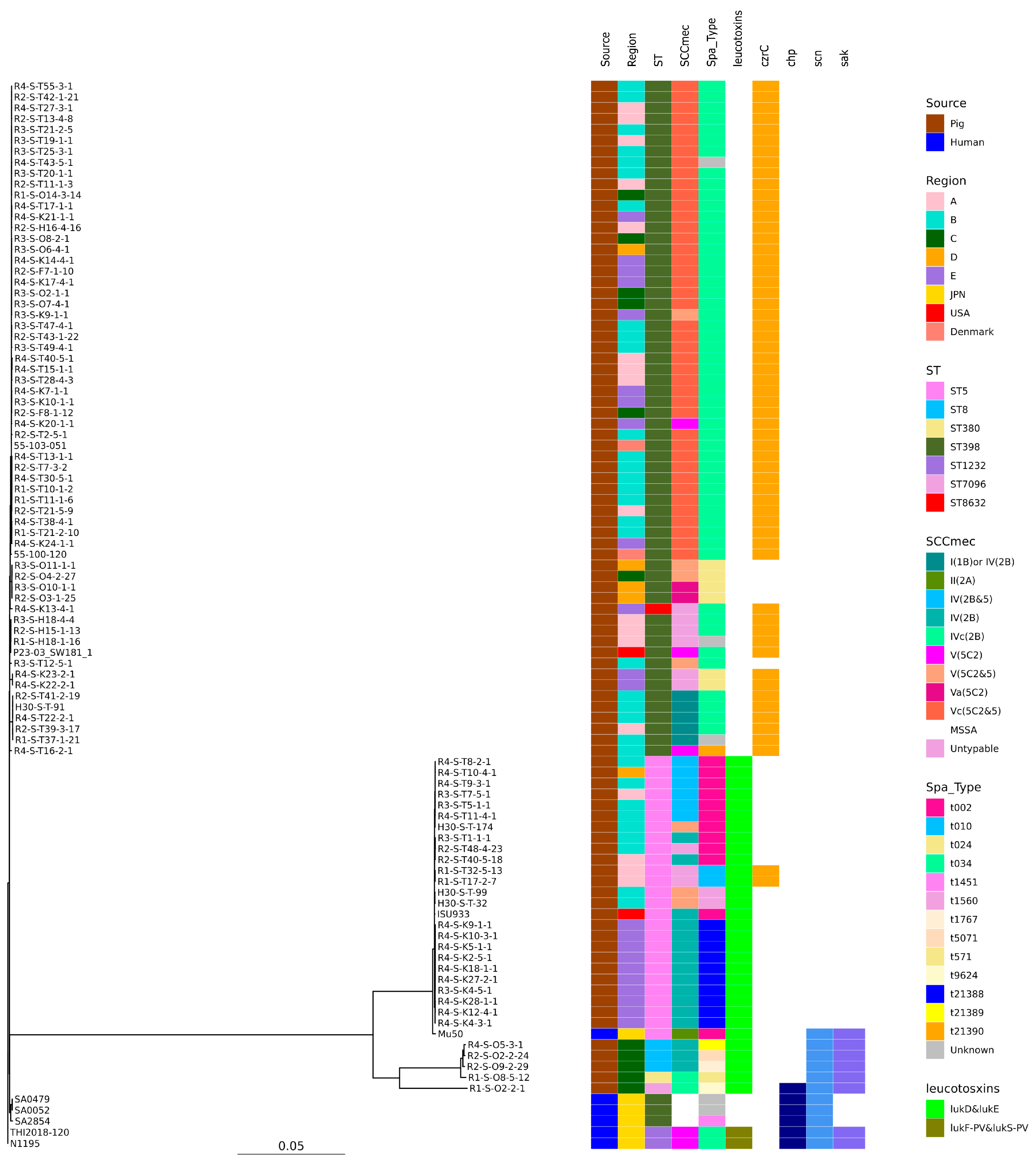
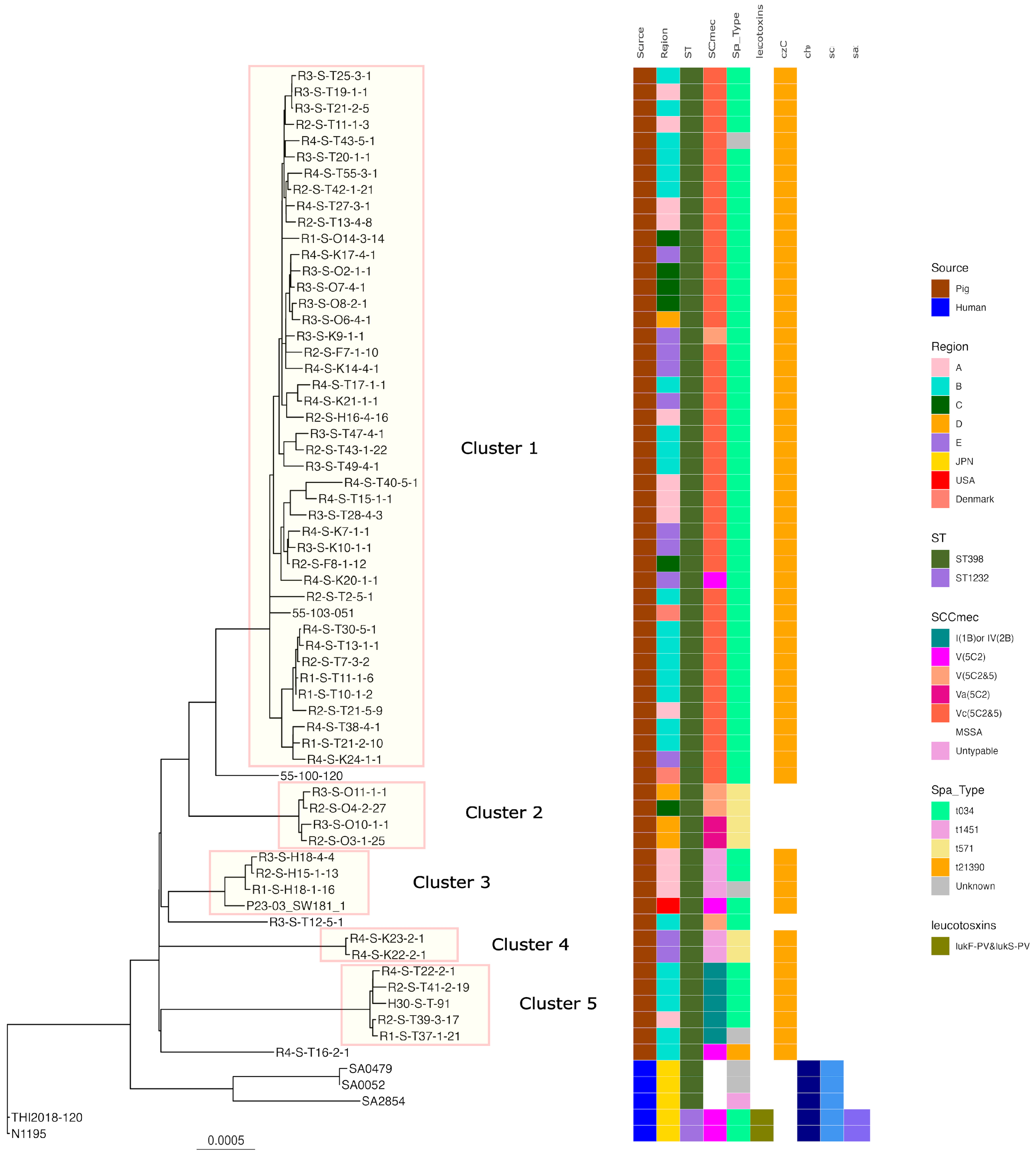
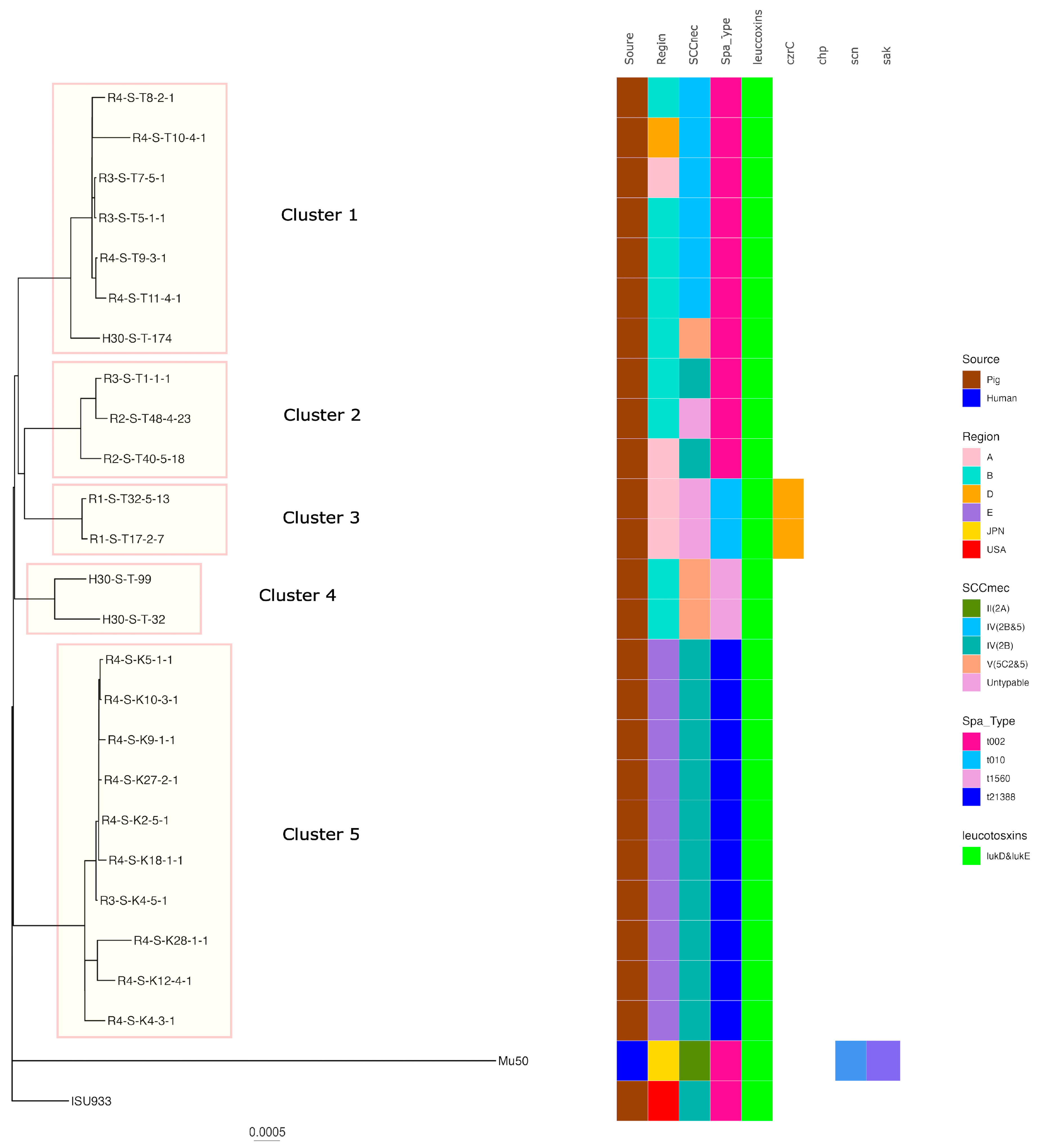
2.5. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Samples from Pigs at the Slaughterhouse
4.2. Staphylococcus aureus Strain Isolation and MRSA Detection
4.3. Antimicrobial Susceptibility Testing
4.4. DNA Sequencing for WGS
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, V.; Araujo, S.; Monteiro, A.; Eira, J.; Pereira, J.E.; Maltez, L.; Igrejas, G.; Lemsaddek, T.S.; Poeta, P. Staphylococcus aureus and MRSA in Livestock: Antimicrobial Resistance and Genetic Lineages. Microorganisms 2023, 11, 124. [Google Scholar] [CrossRef]
- Crespo-Piazuelo, D.; Lawlor, P.G. Livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) prevalence in humans in close contact with animals and measures to reduce on-farm colonisation. Ir. Vet. J. 2021, 74, 21. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef]
- Walesch, S.; Birkelbach, J.; Jezequel, G.; Haeckl, F.P.J.; Hegemann, J.D.; Hesterkamp, T.; Hirsch, A.K.H.; Hammann, P.; Muller, R. Fighting antibiotic resistance-strategies and (pre)clinical developments to find new antibacterials. EMBO Rep. 2023, 24, e56033. [Google Scholar] [CrossRef]
- Khairullah, A.R.; Kurniawan, S.C.; Effendi, M.H.; Sudjarwo, S.A.; Ramandinianto, S.C.; Widodo, A.; Riwu, K.H.P.; Silaen, O.S.M.; Rehman, S. A review of new emerging livestock-associated methicillin-resistant Staphylococcus aureus from pig farms. Vet. World 2023, 16, 46–58. [Google Scholar] [CrossRef]
- Li, J.; Jiang, N.; Ke, Y.; Fessler, A.T.; Wang, Y.; Schwarz, S.; Wu, C. Characterization of pig-associated methicillin-resistant Staphylococcus aureus. Vet. Microbiol. 2017, 201, 183–187. [Google Scholar] [CrossRef]
- Moon, D.C.; Jeong, S.K.; Hyun, B.H.; Lim, S.K. Prevalence and Characteristics of Methicillin-Resistant Staphylococcus aureus Isolates in Pigs and Pig Farmers in Korea. Foodborne Pathog. Dis. 2019, 16, 256–261. [Google Scholar] [CrossRef]
- Sahibzada, S.; Abraham, S.; Coombs, G.W.; Pang, S.; Hernández-Jover, M.; Jordan, D.; Heller, J. Transmission of highly virulent community-associated MRSA ST93 and livestock-associated MRSA ST398 between humans and pigs in Australia. Sci. Rep. 2017, 7, 5273. [Google Scholar] [CrossRef]
- Tanomsridachchai, W.; Changkaew, K.; Changkwanyeun, R.; Prapasawat, W.; Intarapuk, A.; Fukushima, Y.; Yamasamit, N.; Flav Kapalamula, T.; Nakajima, C.; Suthienkul, O.; et al. Antimicrobial Resistance and Molecular Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Slaughtered Pigs and Pork in the Central Region of Thailand. Antibiotics 2021, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- de Neeling, A.J.; van den Broek, M.J.; Spalburg, E.C.; van Santen-Verheuvel, M.G.; Dam-Deisz, W.D.; Boshuizen, H.C.; van de Giessen, A.W.; van Duijkeren, E.; Huijsdens, X.W. High prevalence of methicillin resistant Staphylococcus aureus in pigs. Vet. Microbiol. 2007, 122, 366–372. [Google Scholar] [CrossRef]
- Khanna, T.; Friendship, R.; Dewey, C.; Weese, J.S. Methicillin resistant Staphylococcus aureus colonization in pigs and pig farmers. Vet. Microbiol. 2008, 128, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Lewis, H.C.; Mølbak, K.; Reese, C.; Aarestrup, F.M.; Selchau, M.; Sørum, M.; Skov, R.L. Pigs as source of methicillin-resistant Staphylococcus aureus CC398 infections in humans, Denmark. Emerg. Infect. Dis. 2008, 14, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- DANMAP 2019—Use of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Food Animals, Food and Humans in Denmark. 2019. Available online: https://backend.orbit.dtu.dk/ws/files/235092204/DANMAP_2019.pdf (accessed on 2 October 2023).
- MARAN 2021 Monitoring of Antimicrobial Resistance and Antibiotic Usage in Animals in the Netherlands. 2021. Available online: https://rivm.openrepository.com/handle/10029/625006?show=full (accessed on 2 October 2023).
- Baba, K.; Ishihara, K.; Ozawa, M.; Tamura, Y.; Asai, T. Isolation of meticillin-resistant Staphylococcus aureus (MRSA) from swine in Japan. Int. J. Antimicrob. Agents 2010, 36, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Usui, M.; Motoya, T.; Sugiyama, T.; Tamura, Y. Characterisation of meticillin-resistant Staphylococcus aureus ST97 and ST5 isolated from pigs in Japan. J. Glob. Antimicrob. Resist. 2015, 3, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Yamanaka, M.; Nara, K.; Tanaka, S.; Uema, M.; Asai, T.; Tamura, Y. Isolation of ST398 methicillin-resistant Staphylococcus aureus from pigs at abattoirs in Tohoku region, Japan. J. Vet. Med. Sci. 2020, 82, 1400–1403. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Furuya, Y.; Akama, R.; Harada, S.; Matsuda, M.; Abo, H.; Shirakawa, T.; Kawanishi, M.; Yoshida, E.; Furuno, M.; et al. Molecular epidemiology of methicillin-resistant Staphylococcus aureus isolated from pigs in Japan. Vet. Microbiol. 2022, 273, 109523. [Google Scholar] [CrossRef]
- Van Cleef, B.A.; Broens, E.M.; Voss, A.; Huijsdens, X.W.; Zuchner, L.; Van Benthem, B.H.; Kluytmans, J.A.; Mulders, M.N.; Van De Giessen, A.W. High prevalence of nasal MRSA carriage in slaughterhouse workers in contact with live pigs in The Netherlands. Epidemiol. Infect. 2010, 138, 756–763. [Google Scholar] [CrossRef]
- Smith, T.C.; Gebreyes, W.A.; Abley, M.J.; Harper, A.L.; Forshey, B.M.; Male, M.J.; Martin, H.W.; Molla, B.Z.; Sreevatsan, S.; Thakur, S.; et al. Methicillin-resistant Staphylococcus aureus in pigs and farm workers on conventional and antibiotic-free swine farms in the USA. PLoS ONE 2013, 8, e63704. [Google Scholar] [CrossRef]
- Kinross, P.; Petersen, A.; Skov, R.; Van Hauwermeiren, E.; Pantosti, A.; Laurent, F.; Voss, A.; Kluytmans, J.; Struelens, M.J.; Heuer, O.; et al. Livestock-associated meticillin-resistant Staphylococcus aureus (MRSA) among human MRSA isolates, European Union/European Economic Area countries, 2013. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Kadlec, K.; Fessler, A.T.; Hauschild, T.; Schwarz, S. Novel and uncommon antimicrobial resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Tenhagen, B.A.; Fetsch, A.; Sachsenröder, J.; Käsbohrer, A.; Schroeter, A.; Hammerl, J.A.; Hertwig, S.; Helmuth, R.; Bräunig, J.; et al. Virulence and resistance determinants of German Staphylococcus aureus ST398 isolates from nonhuman sources. Appl. Environ. Microbiol. 2011, 77, 3052–3060. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.C.; Zishiri, O.T.; El Zowalaty, M.E. Methicillin-resistant Staphylococcus aureus: Livestock-associated, antimicrobial, and heavy metal resistance. Infect. Drug Resist. 2018, 11, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Nakaminami, H.; Takadama, S.; Ito, A.; Hasegawa, M.; Jono, C.; Noguchi, M.; Shoshi, M.; Wajima, T.; Fujii, T.; Maruyama, H.; et al. Characterization of SCCmec type IV methicillin-resistant Staphylococcus aureus clones increased in Japanese hospitals. J. Med. Microbiol. 2018, 67, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Sanui, M.; Saga, T.; Harada, S.; Ishii, Y.; Tateda, K.; Lefor, A.K. A fatal infection caused by sequence type 398 methicillin-resistant Staphylococcus aureus carrying the Panton-Valentine leukocidin gene: A case report in Japan. J. Infect. Chemother. 2015, 21, 541–543. [Google Scholar] [CrossRef]
- Weese, J.S. Methicillin-resistant Staphylococcus aureus in animals. ILAR J. 2010, 51, 233–244. [Google Scholar] [CrossRef]
- Ho, P.L.; Chow, K.H.; Lai, E.L.; Law, P.Y.; Chan, P.Y.; Ho, A.Y.; Ng, T.K.; Yam, W.C. Clonality and antimicrobial susceptibility of Staphylococcus aureus and methicillin-resistant S. aureus isolates from food animals and other animals. J. Clin. Microbiol. 2012, 50, 3735–3737. [Google Scholar] [CrossRef]
- Yu, F.; Cienfuegos-Gallet, A.V.; Cunningham, M.H.; Jin, Y.; Wang, B.; Kreiswirth, B.N.; Chen, L. Molecular Evolution and Adaptation of Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) Sequence Type 9. mSystems 2021, 6, e0049221. [Google Scholar] [CrossRef]
- Wagenaar, J.A.; Yue, H.; Pritchard, J.; Broekhuizen-Stins, M.; Huijsdens, X.; Mevius, D.J.; Bosch, T.; Van Duijkeren, E. Unexpected sequence types in livestock associated methicillin-resistant Staphylococcus aureus (MRSA): MRSA ST9 and a single locus variant of ST9 in pig farming in China. Vet. Microbiol. 2009, 139, 405–409. [Google Scholar] [CrossRef]
- Asai, T.; Hiki, M.; Baba, K.; Usui, M.; Ishihara, K.; Tamura, Y. Presence of Staphylococcus aureus ST398 and ST9 in Swine in Japan. Jpn. J. Infect. Dis. 2012, 65, 551–552. [Google Scholar] [CrossRef]
- Furuno, M.; Uchiyama, M.; Nakahara, Y.; Uenoyama, K.; Fukuhara, H.; Morino, S.; Kijima, M. A Japanese trial to monitor methicillin-resistant Staphylococcus aureus (MRSA) in imported swine during the quarantine period. J. Glob. Antimicrob. Resist. 2018, 14, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Dorado-Garcia, A.; Dohmen, W.; Bos, M.E.; Verstappen, K.M.; Houben, M.; Wagenaar, J.A.; Heederik, D.J. Dose-response relationship between antimicrobial drugs and livestock-associated MRSA in pig farming. Emerg. Infect. Dis. 2015, 21, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Dadi, T.H.; Pieper, L.; Vahjen, W.; Franke, A.; Reinert, K.; Zentek, J. Concentration and chemical form of dietary zinc shape the porcine colon microbiome, its functional capacity and antibiotic resistance gene repertoire. ISME J. 2020, 14, 2783–2793. [Google Scholar] [CrossRef] [PubMed]
- Slifierz, M.J.; Friendship, R.; Weese, J.S. Zinc oxide therapy increases prevalence and persistence of methicillin-resistant Staphylococcus aureus in pigs: A randomized controlled trial. Zoonoses Public Health 2015, 62, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Kamio, Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: Structures, pore-forming mechanism, and organization of the genes. Biosci. Biotechnol. Biochem. 2004, 68, 981–1003. [Google Scholar] [CrossRef] [PubMed]
- Laumay, F.; Benchetrit, H.; Corvaglia, A.R.; van der Mee-Marquet, N.; Francois, P. The Staphylococcus aureus CC398 Lineage: An Evolution Driven by the Acquisition of Prophages and Other Mobile Genetic Elements. Genes 2021, 12, 1752. [Google Scholar] [CrossRef]
- Hau, S.J.; Sun, J.; Davies, P.R.; Frana, T.S.; Nicholson, T.L. Comparative Prevalence of Immune Evasion Complex Genes Associated with beta-Hemolysin Converting Bacteriophages in MRSA ST5 Isolates from Swine, Swine Facilities, Humans with Swine Contact, and Humans with No Swine Contact. PLoS ONE 2015, 10, e0142832. [Google Scholar] [CrossRef]
- Hau, S.J.; Haan, J.S.; Davies, P.R.; Frana, T.; Nicholson, T.L. Antimicrobial Resistance Distribution Differs Among Methicillin Resistant Staphylococcus aureus Sequence Type (ST) 5 Isolates From Health Care and Agricultural Sources. Front. Microbiol. 2018, 9, 2102. [Google Scholar] [CrossRef]
- Tenover, F.C.; McDougal, L.K.; Goering, R.V.; Killgore, G.; Projan, S.J.; Patel, J.B.; Dunman, P.M. Characterization of a strain of community-associated methicillin-resistant Staphylococcus aureus widely disseminated in the United States. J. Clin. Microbiol. 2006, 44, 108–118. [Google Scholar] [CrossRef]
- Iwao, Y.; Ishii, R.; Tomita, Y.; Shibuya, Y.; Takano, T.; Hung, W.C.; Higuchi, W.; Isobe, H.; Nishiyama, A.; Yano, M.; et al. The emerging ST8 methicillin-resistant Staphylococcus aureus clone in the community in Japan: Associated infections, genetic diversity, and comparative genomics. J. Infect. Chemother. 2012, 18, 228–240. [Google Scholar] [CrossRef]
- Hagiya, H.; Hisatsune, J.; Kojima, T.; Shiota, S.; Naito, H.; Hagioka, S.; Morimoto, N.; Otsuka, F.; Sugai, M. Comprehensive analysis of systemically disseminated ST8/non-USA300 type community-acquired methicillin-resistant Staphylococcus aureus infection. Intern. Med. 2014, 53, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, J.; Hagiya, H.; Shiota, S.; Sugai, M. Complete Genome Sequence of Systemically Disseminated Sequence Type 8 Staphylococcal Cassette Chromosome mec Type IVl Community-Acquired Methicillin-Resistant Staphylococcus aureus. Genome Announc. 2017, 5. [Google Scholar] [CrossRef]
- Authority, E.F.S. Analysis of the baseline survey on the prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in holdings with breeding pigs, in the EU, 2008—Part A: MRSA prevalence estimates. EFSA J. 2009, 7, 1376. [Google Scholar] [CrossRef]
- Pérez-Roth, E.; Claverie-Martín, F.; Villar, J.; Méndez-Alvarez, S. Multiplex PCR for simultaneous identification of Staphylococcus aureus and detection of methicillin and mupirocin resistance. J. Clin. Microbiol. 2001, 39, 4037–4041. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, T.M. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; VET01; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Furuya, Y.; Matsuda, M.; Harada, S.; Kumakawa, M.; Shirakawa, T.; Uchiyama, M.; Akama, R.; Ozawa, M.; Kawanishi, M.; Shimazaki, Y.; et al. Nationwide Monitoring of Antimicrobial-Resistant Escherichia coli and Enterococcus spp. Isolated From Diseased and Healthy Dogs and Cats in Japan. Front. Vet. Sci. 2022, 9, 916461. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| 2018 * | 2019 * | 2020 * | 2021 * | 2022 * | |
|---|---|---|---|---|---|
| Number of slaughterhouses | 2 | 3 | 4 | 5 | 6 |
| Number of pigs sampled | 240 | 375 | 465 | 515 | 575 |
| Number of positive samples | 7 | 24 | 29 | 39 | 57 |
| Positive rate of samples | 2.9% | 6.4% | 6.2% | 7.6% ** | 9.9% ** |
| Number of farms | 48 | 75 | 93 | 103 | 115 |
| Number of positive farms | 4 | 10 | 19 | 21 | 34 |
| Positive rate of farms | 8.3% | 13.3% | 20.4% | 20.4% | 29.6% ** |
| 2018 * | 2019 * | 2020 * | 2021 * | 2022 * | |
|---|---|---|---|---|---|
| Number of slaughterhouses | 2 | 2 | 2 | 2 | 2 |
| Number of pigs sampled | 240 | 300 | 340 | 340 | 320 |
| Number of positive samples | 7 | 20 | 20 | 24 | 23 |
| Positive rate of samples | 2.9% | 6.7% ** | 5.9% | 7.1% ** | 7.2%** |
| Number of farms | 48 | 60 | 68 | 68 | 64 |
| Number of positive farms | 4 | 7 | 13 | 12 | 15 |
| Positive rate of farms | 8.3% | 11.7% | 19.1% | 17.6% | 23.4% ** |
| ST | Spa Typing | SCCmec | ||||
|---|---|---|---|---|---|---|
| Number | Rate (%) | Number | Rate (%) | |||
| ST398 | t034 | 48 | 54.5 | Vc (5C2&5) | 40 | 45.5 |
| t571 | 6 | 6.8 | V (5C2&5) | 4 | 4.5 | |
| t21390 | 1 | 1.1 | V (5C2) | 2 | 2.3 | |
| UD | 3 | 3.4 | Va (5C2) | 2 | 2.3 | |
| t21388 | I (1B) or IV (2B) | 5 | ||||
| Untypable | 5 | |||||
| 58 | 65.9 | 58 | 65.9 | |||
| ST5 | t002 | 10 | 11.4 | IV (2B) | 12 | 13.6 |
| t21388 | 10 | 11.4 | IV (2B&5) | 6 | 6.8 | |
| t010 | 2 | 2.3 | V (5C2&5) | 3 | 3.4 | |
| t1560 | 2 | 2.3 | Untypable | 3 | 3.4 | |
| 24 | 27.3 | 24 | 27.3 | |||
| ST8 | t21389 | 1 | 1.1 | IV(2B) | 3 | 3.4 |
| t1767 | 1 | 1.1 | 0.0 | |||
| t5071 | 1 | 1.1 | 0.0 | |||
| 3 | 3.4 | 3 | 3.4 | |||
| ST380 | t024 | 1 | 1.1 | IVc (2B) | 1 | 1.1 |
| 1 | 1.1 | 1 | 1.1 | |||
| ST7096 | t9624 | 1 | 1.1 | IVc (2B) | 1 | 1.1 |
| 1 | 1.1 | 1 | 1.1 | |||
| ST8632 | t034 | 1 | 1.1 | Untypable | 1 | 1.1 |
| 1 | 1.1 | 1 | 1.1 | |||
| total | 88 | 88 | ||||
| Resistant Rate (%) | |||||
|---|---|---|---|---|---|
| Antimicrobial Agents | BP | Total (n = 88) | ST398 (n = 58) | ST5 (n = 24) | p-Values Determined by Fisher’s Exact Test (ST398 vs. ST5) |
| MPIPC | 4 | 100 | 100 | 100 | - |
| CFX | 8 | 100 | 100 | 100 | - |
| TC | 16 | 85.2 | 98.3 | 62.5 | p < 0.05 |
| AZM | 8 | 46.6 | 37.9 | 66.7 | p < 0.05 |
| EM | 8 | 46.6 | 37.9 | 66.7 | p < 0.05 |
| CP | 32 | 43.2 | 41.4 | 54.2 | - |
| SM | 64 | 31.8 | 36.2 | 20.8 | - |
| GM | 16 | 2.3 | 0 | 0 | - |
| CPFX | 4 | 18.2 | 8.6 | 33.3 | p < 0.05 |
| Antimicrobial Resistance Genes/ | Total (n = 88) | ST398 (n = 58) | ST5 (n = 24) | ||||
|---|---|---|---|---|---|---|---|
| Antimicrobial Class | Mutation | Number | Rate (%) | Number | Rate (%) in ST398 | Number | Rate (%) in ST5 |
| beta-lactam | mecA | 88 | 100 | 58 | 100 | 24 | 100 |
| tetracycline | tet(K) | 7 | 8.0 | 6 | 25.0 | ||
| tet(L) | 8 | 9.1 | 8 | 33.3 | |||
| tet(M) | 11 | 12.5 | 10 | 17.2 | |||
| tet(K) + tet(M) | 47 | 53.4 | 46 | 79.3 | |||
| tet(L) + tet(M) | 2 | 2.3 | 2 | 3.4 | |||
| 75 | 85.23 | 58 | 100 | 14 | 58.3 | ||
| macrolide | erm(A) | 4 | 4.5 | 3 | 5.2 | ||
| erm(C) | 40 | 45.5 | 22 | 37.9 | 17 | 70.8 | |
| 44 | 50.0 | 25 | 43.1 | 17 | 70.8 | ||
| lincosamide/ | vga(A) | 24 | 27.3 | 2 | 3.4 | 22 | 91.7 |
| streptogramin | vga(E) | 2 | 2.3 | 2 | 3.4 | ||
| lsa(E) + lnu(B) | 36 | 40.9 | 36 | 62.1 | |||
| lsa(E) + lnu(B) + vga(A) | 1 | 1.1 | 1 | 1.7 | |||
| lsa(E) + lnu(B) + vga(E) | 5 | 5.7 | 5 | 8.6 | |||
| lsa(E) + vga(A) | 2 | 2.3 | 0.0 | 2 | 8.3 | ||
| 70.0 | 79.5 | 46.0 | 79.3 | 24 | 100.0 | ||
| phenicol | catA | 3 | 3.4 | 3 | 5.2 | ||
| fexA | 32 | 36.4 | 20 | 34.5 | 12 | 50.0 | |
| 35 | 39.8 | 23 | 39.7 | 12 | 50.0 | ||
| aminoglycoside | aadD1 | 6 | 6.8 | 6 | 25.0 | ||
| aadD1 + ant(6)-Ia + spw | 1 | 1.1 | 1 | 4.2 | |||
| aadD1 + ant(6)-Ia + spw + str | 1 | 1.1 | 1 | 4.2 | |||
| ant(9)-Ia | 31 | 35.2 | 30 | 51.7 | |||
| ant(9)-Ia + str | 15 | 17.0 | 15 | 25.9 | |||
| aac(6′)-Ie/aph(2″)-Ia | 2 | 2.3 | |||||
| str | 4 | 4.5 | 3 | 5.2 | 1 | 4.2 | |
| 60 | 68.2 | 48.0 | 82.8 | 9 | 37.5 | ||
| trimethoprim | dfrG | 49 | 55.7 | 46 | 79.3 | 3 | 12.5 |
| 49 | 55.7 | 46 | 79.3 | 3 | 12.5 | ||
| quinolone | gyrA_S84A + parC_S80F | 7 | 8.0 | 7 | 29.2 | ||
| gyrA_S84L + parC_S80F | 5 | 5.7 | 2 | 8.3 | |||
| gyrA_S84L + parC_S80Y | 4 | 4.5 | 4 | 6.9 | |||
| parC_S80F | 7 | 8.0 | 3 | 5.2 | 4 | 16.7 | |
| 23 | 26.1 | 7 | 12.1 | 13 | 54.2 | ||
| Total | ST398 | ST5 | |||||
|---|---|---|---|---|---|---|---|
| Number | Rate (%) | Number | Rate (%) | Number | Rate (%) | ||
| czrC | + | 56 | 63.6 | 53 | 91.4 | 2 | 8.3 |
| − | 32 | 36.4 | 5 | 8.6 | 22 | 91.7 | |
| LukD + LukE | + | 29 | 33.0 | 0 | 0 | 24 | 100 |
| − | 59 | 67.0 | 58 | 100 | 0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawanishi, M.; Matsuda, M.; Abo, H.; Ozawa, M.; Hosoi, Y.; Hiraoka, Y.; Harada, S.; Kumakawa, M.; Sekiguchi, H. Prevalence and Genetic Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Pigs in Japan. Antibiotics 2024, 13, 155. https://doi.org/10.3390/antibiotics13020155
Kawanishi M, Matsuda M, Abo H, Ozawa M, Hosoi Y, Hiraoka Y, Harada S, Kumakawa M, Sekiguchi H. Prevalence and Genetic Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Pigs in Japan. Antibiotics. 2024; 13(2):155. https://doi.org/10.3390/antibiotics13020155
Chicago/Turabian StyleKawanishi, Michiko, Mari Matsuda, Hitoshi Abo, Manao Ozawa, Yuta Hosoi, Yukari Hiraoka, Saki Harada, Mio Kumakawa, and Hideto Sekiguchi. 2024. "Prevalence and Genetic Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Pigs in Japan" Antibiotics 13, no. 2: 155. https://doi.org/10.3390/antibiotics13020155
APA StyleKawanishi, M., Matsuda, M., Abo, H., Ozawa, M., Hosoi, Y., Hiraoka, Y., Harada, S., Kumakawa, M., & Sekiguchi, H. (2024). Prevalence and Genetic Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Pigs in Japan. Antibiotics, 13(2), 155. https://doi.org/10.3390/antibiotics13020155