
Antimicrobial Resistance in Enterococcus spp. Isolates from Red Foxes (Vulpes vulpes) in Latvia

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Antimicrobial Resistance in Enterococcus Isolates
2.2. Distribution of Antimicrobial Resistance Genes (ARGs) in Enterococcus Isolates
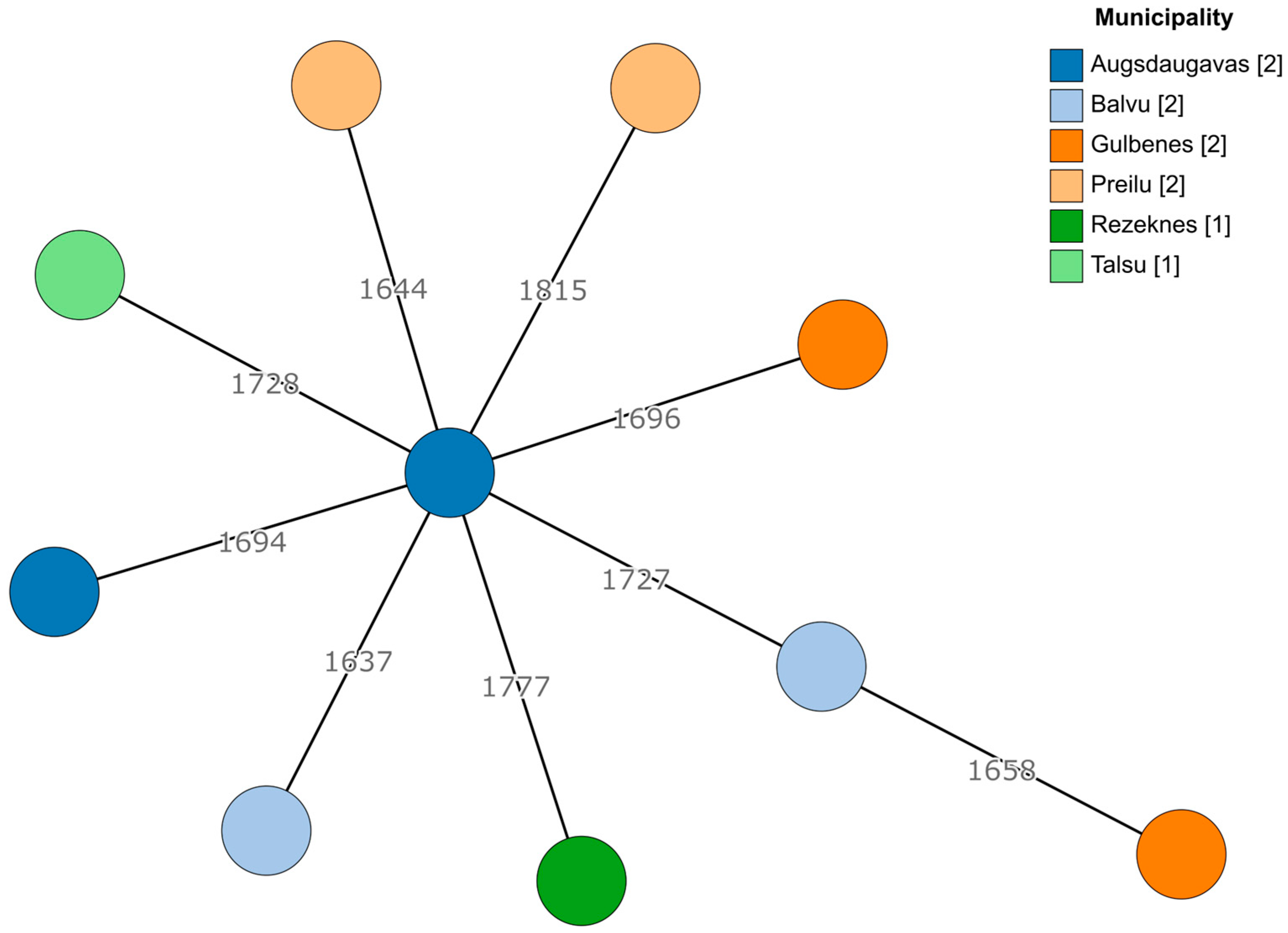
2.3. Genetic Diversity in Enterococcus Isolates
3. Discussion
4. Materials and Methods
4.1. Samples and Sampling
4.2. Microbiological Testing of Samples
4.3. Detection of Antimicrobial Resistance in Enterococcus spp. Isolates
4.4. DNA Extraction and Whole-Genome Sequencing (WGS)
4.5. Genome Assembly, Detection of Antimicrobial Resistance Genes (ARGs) and Core Genome MLST (cgMLST)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aarestrup, F.M.; Butaye, P.; Witte, W. Non-human reservoirs of enterococci. In The Enterococci: Pathogenesis, Molecular Biology, and Antibiotic Resistance; Gilmore, M.S., Clewell, D.B., Courvalin, P., Dunny, G.M., Murray, B.E., Rice, L.B., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 55–100. [Google Scholar]
- Gilmore, M.S.; Lebreton, F.; van Schaik, W. Genomic transition of enterococci from gut commensals to leading causes of multidrug-resistant hospital infection in the antibiotic era. Curr. Opin. Microbiol. 2013, 16, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Whitman, R.L.; Shively, D.A.; Pawlik, H.; Nevers, M.B.; Byappanahalli, M.N. Occurrence of Escherichia coli and enterococci in Cladophora (Chlorophyta) in nearshore water and beach sand of Lake Michigan. Appl. Environ. Microbiol. 2003, 69, 4717–4719. [Google Scholar] [CrossRef] [PubMed]
- Staley, C.; Dunny, G.M.; Sadowsky, M.J. Environmental and animal-associated Enterococci. Adv. Appl. Microbiol. 2014, 87, 147–186. [Google Scholar] [CrossRef]
- Novais, C.; Campos, J.; Freitas, A.R.; Barros, M.; Silveira, E.; Coque, T.M.; Antunes, P.; Peixe, L. Water supply and feed as sources of antimicrobial-resistant Enterococcus spp. in aquacultures of rainbow trout (Oncorhyncus mykiss), Portugal. Sci. Total Environ. 2018, 625, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zarzecka, U.; Zadernowska, A. Enterococci isolated from plant-derived food—Analysis of antibiotic resistance and the occurrence of resistance genes. LWT 2021, 139, 110549. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Kowalczewski, P.Ł.; Babošová, M.; Porhajašová, J.I.; Hikal, W.M.; Fedoriak, M. Bacteriota and Antibiotic Resistance in Spiders. Insects 2022, 13, 680. [Google Scholar] [CrossRef] [PubMed]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Recommendations incorporating the first and second addenda. In Guidelines for Drinking-Water Quality, 3rd ed.; WHO: Geneve, Switzerland, 2008; Volume 1, p. 688. Available online: https://www.who.int/publications/i/item/9789241547611 (accessed on 10 December 2023).
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef]
- Brinkwirth, S.; Ayobami, O.; Eckmanns, T.; Markwart, R. Hospital-acquired infections caused by enterococci: A systematic review and meta-analysis, WHO European Region, 1 January 2010 to 4 February 2020. Euro Surveill. 2021, 26, 2001628. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef]
- Cimen, C.; Berends, M.S.; Bathoorn, E.; Lokate, M.; Voss, A.; Friedrich, A.W.; Glasner, C.; Hamprecht, A. Vancomycin-resistant enterococci (VRE) in hospital settings across European borders: A scoping review comparing the epidemiology in the Netherlands and Germany. Antimicrob. Resist. Infect. Control 2023, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial Resistance in the EU/EEA (EARS-Net)-Annual Epidemiological Report 2019; ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- Markwart, R.; Willrich, N.; Haller, S.; Noll, I.; Koppe, U.; Werner, G.; Eckmanns, T. The rise in vancomycin-resistant Enterococcus faecium in Germany: Data from the German Antimicrobial Resistance Surveillance (ARS). Antimicrob. Resist. Infect. Control 2019, 8, 147. [Google Scholar] [CrossRef]
- Paschoalini, B.R.; Nuñez, K.V.M.; Maffei, J.T.; Langoni, H.; Guimarães, F.F.; Gebara, C.; Freitas, N.E.; dos Santos, M.V.; Fidelis, C.E.; Kappes, R.; et al. The emergence of antimicrobial resistance and virulence characteristics in Enterococcus species isolated from bovine milk. Antibiotics 2023, 12, 1243. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, S.P.; Stokes, H.W.; Roy Chowdhury, P. Mobile elements, zoonotic pathogens and commensal bacteria: Conduits for the delivery of resistance genes into humans, production animals and soil microbiota. Front. Microbiol. 2023, 4, 86. [Google Scholar] [CrossRef] [PubMed]
- Lawpidet, P.; Tengjaroenkul, B.; Saksangawong, C.; Sukon, P. Global prevalence of vancomycin-resistant enterococci in food of animal origin: A meta-analysis. Foodborne Pathog. Dis. 2021, 18, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Dungan, R.S.; Bjorneber, D.L. Antimicrobial resistance in Escherichia coli and enterococcal isolates from irrigation return flows in a high-desert watershed. Front. Microbiol. 2021, 12, 660697. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar] [CrossRef]
- Anjum, M.F.; Schmitt, H.B.; Börjesson, S.; Berendonk, T.U.; on behalf of the WAWES network. The potential of using E. coli as an indicator for the surveillance of antimicrobial resistance (AMR) in the environment. Curr. Opin. Microbiol. 2021, 64, 152–158. [Google Scholar] [CrossRef]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef]
- Biggel, M.; Nüesch-Inderbinen, M.; Raschle, S.; Stevens, M.J.A.; Stephan, R. Spread of vancomycin-resistant Enterococcus faecium ST133 in the aquatic environment in Switzerland. J. Glob. Antimicrob. Resist. 2021, 27, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Decision (EU) 2020/1729 of 17 November 2020 on the Monitoring and Reporting of Antimicrobial Resistance in Zoonotic and Commensal Bacteria and Repealing Implementing Decision 2013/652/EU. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32020D1729 (accessed on 1 December 2023).
- Malik, F.; Nawaz, M.; Anjum, A.A.; Firyal, S.; Shahid, M.A.; Irfan, S.; Ahmed, F.; Bhatti, A.A. Molecular characterization of antibiotic resistance in poultry gut origin Enterococci and horizontal gene transfer of antibiotic resistance to Staphylococcus aureus. Pak. Vet. J. 2022, 42, 383–389. [Google Scholar] [CrossRef]
- Grassotti, T.T.; de Angelis Zvoboda, D.; da Fontoura Xavier Costa, L.; de Araújo, A.J.G.; Pereira, R.I.; Soares, R.O.; Wagner, P.G.C.; Frazzon, J.; Frazzon, A.P.G. Antimicrobial resistance profiles in Enterococcus spp. isolates from fecal samples of wild and captive black capuchin monkeys (Sapajus nigritus) in South Brazil. Front. Microbiol. 2018, 9, 2366. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as sentinels of antimicrobial resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [CrossRef] [PubMed]
- Nowakiewicz, A.; Zięba, P.; Gnat, S.; Trościeńczyk, A.; Osińska, M.; Łagowski, D.; Kosior-Korzecka, U.; Puzio, I. A significant number of multi-drug resistant Enterococcus faecalis in wildlife animals; Long-term consequences and new or known reservoirs of resistance. Sci. Total Environ. 2020, 705, 135830. [Google Scholar] [CrossRef] [PubMed]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthier-Clerc, M.; et al. Antimicrobial Resistance in Wildlife. J. Appl. Ecol. 2016, 53, 519–529. [Google Scholar] [CrossRef]
- Díaz-Ruiz, F.; Delibes-Mateos, M.; García-Moreno, J.L.; López-Martín, J.M.; Ferreira, C.; Ferrers, P. Biogeographical patterns in the diet of an opportunistic predator: The red fox Vulpes vulpes in the Iberian Peninsula. Mammal Rev. 2013, 43, 59–70. [Google Scholar] [CrossRef]
- Lindsø, L.K.; Dupont, P.; Rød-Eriksen, L.; Andersskog, I.P.Ø.; Ulvund, K.R.; Flagstad, Ø.; Bischof, R.; Eide, N.E. Estimating red fox density using non-invasive genetic sampling and spatial capture–recapture modelling. Oecologia 2022, 198, 139–151. [Google Scholar] [CrossRef]
- Mo, S.S.; Urdahl, A.M.; Madslien, K.; Sunde, M.; Nesse, L.L.; Slettemeås, J.S.; Norströn, M. What does the fox say? Monitoring antimicrobial resistance in the environment using wild red foxes as an indicator. PLoS ONE 2018, 13, e0198019. [Google Scholar] [CrossRef]
- Radhouani, H.; Igrejas, G.; Conçalves, A.; Pacheco, R.; Monteiro, R.; Sargo, R.; Brito, F.; Torres, C.; Poeta, P. Antimicrobial resistance and virulence genes in Escherichia coli and enterococci from red foxes (Vulpes vulpes). Anaerobe 2013, 23, 82–86. [Google Scholar] [CrossRef]
- Oliveira de Araujo, G.; Huff, R.; Favarini, M.O.; Mann, M.B.; Peters, F.B.; Frazzon, J.; Guedes Frazzon, A.P. Multidrug resistance in Enterococci isolated from wild Pampas foxes (Lycalopex gymnocercus) and Geoffroy’s cats (Leopardus geoffroyi) in the Brazilian Pampa Biome. Front. Vet. Sci. 2020, 7, 606377. [Google Scholar] [CrossRef] [PubMed]
- García, L.A.; Torres, C.; López, A.R.; Rodríguez, C.O.; Valencia, C.S. Antimicrobial resistance of Enterococcus species isolated from wild mammals in Aragón, Spain. J. Vet. Res. 2022, 66, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.; Hipólito, D.; Figueiredo, A.; Fonseca, C.; Caetano, T.; Mendo, S. Unravelling the diversity and abundance of the red fox (Vulpes vulpes) faecal resistome and their phenotypic antibiotic susceptibility of indicator bacteria. Animals 2022, 12, 2572. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Stępień-Pyśniak, D.; Gnat, S.; Fratini, F.; Urban-Chmiel, R.; Cerri, R.; Winiarczyk, S.; Turchi, B. Antibiotic susceptibility and virulence genes in Enterococcus isolates from wild mammals living in Tuscany, Italy. Microb. Drug Resist. 2020, 26, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Víchová, B.; Bona, M.; Miterpáková, M.; Kralijk, J.; Čabanová, V.; Nemčíková, G.; Hurníková, Z.; Oravec, M. Fleas and ticks of red foxes as vectors of canine bacterial and parasitic pathogens, in Slovakia, Central Europe. Vector Borne Zoonotic Dis. 2018, 18, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Central Statistical Bureau, Republic of Latvia. Usually Resident Population Density in Regions, Cities and Towns, Municipalities, and Rural Territories, at the Beginning of Year 2022–2023. Available online: https://data.stat.gov.lv/pxweb/en/OSP_PUB/START__POP__IR__IRD/IRD062/ (accessed on 1 December 2023).
- Busani, L.; Del Grosso, M.; Paladini, C.; Graziani, C.; Pantosti, A.; Biavasco, F.; Caprioli, A. Antimicrobial susceptibility of vancomycin-susceptible and -resistant enterococci isolated in Italy from raw meat products, farm animals, and human infections. Int. J. Food Microbiol. 2004, 97, 17–22. [Google Scholar] [CrossRef] [PubMed]
- EMA/299538/2023; Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2022. European Medicines Agency, European Surveillance of Veterinary Antimicrobial Consumption: Amsterdam, The Netherlands, 2022.
- Terentjeva, M.; Streikiša, M.; Avsejenko, J.; Trofimova, J.; Kovaļenko, K.; Elferts, D.; Bērziņš, A. Prevalence and antimicrobial resistance of Escherichia coli, Enterococcus spp. and the major foodborne pathogens in calves in Latvia. Foodborne Pathog. Dis. 2019, 16, 35–41. [Google Scholar] [CrossRef]
- Zhou, X.; Willems, R.J.L.; Friedrich, A.W.; Rossen, J.W.A.; Bathoorn, E. Enterococcus faecium: From microbiological insights to practical recommendations for infection control and diagnostics. Antimicrob. Resist. Infect. Control 2020, 9, 130. [Google Scholar] [CrossRef]
- Radhouani, H.; Igrejas, G.; Carvalho, C.; Pinto, L.; Gonçalves, A.; Lopez, M.; Sargo, R.; Cardoso, L.; Martinho, A.; Rego, V.; et al. Clonal lineages, antibiotic resistance and virulence factors in vancomycin-resistant Enterococci isolated from fecal samples of red foxes (Vulpes Vulpes). J. Wildl. Dis. 2011, 47, 769–773. [Google Scholar] [CrossRef]
- Cagnoli, G.; Bertelloni, F.; Interrante, P.; Ceccherelli, R.; Marzoni, M.; Ebani, V.V. Antimicrobial-resistant Enterococcus spp. in wild avifauna from central Italy. Antibiotics 2022, 11, 852. [Google Scholar] [CrossRef]
- Nowakiewicz, A.; Ziołkowska, G.; Zieba, P.; Kostruba, A. Undomesticated animals as a reservoir of multidrug resistant Enterococcus in eastern Poland. J. Wildl. Dis. 2014, 50, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef] [PubMed]
- del Campo, R.; Galán, J.C.; Tenorio, C.; Ruiz-Garbajosa, P.; Zarazaga, M.; Torres, C.; Baquero, F. New aac(6′)-I genes in Enterococcus hirae and Enterococcus durans: Effect on β-lactam/aminoglycoside synergy. J. Antimicrob. Chemother. 2005, 55, 1053–1055. [Google Scholar] [CrossRef] [PubMed]
- Semedo-Lamsaddek, T.; Nóbrega, C.S.; Ribeiro, T.; Pedroso, N.M.; Sales-Luís, T.; Lemsaddek, A.; Tenreiro, R.; Tavares, L.; Vilela, C.L.; Oliveira, M. Virulence traits and antibiotic resistance among enterococci isolated from Eurasian otter (Lutra lutra). Vet. Microbiol. 2013, 163, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Shete, V.; Grover, N.; Kumar, M. Analysis of aminoglycoside modifying enzyme genes responsible for high-level aminoglycoside resistance among Enterococcal isolates. J. Pathog. 2017, 64, 3256952. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Agerso, Y.; Smidt, P.G.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Huys, G.; D’Haene, K.; Collard, J.M.; Swings, J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl. Environ. Microbiol. 2004, 70, 1555–1562. [Google Scholar] [CrossRef]
- Roberts, A.P.; Mullany, P. A modular master on the move: The Tn916 family of mobile genetic elements. Trends Microbiol. 2009, 17, 251–258. [Google Scholar] [CrossRef]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid encoded tetracycline determinants tet(L) and tet(M). J. Antimicrob. Chemother. 2016, 71, 871–881. [Google Scholar] [CrossRef]
- Miller, W.R.; Murray, B.E.; Rice, L.B.; Arias, C.A. Resistance in vancomycin-resistant enterococci. Infect. Dis. Clin. N. Am. 2020, 34, 751–771. [Google Scholar] [CrossRef]
- Singh, K.V.; Malathum, K.; Murray, B.E. Disruption of an Enterococcus faecium species-specific gene, a homologue of acquired macrolide resistance genes of staphylococci, is associated with an increase in macrolide susceptibility. Antimicrob. Agents Chemother. 2001, 45, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.N.; Elleboudy, N.S.; El-Housseuny, G.S.; Yassien, M.A. Cloning and sequencing of lsaE efflux pump gene from MDR enterococci and its role in erythromycin resistance. Infect. Genet. Evol. 2021, 94, 105010. [Google Scholar] [CrossRef] [PubMed]
- Nowakiewicz, A.; Ziółkowska, G.; Zięba, P.; Gnat, S.; Trościańczyk, A.; Adaszek, Ł. Characterization of multidrug resistant E. faecalis strains from pigs of local origin by ADSRRS-fingerprinting and MALDI -TOF MS; evaluation of the compatibility of methods employed for multidrug resistance analysis. PLoS ONE 2017, 12, e0171160. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.S.; Lee, S.M.; Gan, H.M.; Dykes, G.A.; Rahman, S. Genetic diversity of Enterococcus faecalis isolated from environmental, animal and clinical sources in Malaysia. J. Infect. Public Health 2017, 10, 617–623. [Google Scholar] [CrossRef]
- Cabinets of Ministers. Cabinet Regulations No.178. Procedures for the Prevention and Combating Rabies. Available online: https://likumi.lv/ta/en/en/id/205795 (accessed on 1 December 2023).
- Food and Veterinary Service. State Program for Surveillance and Control of Rabies. 2021. Available online: https://www.pvd.gov.lv/lv/media/1109/download (accessed on 1 December 2023).
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical Breakpoints. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 1 December 2023).
- Mölder, F.; Jablonski, K.P.; Letcher, B.; Hall, M.B.; Tomkins-Tinch, C.H.; Sochat, V.; Forster, J.; Lee, S.; Twardziok, S.O.; Kanitz, A.; et al. Sustainable Data Analysis with Snakemake. F1000Res 2021, 10, 33. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 December 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Vasimuddin, M.; Misra, S.; Li, H.; Aluru, S. Efficient architecture-aware acceleration of BWA-MEM for multicore systems. In Proceedings of the 2019 IEEE International Parallel and Distributed Processing Symposium (IPDPS), Rio de Janeiro, Brazil, 20–24 May 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 314–324. [Google Scholar]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the reference gene catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; Von Haeseler, A.; Stoye, J.; et al. Updating Benchtop Sequencing Performance Comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Antimicrobial Agent | Range of Concentrations (μg/mL) | E. faecalis (n = 12) | E. faecium (n = 17) | E. durans (n = 3) | E. hirae (n = 2) | Resistance Threshold (μg/mL) |
|---|---|---|---|---|---|---|
| No. of Resistant Isolates (%) | ||||||
| Ampicillin | 0.5–64 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >4 |
| Chloramphenicol | 4–128 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >32 |
| Ciprofloxacin | 0.12–16 | 0 (0) | 2 (12) | 0 (0) | 0 (0) | >4 |
| Daptomycin | 0.25–32 | 0 (0) | 1 (6) | 0 (0) | 1 (50) | >4 |
| Erythromycin | 1–128 | 0 (0) | 3 (18) | 0 (0) | 0 (0) | >4 |
| Gentamicin | 8–1024 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >32 |
| Linezolid | 0.5–64 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >4 |
| Quinupristin/ dalfopristin | 0.5–64 | NA | 1 (6) | NA | NA | E. faecalis: NA, E. faecium: >4 |
| Teicoplanin | 0.5–64 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >2 |
| Tetracycline | 1–128 | 1 (8) | 4 (24) | 1 (33) | 0 (0) | >4 |
| Tigecycline | 0.03–4 | 0 (0) | 0 (0) | 1 (33) | 1 (50) | >0.25 |
| Vancomycin | 1–128 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | >4 |
| Enterococcus Species | Aminoglycosides | Tetracyclines | Macrolide–Lincosamide–Streptogramin AB | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| aac(6′)-I | aac(6′)-Iih | ant(6)-Ia | spw | tet(M) | tet(L) | tet(S) | lsa(A) | lsa(E) | msr(C) | lnu(B) | lnu(G) | |
| No. of Isolates (%) | ||||||||||||
| E. faecalis (n = 12) | 1 (8) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (8) | 11(92) | 0 (0) | 1 (8) | 0 (0) | 0 (0) |
| E. faecium (n = 17) | 16 (94) | 0 (0) | 2 (12) | 1(6) | 3 (18) | 1 (6) | 0 (0) | 1 (6) | 1 (6) | 16 (94) | 1 (6) | 1 (6) |
| E. durans (n = 3) | 2 (66) | 1 (33) | 0 (0) | 0 (0) | 1 (33) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| E. hirae (n = 2) | 1 (50) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Total | 20 (63) | 1 (3) | 2 (6) | 1(3) | 4 (13) | 1 (3) | 1 (3) | 12(34) | 1 (3) | 17 (53) | 1 (3) | 1 (3) |
| Isolate | Identified Phenotypic Resistance | Identified Antimicrobial Resistance Genes |
|---|---|---|
| 429441 E. faecium | Q/D8 | lsa(A) |
| 429441 E. faecalis | TET64 | aac(6′)-I, tet(S), msr(C) |
| 433771 E. faecium | TET64 | aac(6′)-I, msr(C), lnu(G), tet(M) |
| 3808 E. faecium | ERY8 | aac(6′)-I, msr(C) |
| 426383 E. faecium | TET64 | aac(6′)-I, msr(C), tet(M) |
| 428642/1 E. faecium | DAP8 | aac(6′)-I, msr(C) |
| 426804 E. faecium | CIP8 | aac(6′)-I, msr(C) |
| 428644 E. faecium | CIP8 | aac(6′)-I, msr(C) |
| 427166/2 E. durans | TET64 | aac(6′)-Iih, tet(M) |
| 23152 E. faecium | ERY8 | aac(6′)-I, msr(C) |
| 2123 E. faecium | TET128 | ant(6)-Ia, aac(6′)-I, lnu(B), lsa(E), msr(C), tet(L), tet(M), spw |
| 424211/2 E. faecium | TET>128, ERY64 | aac(6′)-I, msr(C) |
| 424365 E. hirae | TIG0.5 | aac(6′)-I |
| 421603/1 E. durans | TIG1 | aac(6′)-I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terentjeva, M.; Ķibilds, J.; Avsejenko, J.; Cīrulis, A.; Labecka, L.; Bērziņš, A. Antimicrobial Resistance in Enterococcus spp. Isolates from Red Foxes (Vulpes vulpes) in Latvia. Antibiotics 2024, 13, 114. https://doi.org/10.3390/antibiotics13020114
Terentjeva M, Ķibilds J, Avsejenko J, Cīrulis A, Labecka L, Bērziņš A. Antimicrobial Resistance in Enterococcus spp. Isolates from Red Foxes (Vulpes vulpes) in Latvia. Antibiotics. 2024; 13(2):114. https://doi.org/10.3390/antibiotics13020114
Chicago/Turabian StyleTerentjeva, Margarita, Juris Ķibilds, Jeļena Avsejenko, Aivars Cīrulis, Linda Labecka, and Aivars Bērziņš. 2024. "Antimicrobial Resistance in Enterococcus spp. Isolates from Red Foxes (Vulpes vulpes) in Latvia" Antibiotics 13, no. 2: 114. https://doi.org/10.3390/antibiotics13020114
APA StyleTerentjeva, M., Ķibilds, J., Avsejenko, J., Cīrulis, A., Labecka, L., & Bērziņš, A. (2024). Antimicrobial Resistance in Enterococcus spp. Isolates from Red Foxes (Vulpes vulpes) in Latvia. Antibiotics, 13(2), 114. https://doi.org/10.3390/antibiotics13020114