
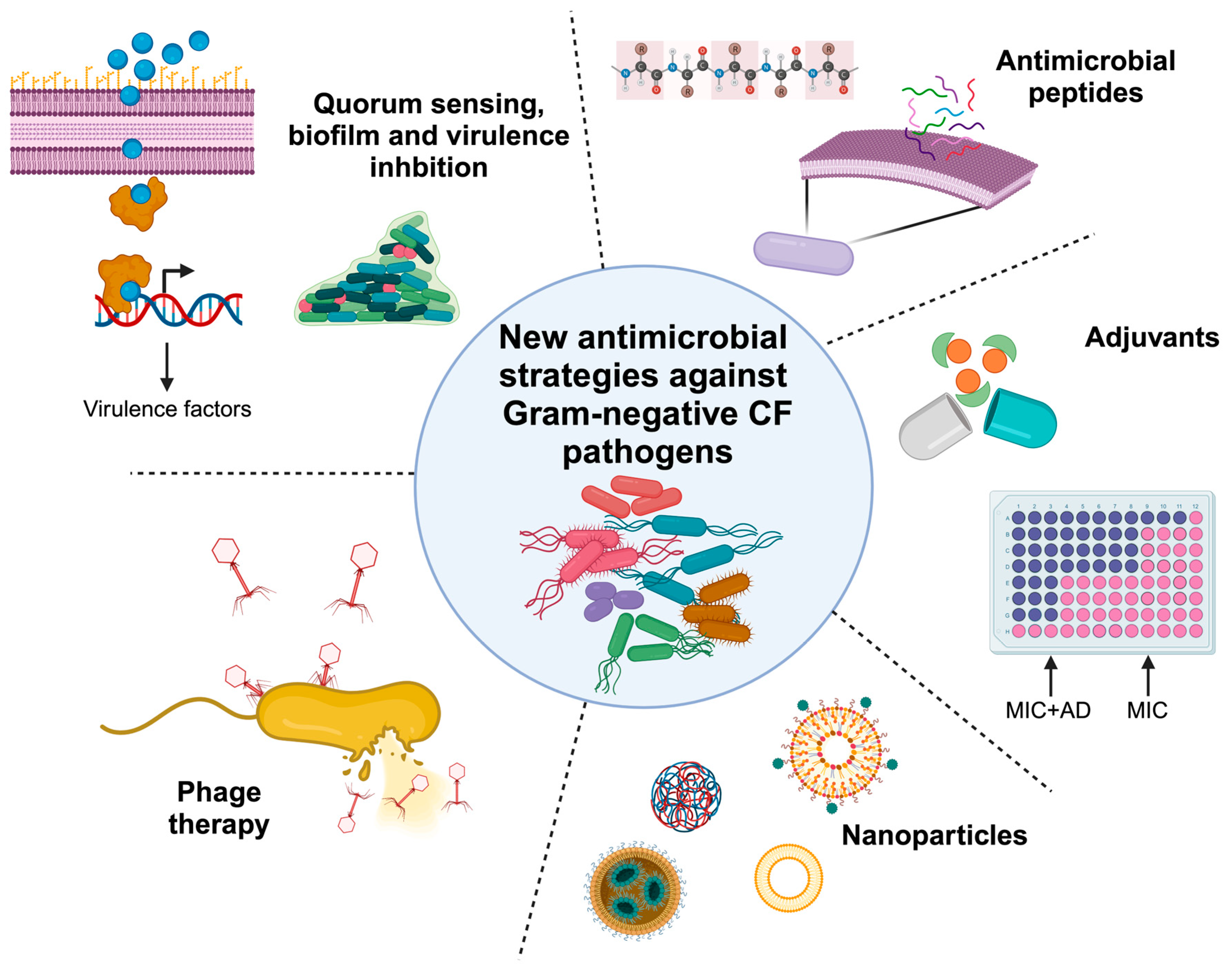
New Antimicrobial Strategies to Treat Multi-Drug Resistant Infections Caused by Gram-Negatives in Cystic Fibrosis


Abstract
1. Introduction
2. Virulence and Biofilm Inhibition
2.1. Synthetic Molecules
2.1.1. Quorum Sensing Inhibitors
2.1.2. Biofilm Inhibitors

{kind=link}
{kind=link}
{kind=link}
| Synthetic Molecules | ||
|---|---|---|
| Biofilm Inhibitors | Bacteria | Reference |
| N-acetylcysteine (NAC) | P. aeruginosa, Bcc and S. maltophilia | [22,23,24] |
| Nitric oxide donors, sodium nitroprusside | P. aeruginosa | [26,27] |
| Oligo G | P. aeruginosa, H. influenzae | [30,31,32] |
| Deflazacort, PYED-1 | S. maltophilia | [33] |
2.1.3. Anti-Virulence Molecules
2.2. Natural Compounds
2.2.1. Quorum Sensing Inhibitors
2.2.2. Biofilm Inhibitors
3. Phage Therapy
3.1. H. influenzae
3.2. A. xylosoxidans
3.3. B. cepacia Complex
3.4. S. maltophilia
3.5. P. aeruginosa
4. Antibiotic Adjuvants
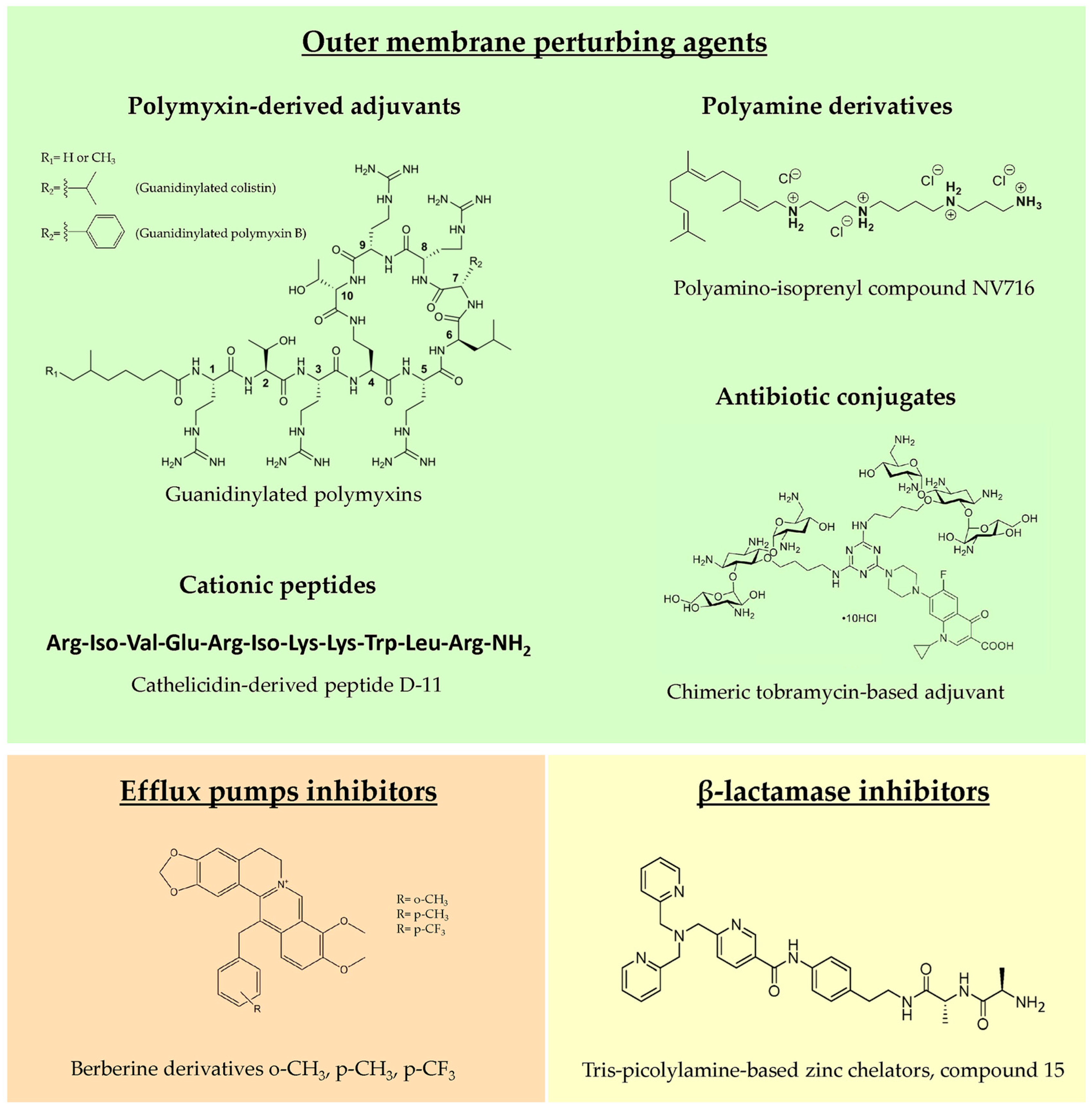
4.1. Outer Membrane Perturbing Agents
4.1.1. Polymyxin-Derived Adjuvants
4.1.2. Polyamine Derivatives
4.1.3. Cationic Peptides
4.1.4. Antibiotic Conjugates
4.2. Efflux Pump Inhibitors
4.3. β-lactamase Inhibitors
4.4. Other Adjuvants
5. Antimicrobial Peptides
5.1. Natural Antimicrobial Peptides
5.2. Strategies to Overcome the Drawbacks of the Use of Antimicrobial Peptides In Vivo
5.3. Antimicrobial Peptides in Clinical Trials for Cystic Fibrosis Infections
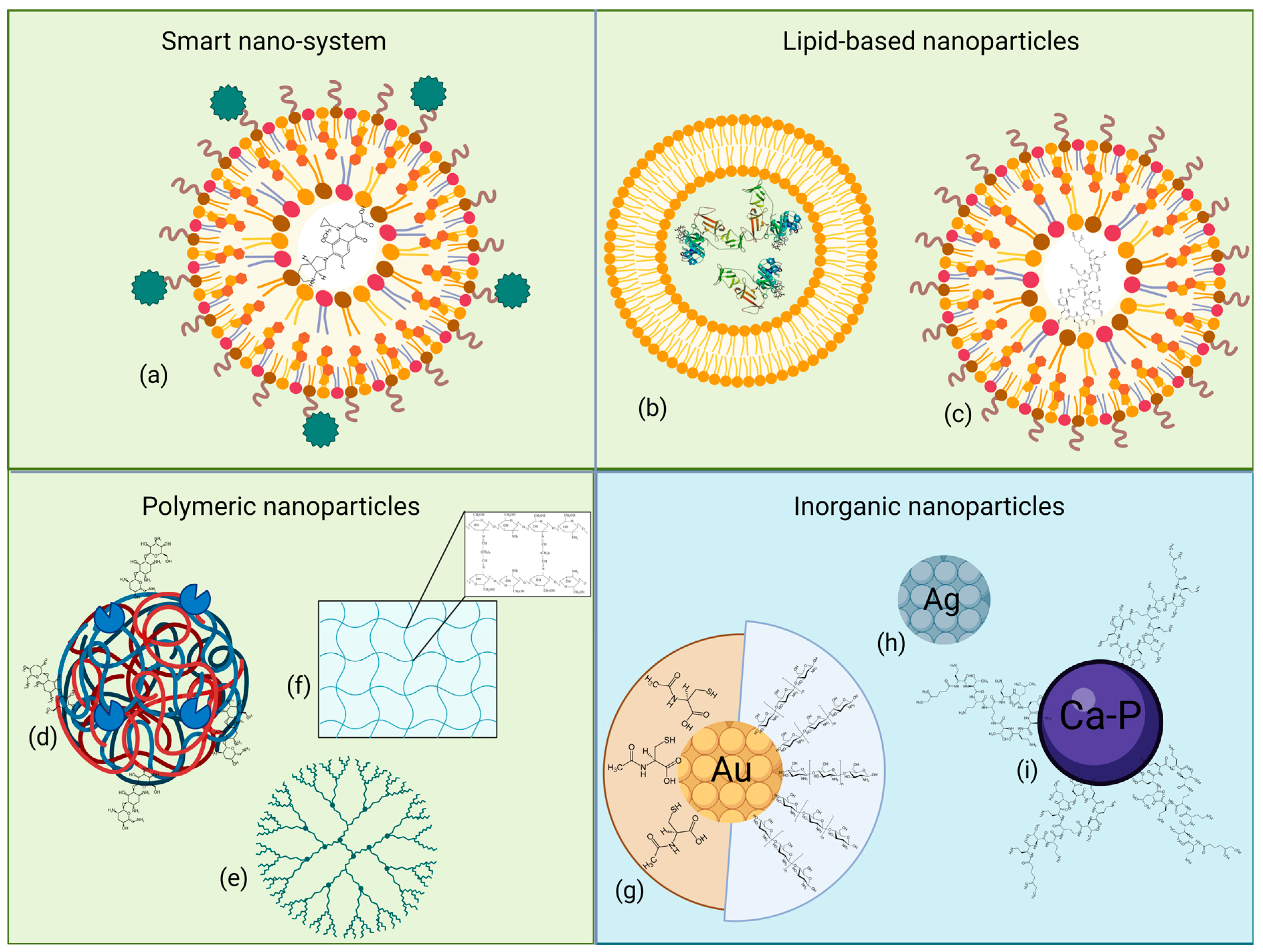
6. Nanoparticles
6.1. Organic Nanoparticles
6.1.1. Smart Nano-Systems
6.1.2. Lipid-Based Nanoparticles
6.1.3. Polymeric Nanoparticles
6.2. Inorganic Nanoparticles
7. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Morrison, C.B.; Markovetz, M.R.; Ehre, C. Mucus, mucins, and cystic fibrosis. Pediatr. Pulmonol. 2019, 54 (Suppl. S3), S84–S96. [Google Scholar] [CrossRef] [PubMed]
- Françoise, A.; Héry-Arnaud, G. The Microbiome in Cystic Fibrosis Pulmonary Disease. Genes 2020, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Perikleous, E.P.; Gkentzi, D.; Bertzouanis, A.; Paraskakis, E.; Sovtic, A.; Fouzas, S. Antibiotic Resistance in Patients with Cystic Fibrosis: Past, Present, and Future. Antibiotics 2023, 12, 217. [Google Scholar] [CrossRef] [PubMed]
- Munguia, J.; Nizet, V. Pharmacological Targeting of the Host-Pathogen Interaction: Alternatives to Classical Antibiotics to Combat Drug-Resistant Superbugs. Trends Pharmacol. Sci. 2017, 38, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Trespidi, G.; Chiarelli, L.R.; Barbieri, G.; Buroni, S. Quorum Sensing as Antivirulence Target in Cystic Fibrosis Pathogens. Int. J. Mol. Sci. 2019, 20, 1838. [Google Scholar] [CrossRef]
- Ballante, F.; Turkina, M.V.; Ntzouni, M.; Magnusson, K.E.; Vikström, E. Modified N-acyl-L-homoserine lactone compounds abrogate Las-dependent quorum-sensing response in human pathogen Pseudomonas aeruginosa. Front. Mol. Biosci. 2023, 10, 1264773. [Google Scholar] [CrossRef]
- Chang, Y.; Wang, P.C.; Ma, H.M.; Chen, S.Y.; Fu, Y.H.; Liu, Y.Y.; Wang, X.; Yu, G.C.; Huang, T.; Hibbs, D.E.; et al. Design, synthesis and evaluation of halogenated furanone derivatives as quorum sensing inhibitors in Pseudomonas aeruginosa. Eur. J. Pharm. Sci. 2019, 140, 105058. [Google Scholar] [CrossRef]
- Gómez, A.C.; Lyons, T.; Mamat, U.; Yero, D.; Bravo, M.; Daura, X.; Elshafee, O.; Brunke, S.; Gahan, C.G.M.; O’Driscoll, M.; et al. Synthesis and evaluation of novel furanones as biofilm inhibitors in opportunistic human pathogens. Eur. J. Med. Chem. 2022, 242, 114678. [Google Scholar] [CrossRef]
- Carullo, G.; Di Bonaventura, G.; Rossi, S.; Lupetti, V.; Tudino, V.; Brogi, S.; Butini, S.; Campiani, G.; Gemma, S.; Pompilio, A. Deelopment of Quinazolinone Derivatives as Modulators of Virulence Factors of Pseudomonas aeruginosa Cystic Fibrosis Strains. Molecules 2023, 28, 6535. [Google Scholar] [CrossRef]
- Miao, Z.Y.; Zhang, X.Y.; Yang, M.H.; Huang, Y.J.; Lin, J.; Chen, W.M. 3-Hydroxypyridin-4[1H]-one Derivatives as pqs Quorum Sensing Inhibitors Attenuate Virulence and Reduce Antibiotic Resistance in Pseudomonas aeruginosa. J. Med. Chem. 2023, 66, 15823–15846. [Google Scholar] [CrossRef] [PubMed]
- Schütz, C.; Ho, D.K.; Hamed, M.M.; Abdelsamie, A.S.; Röhrig, T.; Herr, C.; Kany, A.M.; Rox, K.; Schmelz, S.; Siebenbürger, L.; et al. A New PqsR Inverse Agonist Potentiates Tobramycin Efficacy to Eradicate Pseudomonas aeruginosa Biofilms. Adv. Sci. 2021, 8, e2004369. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, V.; D’Angelo, F.; Pavoncello, V.; Fiscarelli, E.V.; Visca, P.; Rampioni, G.; Leoni, L. Identification of FDA-approved antivirulence drugs targeting the Pseudomonas aeruginosa quorum sensing effector protein PqsE. Virulence 2020, 11, 652–668. [Google Scholar] [CrossRef]
- Cornforth, D.M.; Dees, J.L.; Ibberson, C.B.; Huse, H.K.; Mathiesen, I.H.; Kirketerp-Møller, K.; Wolcott, R.D.; Rumbaugh, K.P.; Bjarnsholt, T.; Whiteley, M. Pseudomonas aeruginosa transcriptome during human infection. Proc. Natl. Acad. Sci. USA 2018, 115, E5125–E5134. [Google Scholar] [CrossRef] [PubMed]
- Collalto, D.; Giallonardi, G.; Fortuna, A.; Meneghini, C.; Fiscarelli, E.; Visca, P.; Imperi, F.; Rampioni, G.; Leoni, L. In vitro Activity of Antivirulence Drugs Targeting the las or pqs Quorum Sensing Against Cystic Fibrosis Pseudomonas aeruginosa Isolates. Front. Microbiol. 2022, 13, 845231. [Google Scholar] [CrossRef]
- Imperi, F.; Fiscarelli, E.V.; Visaggio, D.; Leoni, L.; Visca, P. Activity and Impact on Resistance Development of Two Antivirulence Fluoropyrimidine Drugs in Pseudomonas aeruginosa. Front. Cell Infect. Microbiol. 2019, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Hibbert, T.M.; Whiteley, M.; Renshaw, S.A.; Neill, D.R.; Fothergill, J.L. Emerging strategies to target virulence in Pseudomonas aeruginosa respiratory infections. Crit. Rev. Microbiol. 2023, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, Y.; Xiang, S.; Xu, K.; Wang, B.; Jia, A.Q. Tyramine, one quorum sensing inhibitor, reduces pathogenicity and restores tetracycline susceptibility in Burkholderia cenocepacia. Biochem. Pharmacol. 2023, 218, 115906. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Chiarelli, L.R.; Makarov, V.; Brackman, G.; Israyilova, A.; Azzalin, A.; Forneris, F.; Riabova, O.; Savina, S.; Coenye, T.; et al. Discovery of new diketopiperazines inhibiting Burkholderia cenocepacia quorum sensing in vitro and in vivo. Sci. Rep. 2016, 6, 32487. [Google Scholar] [CrossRef]
- Buroni, S.; Scoffone, V.C.; Fumagalli, M.; Makarov, V.; Cagnone, M.; Trespidi, G.; De Rossi, E.; Forneris, F.; Riccardi, G.; Chiarelli, L.R. Investigating the Mechanism of Action of Diketopiperazines Inhibitors of the Burkholderia cenocepacia Quorum Sensing Synthase CepI: A Site-Directed Mutagenesis Study. Front. Pharmacol. 2018, 9, 836. [Google Scholar] [CrossRef]
- Huedo, P.; Kumar, V.P.; Horgan, C.; Yero, D.; Daura, X.; Gibert, I.; O’Sullivan, T.P. Sulfonamide-based diffusible signal factor analogs interfere with quorum sensing in Stenotrophomonas maltophilia and Burkholderia cepacia. Future Med. Chem. 2019, 11, 1565–1582. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Liu, Y. N-acetylcysteine inhibit biofilms produced by Pseudomonas aeruginosa. BMC Microbiol. 2010, 10, 140. [Google Scholar] [CrossRef]
- Aiyer, A.; Visser, S.K.; Bye, P.; Britton, W.J.; Whiteley, G.S.; Glasbey, T.; Kriel, F.H.; Farrell, J.; Das, T.; Manos, J. Effect of N-acetylcysteine in combination with antibiotics on the biofilms of three cystic fibrosis pathogens of emerging importance. Antibiotics 2021, 10, 1176. [Google Scholar] [CrossRef] [PubMed]
- Pollini, S.; Di Pilato, V.; Landini, G.; Di Maggio, T.; Cannatelli, A.; Sottotetti, S.; Cariani, L.; Aliberti, S.; Blasi, F.; Sergio, F.; et al. In vitro activity of N-acetylcysteine against Stenotrophomonas maltophilia and Burkholderia cepacia complex grown in planktonic phase and biofilm. PLoS ONE 2018, 13, e0203941. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, F.N.; Hacioglu, M.; Aldogan, E.H. Impact of N-acetylcysteine and antibiotics against single and dual species biofilms of Pseudomonas aeruginosa and Achromobacter xylosoxidans. Curr. Microbiol. 2022, 80, 5. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.B.; Kragh, K.N.; Hultqvist, L.D.; Rybtke, M.; Nilsson, M.; Jakobsen, T.H.; Givskov, M.; Tolker-Nielsen, T. Induction of Native c-di-GMP Phosphodiesterases Leads to Dispersal of Pseudomonas aeruginosa Biofilms. Antimicrob. Agents Chemother. 2021, 65, e02431-20. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.M.; Webb, J.S. Optimization of nitric oxide donors for investigating biofilm dispersal response in Pseudomonas aeruginosa clinical isolates. Appl. Microbiol. Biotechnol. 2020, 104, 8859–8869. [Google Scholar] [CrossRef]
- Sorbo, L.D.; Michaelsen, V.S.; Ali, A.; Wang, A.; Ribeiro, R.V.P.; Cypel, M. High Doses of Inhaled Nitric Oxide as an Innovative Antimicrobial Strategy for Lung Infections. Biomedicines 2022, 10, 1525. [Google Scholar] [CrossRef]
- Daboor, S.M.; Rohde, J.R.; Cheng, Z. Disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate lyase enhances pathogen eradication by antibiotics. J. Cyst. Fibros. 2021, 20, 264–270. [Google Scholar] [CrossRef]
- Marienborg, K.; Ambur, O.H.; Økstad, O.A.L.; Skaare, D. The alginate polymer OligoG alters susceptibility of biofilm-embedded non-typeable Haemophilus influenzae to ampicillin and ciprofloxacin. JAC Antimicrob. Resist. 2023, 5, dlad046. [Google Scholar] [CrossRef]
- Powell, L.C.; Pritchard, M.F.; Ferguson, E.L.; Powell, K.A.; Patel, S.U.; Rye, P.D.; Sakellakou, S.M.; Buurma, N.J.; Brilliant, C.D.; Copping, J.M.; et al. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. NPJ Biofilms Microbiomes 2018, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- van Koningsbruggen-Rietschel, S.; Davies, J.C.; Pressler, T.; Fischer, R.; MacGregor, G.; Donaldson, S.H.; Smerud, K.; Meland, N.; Mortensen, J.; Fosbøl, M.Ø.; et al. Inhaled dry powder alginate oligosaccharide in cystic fibrosis: A randomised, double-blind, placebo-controlled, crossover phase 2b study. ERJ Open Res. 2020, 6, 00132-2020. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.; Vollaro, A.; Esposito, E.P.; D’Alonzo, D.; Guaragna, A.; Zarrilli, R.; De Gregorio, E. Antibacterial and Antivirulence Activity of Glucocorticoid PYED-1 against Stenotrophomonas maltophilia. Antibiotics 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Oluwabusola, E.T.; Katermeran, N.P.; Poh, W.H.; Goh, T.M.B.; Tan, L.T.; Diyaolu, O.; Tabudravu, J.; Ebel, R.; Rice, S.A.; Jaspars, M. Inhibition of the Quorum Sensing System, Elastase Production and Biofilm Formation in Pseudomonas aeruginosa by Psammaplin A and Bisaprasin. Molecules 2022, 27, 1721. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, P.; Guo, T.; Gu, X.; Bai, B.; Zhang, S.; Chang, X.; Wang, Y.; Ma, S. Design, synthesis and evaluation of oxazolopyridinone derivatives as quorum sensing inhibitors. Bioorg Chem. 2023, 130, 106266. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Revtovich, A.V.; Deyanov, A.E.; Kirienko, N.V. Pyoverdine Inhibitors and Gallium Nitrate Synergistically Affect Pseudomonas aeruginosa. mSphere 2021, 6, e0040121. [Google Scholar] [CrossRef]
- Ali, S.O.; Yu, X.Q.; Robbie, G.J.; Wu, Y.; Shoemaker, K.; Yu, L.; DiGiandomenico, A.; Keller, A.E.; Anude, C.; Hernandez-Illas, M.; et al. Phase 1 study of MEDI3902, an investigational anti-Pseudomonas aeruginosa PcrV and Psl bispecific human monoclonal antibody, in healthy adults. Clin. Microbiol. Infect. 2019, 25, e1–e629. [Google Scholar] [CrossRef]
- Sheremet, A.B.; Zigangirova, N.A.; Zayakin, E.S.; Luyksaar, S.I.; Kapotina, L.N.; Nesterenko, L.N.; Kobets, N.V.; Gintsburg, A.L. Small Molecule Inhibitor of Type Three Secretion System Belonging to a Class 2,4-disubstituted-4H-[1,3,4]-thiadiazine-5-ones Improves Survival and Decreases Bacterial Loads in an Airway Pseudomonas aeruginosa Infection in Mice. Biomed Res. Int. 2018, 2018, 5810767. [Google Scholar] [CrossRef]
- Shehaj, L.; Choudary, S.K.; Makwana, K.M.; Gallo, M.C.; Murphy, T.F.; Kritzer, J.A. Small-Molecule Inhibitors of Haemophilus influenzae IgA1 Protease. ACS Infect. Dis. 2019, 5, 1129–1138. [Google Scholar] [CrossRef]
- Bové, M.; Coenye, T. The anti-virulence activity of the non-mevalonate pathway inhibitor FR900098 towards Burkholderia cenocepacia is maintained during experimental evolution. Microbiology 2022, 168, 1770. [Google Scholar] [CrossRef]
- Zhang, Y.; Sass, A.; Van Acker, H.; Wille, J.; Verhasselt, B.; Van Nieuwerburgh, F.; Kaever, V.; Crabbé, A.; Coenye, T. Coumarin Reduces Virulence and Biofilm Formation in Pseudomonas aeruginosa by Affecting Quorum Sensing, Type III Secretion and C-di-GMP Levels. Front. Microbiol. 2018, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, S.Y.; Hu, J.Y.; Chen, Q.X.; Jiao, S.M.; Xiao, H.C.; Zhang, Q.; Xu, J.; Zhao, J.F.; Zhou, H.B.; et al. Novel Coumarin Derivatives Inhibit the Quorum Sensing System and Iron Homeostasis as Antibacterial Synergists against Pseudomonas aeruginosa. J. Med. Chem. 2023, 66, 14735–14754. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Dong, B.; Wang, K.; Cai, S.; Liu, T.; Cheng, X.; Lei, D.; Chen, Y.; Li, Y.; Kong, J.; et al. Baicalin inhibits biofilm formation, attenuates the quorum sensing-controlled virulence and enhances Pseudomonas aeruginosa clearance in a mouse peritoneal implant infection model. PLoS ONE 2017, 12, e0176883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Guo, Q.; Wei, Z.; Yang, Q.; Guo, Z.; Shen, L.; Duan, K.; Chen, L. Baicalin Represses Type Three Secretion System of Pseudomonas aeruginosa through PQS System. Molecules 2021, 26, 1497. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.; Slachmuylders, L.; Van Acker, H.; Vandenbussche, I.; Ostyn, L.; Bové, M.; Crabbé, A.; Chiarelli, L.R.; Buroni, S.; Van Nieuwerburgh, F.; et al. Various Evolutionary Trajectories Lead to Loss of the Tobramycin-Potentiating Activity of the Quorum-Sensing Inhibitor Baicalin Hydrate in Burkholderia cenocepacia Biofilms. Antimicrob. Agents Chemother. 2019, 63, e02092-18. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, K.; Wang, G.; Cui, B.; Song, S.; Sun, X.; Deng, Y. Oridonin Attenuates Burkholderia cenocepacia Virulence by Suppressing Quorum-Sensing Signaling. Microbiol. Spectr. 2022, 10, e0178722. [Google Scholar] [CrossRef]
- Ghosh, D.; Seth, M.; Mondal, P.; Mukhopadhyay, S.K. Biocontrol of biofilm forming Burkholderia cepacia using a quorum quenching crude lactonase enzyme extract from a marine Chromohalobacter sp. strain D23. Arch. Microbiol. 2023, 205, 374. [Google Scholar] [CrossRef]
- Kim, H.R.; Lee, D.; Eom, Y.B. Anti-biofilm and Anti-Virulence Efficacy of Celastrol Against Stenotrophomonas maltophilia. Int. J. Med. Sci. 2018, 15, 617–627. [Google Scholar] [CrossRef]
- Elekhnawy, E.; Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Alqarni, M.; Batiha, G.E.; Obaidullah, A.J.; Fawzy, H.M. Histological assessment, anti-quorum sensing, and anti-biofilm activities of Dioon spinulosum extract: In vitro and in vivo approach. Sci. Rep. 2022, 12, 180. [Google Scholar] [CrossRef]
- Lawrence, J.A.; Huang, Z.; Rathinavelu, S.; Hu, J.F.; Garo, E.; Ellis, M.; Norman, V.L.; Buckle, R.; Williams, R.B.; Starks, C.M.; et al. Optimized plant compound with potent anti-biofilm activity across gram-negative species. Bioorg Med. Chem. 2020, 28, 115229. [Google Scholar] [CrossRef]
- Balázs, V.L.; Filep, R.; Répás, F.; Kerekes, E.; Szabó, P.; Kocsis, B.; Böszörményi, A.; Krisch, J.; Horváth, G. Immortelle [Helichrysum italicum [Roth] G. Don] Essential Oil Showed Antibacterial and Biofilm Inhibitory Activity against Respiratory Tract Pathogens. Molecules 2022, 27, 5518. [Google Scholar] [CrossRef] [PubMed]
- Piasecki, B.; Balázs, V.L.; Kieltyka-Dadasiewicz, A.; Szabó, P.; Kocsis, B.; Horváth, G.; Ludwiczuk, A. Microbiological Studies on the Influence of Essential Oils from Several Origanum Species on Respiratory Pathogens. Molecules 2023, 28, 3044. [Google Scholar] [CrossRef] [PubMed]
- Bakó, C.; Balázs, V.L.; Kerekes, E.; Kocsis, B.; Nagy, D.U.; Szabó, P.; Micalizzi, G.; Mondello, L.; Krisch, J.; Pethő, D.; et al. Flowering phenophases influence the antibacterial and anti-biofilm effects of Thymus vulgaris L. essential oil. BMC Complement. Med. Ther. 2023, 23, 168. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.O.; Zedan, H.H.; Ibrahim, Y.M. Quorum sensing inhibitory effect of bergamot oil and aspidosperma extract against Chromobacterium violaceum and Pseudomonas aeruginosa. Arch. Microbiol. 2021, 203, 4663–4675. [Google Scholar] [CrossRef]
- Damar-Çelik, D.; Mataracı-Kara, E.; Savage, P.B.; Özbek-Çelik, B. Antibacterial and antibiofilm activities of ceragenins against Achromobacter species isolated from cystic fibrosis patients. J. Chemother. 2021, 33, 216–227. [Google Scholar] [CrossRef]
- Peters, M.K.; Astafyeva, Y.; Han, Y.; Macdonald, J.F.H.; Indenbirken, D.; Nakel, J.; Virdi, S.; Westhoff, G.; Streit, W.R.; Krohn, I. Novel marine metalloprotease-new approaches for inhibition of biofilm formation of Stenotrophomonas maltophilia. Appl. Microbiol. Biotechnol. 2023, 107, 7119–7134. [Google Scholar] [CrossRef]
- ElBaradei, A.; Yakout, M.A. Stenotrophomonas maltophilia: Genotypic Characterization of Virulence Genes and The Effect of Ascorbic Acid on Biofilm Formation. Curr. Microbiol. 2022, 79, 180. [Google Scholar] [CrossRef]
- Karunanidhi, A.; Ghaznavi-Rad, E.; Hamat, R.A.; Pichika, M.R.; Lung, L.T.T.; Mohd Fauzi, F.; Chigurupati, S.; van Belkum, A.; Neela, V. Antibacterial and Antibiofilm Activities of Nonpolar Extracts of Allium stipitatum Regel. against Multidrug Resistant Bacteria. Biomed Res. Int. 2018, 2018, 9845075. [Google Scholar] [CrossRef]
- Wang, D.; Naqvi, S.T.A.; Lei, F.; Zhang, Z.; Yu, H.; Ma, L.Z. Glycosyl hydrolase from Pseudomonas fluorescens inhibits the biofilm formation of Pseudomonads. Biofilm 2023, 6, 100155. [Google Scholar] [CrossRef]
- Paul, V.D.; Sundarrajan, S.; Rajagopalan, S.S.; Hariharan, S.; Kempashanaiah, N.; Padmanabhan, S.; Sriram, B.; Ramachandran, J. Lysis-deficient phages as novel therapeutic agents for controlling bacterial infection. BMC Microbiol. 2011, 11, 195. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, N.M.; Nunes, D.D.G.; Shiach, J.; Hodel, K.V.S.; Barbosa, J.D.V.; Rodrigues, L.A.P.; Coler, B.S.; Soares, M.B.P.; Badaró, R. Current clinical landscape and global potential of bacteriophage therapy. Viruses 2023, 15, 1020. [Google Scholar] [CrossRef]
- Pirnay, J.P.; Kutter, E. Bacteriophages: It’s a medicine, Jim, but not as we know it. Lancet Infect. Dis. 2021, 21, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Kaabi, S.A.G.; Musafer, H.K. An experimental mouse model for phage therapy of bacterial pathogens causing bacteremia. Microb. Pathog. 2019, 137, 103770. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, N.; Zhvaniya, P.; Balarjishvili, N.; Bolkvadze, D.; Nadareishvili, L.; Nizharadze, D.; Wittmann, J.; Rohde, C.; Kutateladze, M. Phage therapy against Achromobacter xylosoxidans lung infection in a patient with cystic fibrosis: A case report. Res. Microbiol. 2018, 169, 540–542. [Google Scholar] [CrossRef]
- Gainey, A.B.; Burch, A.K.; Brownstein, M.J.; Brown, D.E.; Fackler, J.; Horne, B.; Biswas, B.; Bivens, B.N.; Malagon, F.; Daniels, R. Combining bacteriophages with cefiderocol and meropenem/vaborbactam to treat a pan-drug resistant Achromobacter species infection in a pediatric cystic fibrosis patient. Pediatr. Pulmonol. 2020, 55, 2990–2994. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, D.; Merabishvili, M.; Caudron, E.; Lannoy, D.; Van Simaey, L.; Duyvejonck, H.; Guillemain, R.; Thumerelle, C.; Podglajen, I.; Compain, F.; et al. A case of phage therapy against pandrug-resistant Achromobacter xylosoxidans in a 12-year-old lung-transplanted cystic fibrosis patient. Viruses 2021, 13, 60. [Google Scholar] [CrossRef]
- Cobián Güemes, A.G.; Le, T.; Rojas, M.I.; Jacobson, N.E.; Villela, H.; McNair, K.; Hung, S.H.; Han, L.; Boling, L.; Octavio, J.C.; et al. Compounding Achromobacter phages for therapeutic applications. Viruses 2023, 15, 1665. [Google Scholar] [CrossRef]
- Godoy, B.; Yao, G.; Le, T.; Vizoso-Pinto, M.G.; Gill, J.; Gonzalez, C.; Liu, M. Complete genome sequence of Burkholderia gladioli Myophage Mana. Microbiol. Resour. Announc. 2021, 10, e00402-21. [Google Scholar] [CrossRef]
- Davis, C.M.; Ruest, M.K.; Cole, J.H.; Dennis, J.J. The Isolation and characterization of a broad host range Bcep22-like podovirus JC1. Viruses 2022, 14, 938. [Google Scholar] [CrossRef]
- Lauman, P.; Dennis, J.J. Synergistic Interactions among Burkholderia cepacia Complex-Targeting Phages Reveal a Novel Therapeutic Role for Lysogenization-Capable Phages. Microbiol. Spectr. 2023, 11, e0443022. [Google Scholar] [CrossRef]
- Mankovich, A.G.; Maciel, K.; Kavanaugh, M.; Kistler, E.; Muckle, E.; Weingart, C.L. Phage-antibiotic synergy reduces Burkholderia cenocepacia population. BMC Microbiol. 2023, 23, 2. [Google Scholar] [CrossRef]
- Ruest, M.K.; Supina, B.S.I.; Dennis, J.J. Bacteriophage steering of Burkholderia cenocepacia toward reduced virulence and increased antibiotic sensitivity. J. Bacteriol. 2023, 205, e0019623. [Google Scholar] [CrossRef]
- Haidar, G.; Chan, B.K.; Cho, S.T.; Hughes Kramer, K.; Nordstrom, H.R.; Wallace, N.R.; Stellfox, M.E.; Holland, M.; Kline, E.G.; Kozar, J.M.; et al. Phage therapy in a lung transplant recipient with cystic fibrosis infected with multidrug-resistant Burkholderia multivorans. Transpl. Infect. Dis. 2023, 25, e14041. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.G.; Lin, A.; Dennis, J.J. Isolation and characterization of the novel bacteriophage AXL3 against Stenotrophomonas maltophilia. Int. J. Mol. Sci. 2020, 21, 6338. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.G.; Lin, A.; Dennis, J.J. Characterization of Stenotrophomonas maltophilia phage AXL1 as a member of the genus Pamexvirus encoding resistance to trimethoprim-sulfamethoxazole. Sci. Rep. 2022, 12, 10299. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.L.; McCutcheon, J.G.; Dennis, J.J. Characterization of Novel Broad-Host-Range Bacteriophage DLP3 Specific to Stenotrophomonas maltophilia as a Potential Therapeutic Agent. Front. Microbiol. 2020, 11, 1358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, R.; Hu, Y.; Liu, Y.; Wang, L.; An, X.; Song, L.; Shi, T.; Fan, H.; Tong, Y.; et al. Biological characteristics and genomic analysis of a Stenotrophomonas maltophilia phage vB_SmaS_BUCT548. Virus Genes. 2021, 57, 205–216. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Liu, X.; Ning, H.; Lin, H.; Wang, J. Biological characterization and complete genome analysis of a novel Stenotrophomonas maltophilia phage vB_SM_ytsc_ply2008005c. Virus Res. 2022, 318, 198856. [Google Scholar] [CrossRef]
- Han, K.; He, X.; Fan, H.; Song, L.; An, X.; Li, M.; Tong, Y. Characterization and genome analysis of a novel Stenotrophomonas maltophilia bacteriophage BUCT598 with extreme pH resistance. Virus Res. 2022, 314, 198751. [Google Scholar] [CrossRef]
- Han, P.; Zhang, W.; Pu, M.; Li, Y.; Song, L.; An, X.; Li, M.; Li, F.; Zhang, S.; Fan, H.; et al. Characterization of the bacteriophage BUCT603 and therapeutic potential evaluation against drug-resistant Stenotrophomonas maltophilia in a mouse model. Front. Microbiol. 2022, 13, 906961. [Google Scholar] [CrossRef]
- Li, Y.; Pu, M.; Han, P.; Li, M.; An, X.; Song, L.; Fan, H.; Chen, Z.; Tong, Y. Efficacy in Galleria mellonella larvae and application potential assessment of a new bacteriophage BUCT700 extensively lyse Stenotrophomonas maltophilia. Microbiol. Spectr. 2023, 11, e0403022. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Cheng, Y.Y.; Deng, J.; Cai, Z.; Zhong, L.; Qu, J.X.; Wang, K.; Yang, L. Genomic insights into the phage-defense systems of Stenotrophomonas maltophilia clinical isolates. Microbiol. Res. 2024, 278, 127528. [Google Scholar] [CrossRef] [PubMed]
- Koshy, C.M.; Sugumar, S. Isolation, characterization, and genome analysis of novel bacteriophage—Stenotrophomonas phage CM1. Microb. Pathog. 2023, 185, 106403. [Google Scholar] [CrossRef]
- Lin, Y.M.; Chang, R.Y.K.; Britton, W.J.; Morales, S.; Kutter, E.; Li, J.; Chan, H.K. Inhalable combination powder formulations of phage and ciprofloxacin for P. aeruginosa respiratory infections. Eur. J. Pharm. Biopharm. 2019, 142, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Luscher, A.; Simonin, J.; Falconnet, L.; Valot, B.; Hocquet, D.; Chanson, M.; Resch, G.; Köhler, T.; van Delden, C. Combined bacteriophage and antibiotic treatment prevents Pseudomonas aeruginosa infection of wild type and cftr- epithelial cells. Front. Microbiol. 2020, 11, 1947. [Google Scholar] [CrossRef] [PubMed]
- Raz, A.; Serrano, A.; Hernandez, A.; Euler, C.W.; Fischetti, V.A. Isolation of phage lysins that effectively kill Pseudomonas aeruginosa in mouse models of lung and skin infection. Antimicrob. Agents Chemother. 2019, 63, e00024-19. [Google Scholar] [CrossRef] [PubMed]
- Cafora, M.; Deflorian, G.; Forti, F.; Ferrari, L.; Binelli, G.; Briani, F.; Ghisotti, D.; Pistocchi, A. Phage therapy against Pseudomonas aeruginosa infections in a cystic fibrosis zebrafish model. Sci. Rep. 2019, 9, 1527. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.M.; Cai, A.; Chen, A.; Gittrich, M.R.; Sullivan, M.B.; Wozniak, D.J.; Abedon, S.T. Friends or foes? rapid determination of dissimilar colistin and ciprofloxacin antagonism of Pseudomonas aeruginosa phages. Pharmaceuticals 2021, 14, 1162. [Google Scholar] [CrossRef]
- Fiscarelli, E.V.; Rossitto, M.; Rosati, P.; Essa, N.; Crocetta, V.; Di Giulio, A.; Lupetti, V.; Di Bonaventura, G.; Pompilio, A. In vitro newly isolated environmental phage activity against biofilms preformed by Pseudomonas aeruginosa from patients with cystic fibrosis. Microorganisms 2021, 9, 478. [Google Scholar] [CrossRef]
- Chen, F.; Cheng, X.; Li, J.; Yuan, X.; Huang, X.; Lian, M.; Li, W.; Huang, T.; Xie, Y.; Liu, J.; et al. Novel Lytic Phages Protect Cells and Mice against Pseudomonas aeruginosa Infection. J. Virol. 2021, 95, e01832-20. [Google Scholar] [CrossRef] [PubMed]
- Camens, S.; Liu, S.; Hon, K.; Bouras, G.S.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Preclinical development of a bacteriophage cocktail for treating multidrug resistant Pseudomonas aeruginosa infections. Microorganisms 2021, 9, 2001. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.; Morales, S.; Alton, E.W.F.W.; Davies, J.C. Lytic bacteriophage is a promising adjunct to common antibiotics across cystic fibrosis clinical strains and culture models of Pseudomonas aeruginosa infection. Antibiotics 2023, 12, 593. [Google Scholar] [CrossRef]
- Qin, S.; Liu, Y.; Chen, Y.; Hu, J.; Xiao, W.; Tang, X.; Li, G.; Lin, P.; Pu, Q.; Wu, Q.; et al. Engineered bacteriophages containing anti-CRISPR suppress infection of antibiotic-resistant P. aeruginosa. Microbiol. Spectr. 2022, 10, e0160222. [Google Scholar] [CrossRef]
- Cafora, M.; Poerio, N.; Forti, F.; Loberto, N.; Pin, D.; Bassi, R.; Aureli, M.; Briani, F.; Pistocchi, A.; Fraziano, M. Evaluation of phages and liposomes as combination therapy to counteract Pseudomonas aeruginosa infection in wild-type and CFTR-null models. Front. Microbiol. 2022, 13, 979610. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, H.R.; Evans, D.R.; Finney, A.G.; Westbrook, K.J.; Zamora, P.F.; Hofstaedter, C.E.; Yassin, M.H.; Pradhan, A.; Iovleva, A.; Ernst, R.K.; et al. Genomic characterization of lytic bacteriophages targeting genetically diverse Pseudomonas aeruginosa clinical isolates. iScience 2022, 25, 104372. [Google Scholar] [CrossRef]
- Tamma, P.D.; Souli, M.; Billard, M.; Campbell, J.; Conrad, D.; Ellison, D.W.; Evans, B.; Evans, S.R.; Greenwood-Quaintance, K.E.; Filippov, A.A.; et al. Antbacterial Resistance Leadership Group. Safety and microbiological activity of phage therapy in persons with cystic fibrosis colonized with Pseudomonas aeruginosa: Study protocol for a phase 1b/2, multicenter, randomized, double-blind, placebo-controlled trial. Trials 2022, 23, 1057. [Google Scholar]
- Hahn, A.; Sami, I.; Chaney, H.; Koumbourlis, A.C.; Del Valle Mojica, C.; Cochrane, C.; Chan, B.K.; Koff, J.L. Bacteriophage therapy for pan-drug-resistant Pseudomonas aeruginosa in two persons with cystic fibrosis. J. Investig. Med. High. Impact Case Rep. 2023, 11, 23247096231188243. [Google Scholar] [CrossRef]
- Geyer, J.; Krupa, K.A.; Harris, Z.M.; Sun, Y.; Sharma, L.; Würstle, S.; Hu, B.; Stanley, G.; Rajagopalan, G.; Pellot, E.; et al. A novel zinc [II] porphyrin is synergistic with PEV2 bacteriophage against Pseudomonas aeruginosa infections. Antibiotics 2023, 12, 735. [Google Scholar] [CrossRef]
- Ofek, I.; Cohen, S.; Rahmani, R.; Kabha, K.; Tamarkin, D.; Herzig, Y.; Rubinstein, E. Antibacterial synergism of polymyxin B nonapeptide and hydrophobic antibiotics in experimental Gram-negative infections in mice. Antimicrob. Agents Chemother. 1994, 38, 374–377. [Google Scholar] [CrossRef]
- Domalaon, R.; Berry, L.; Tays, Q.; Zhanel, G.G.; Schweizer, F. Development of dilipid polymyxins: Investigation on the effect of hydrophobicity through its fatty acyl component. Bioorg Chem. 2018, 80, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, D.M.; Ramirez, D.; Arthur, G.; Zhanel, G.; Schweizer, F. Guanidinylated polymyxins as outer membrane permeabilizers capable of potentiating rifampicin, erythromycin, ceftazidime and aztreonam against Gram-negative bacteria. Antibiotics 2022, 11, 1277. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Lu, C.D. Polyamines increase antibiotic susceptibility in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Pieri, C.; Borselli, D.; Di Giorgio, C.; De Méo, M.; Bolla, J.M.; Vidal, N.; Combes, S.; Brunel, J.M. New Ianthelliformisamine derivatives as antibiotic enhancers against resistant Gram-negative bacteria. J. Med. Chem. 2014, 57, 4263–4272. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Brunel, J.M.; Preusse, M.; Mozaheb, N.; Willger, S.D.; Larrouy-Maumus, G.; Baatsen, P.; Häussler, S.; Bolla, J.M.; Van Bambeke, F. The membrane-active polyaminoisoprenyl compound NV716 re-sensitizes Pseudomonas aeruginosa to antibiotics and reduces bacterial virulence. Commun. Biol. 2022, 5, 871. [Google Scholar] [CrossRef]
- Wang, G.; Brunel, J.M.; Bolla, J.M.; Van Bambeke, F. The polyaminoisoprenyl potentiator NV716 revives old disused antibiotics against intracellular forms of infection by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2021, 65, e02028-20. [Google Scholar] [CrossRef]
- Douafer, H.; Andrieu, V.; Wafo, E.; Brunel, J.M. Characterization of a new aerosol antibiotic/adjuvant combination for the treatment of P. aeruginosa lung infections. Int. J. Pharm. 2020, 586, 119548. [Google Scholar] [CrossRef]
- Dhanda, G.; Mukherjee, R.; Basak, D.; Haldar, J. Small-molecular adjuvants with weak membrane perturbation potentiate antibiotics against Gram-negative superbugs. ACS Infect. Dis. 2022, 8, 1086–1097. [Google Scholar] [CrossRef]
- Li, S.A.; Cadelis, M.M.; Sue, K.; Blanchet, M.; Vidal, N.; Brunel, J.M.; Bourguet-Kondracki, M.L.; Copp, B.R. 6-bromoindolglyoxylamido derivatives as antimicrobial agents and antibiotic enhancers. Bioorg. Med. Chem. 2019, 27, 2090–2099. [Google Scholar] [CrossRef]
- Cadelis, M.M.; Pike, E.I.W.; Kang, W.; Wu, Z.; Bourguet-Kondracki, M.L.; Blanchet, M.; Vidal, N.; Brunel, J.M.; Copp, B.R. Exploration of the antibiotic potentiating activity of indolglyoxylpolyamines. Eur. J. Med. Chem. 2019, 183, 111708. [Google Scholar] [CrossRef]
- Cadelis, M.M.; Li, S.A.; Bourguet-Kondracki, M.L.; Blanchet, M.; Douafer, H.; Brunel, J.M.; Copp, B.R. Spermine derivatives of indole-3-carboxylic acid, indole-3-acetic acid and indole-3-acrylic acid as Gram-negative antibiotic adjuvants. ChemMedChem 2021, 16, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Pearce, A.N.; Chen, D.; Edmeades, L.R.; Cadelis, M.M.; Troudi, A.; Brunel, J.M.; Bourguet-Kondracki, M.L.; Copp, B.R. Repurposing primaquine as a polyamine conjugate to become an antibiotic adjuvant. Bioorg. Med. Chem. 2021, 38, 116110. [Google Scholar] [CrossRef] [PubMed]
- Sue, K.; Cadelis, M.M.; Gill, E.S.; Rouvier, F.; Bourguet-Kondracki, M.L.; Brunel, J.M.; Copp, B.R. Indole-3-acetamido-polyamines as antimicrobial agents and antibiotic adjuvants. Biomolecules 2023, 13, 1226. [Google Scholar] [CrossRef] [PubMed]
- Cadelis, M.M.; Liu, T.; Sue, K.; Rouvier, F.; Bourguet-Kondracki, M.L.; Brunel, J.M.; Copp, B.R. Structure-activity relationship studies of indolglyoxyl-polyamine conjugates as antimicrobials and antibiotic potentiators. Pharmaceuticals 2023, 16, 823. [Google Scholar] [CrossRef]
- Bradshaw, J. Cationic antimicrobial peptides: Issues for potential clinical use. BioDrugs 2003, 17, 233–240. [Google Scholar] [CrossRef]
- Domalaon, R.; Sanchak, Y.; Koskei, L.C.; Lyu, Y.; Zhanel, G.G.; Arthur, G.; Schweizer, F. Short proline-rich lipopeptide potentiates minocycline and rifampin against multidrug- and extensively drug-resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e02374-17. [Google Scholar] [CrossRef]
- Domalaon, R.; Brizuela, M.; Eisner, B.; Findlay, B.; Zhanel, G.G.; Schweizer, F. Dilipid ultrashort cationic lipopeptides as adjuvants for chloramphenicol and other conventional antibiotics against Gram-negative bacteria. Amino Acids 2019, 51, 383–393. [Google Scholar] [CrossRef]
- Ramirez, D.; Berry, L.; Domalaon, R.; Brizuela, M.; Schweizer, F. Dilipid ultrashort tetrabasic peptidomimetics potentiate novobiocin and rifampicin against multidrug-resistant Gram-negative bacteria. ACS Infect. Dis. 2020, 6, 1413–1426. [Google Scholar] [CrossRef]
- Xia, Y.; Cebrián, R.; Xu, C.; Jong, A.; Wu, W.; Kuipers, O.P. Elucidating the mechanism by which synthetic helper peptides sensitize Pseudomonas aeruginosa to multiple antibiotics. PLoS Pathog. 2021, 17, e1009909. [Google Scholar] [CrossRef]
- Chan, L.W.; Hern, K.E.; Ngambenjawong, C.; Lee, K.; Kwon, E.J.; Hung, D.T.; Bhatia, S.N. Selective permeabilization of Gram-negative bacterial membranes using multivalent peptide constructs for antibiotic sensitization. ACS Infect. Dis. 2021, 7, 721–732. [Google Scholar] [CrossRef]
- Cezard, A.; Fouquenet, D.; Vasseur, V.; Jeannot, K.; Launaym, F.; Si-Tahar, M.; Hervé, V. Poly-L-Lysine to Fight Antibiotic Resistances of Pseudomonas aeruginosa. Int. J. Mol. Sci. 2023, 24, 2851. [Google Scholar] [CrossRef] [PubMed]
- Bulitta, J.B.; Ly, N.S.; Landersdorfer, C.B.; Wanigaratne, N.A.; Velkov, T.; Yadav, R.; Oliver, A.; Martin, L.; Shin, B.S.; Forrest, A.; et al. Two mechanisms of killing of Pseudomonas aeruginosa by tobramycin assessed at multiple inocula via mechanism-based modeling. Antimicrob. Agents Chemother. 2015, 59, 2315–2327. [Google Scholar] [CrossRef]
- Idowu, T.; Arthur, G.; Zhanel, G.G.; Schweizer, F. Heterodimeric Rifampicin-Tobramycin conjugates break intrinsic resistance of Pseudomonas aeruginosa to doxycycline and chloramphenicol in vitro and in a Galleria mellonella in vivo model. Eur. J. Med. Chem. 2019, 174, 16–32. [Google Scholar] [CrossRef]
- Berry, L.; Domalaon, R.; Brizuela, M.; Zhanel, G.G.; Schweizer, F. Polybasic peptide-levofloxacin conjugates potentiate fluoroquinolones and other classes of antibiotics against multidrug-resistant Gram-negative bacteria. Medchemcomm 2019, 10, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Idowu, T.; Ammeter, D.; Brizuela, M.; Jackson, G.; Alam, S.; Schweizer, F. Overcoming β-Lactam resistance in Pseudomonas aeruginosa using non-canonical tobramycin-based antibiotic adjuvants. Bioorg. Med. Chem. Lett. 2020, 30, 127575. [Google Scholar] [CrossRef]
- Yang, X.; Domalaon, R.; Lyu, Y.; Zhanel, G.G.; Schweizer, F. Tobramycin-linked efflux pump inhibitor conjugates synergize fluoroquinolones, rifampicin and fosfomycin against multidrug-resistant Pseudomonas aeruginosa. J. Clin. Med. 2018, 7, 158. [Google Scholar] [CrossRef] [PubMed]
- Idowu, T.; Ammeter, D.; Rossong, H.; Zhanel, G.G.; Schweizer, F. Homodimeric tobramycin adjuvant repurposes novobiocin as an effective antibacterial agent against Gram-negative bacteria. J. Med. Chem. 2019, 62, 9103–9115. [Google Scholar] [CrossRef]
- Idowu, T.; Ammeter, D.; Arthur, G.; Zhanel, G.G.; Schweizer, F. Potentiation of β-lactam antibiotics and β-lactam/β-lactamase inhibitor combinations against MDR and XDR Pseudomonas aeruginosa using non-ribosomal tobramycin-cyclam conjugates. J. Antimicrob. Chemother. 2019, 74, 2640–2648. [Google Scholar] [CrossRef]
- Dhiman, S.; Ramirez, D.; Arora, R.; Gandhi, K.; Wimalasekara, R.; Arthur, G.; Kumar, A.; Schweizer, F. Trimeric Tobramycin/Nebramine synergizes β-lactam antibiotics against Pseudomonas aeruginosa. ACS Omega 2023, 8, 29359–29373. [Google Scholar] [CrossRef]
- Dhiman, S.; Ramirez, D.; Li, Y.; Kumar, A.; Arthur, G.; Schweizer, F. Chimeric tobramycin-based adjuvant TOB-TOB-CIP potentiates fluoroquinolone and β-lactam antibiotics against multidrug-resistant Pseudomonas aeruginosa. ACS Infect. Dis. 2023, 9, 864–885. [Google Scholar] [CrossRef] [PubMed]
- Scoffone, V.C.; Trespidi, G.; Barbieri, G.; Irudal, S.; Perrin, E.; Buroni, S. Role of RND efflux pumps in drug resistance of cystic fibrosis pathogens. Antibiotics 2021, 10, 863. [Google Scholar] [CrossRef] [PubMed]
- Perrin, E.; Fondi, M.; Papaleo, M.C.; Maida, I.; Buroni, S.; Pasca, M.R.; Riccardi, G.; Fani, R. Exploring the HME and HAE1 efflux systems in the genus Burkholderia. BMC Evol. Biol. 2010, 10, 164. [Google Scholar] [CrossRef] [PubMed]
- Giorgini, G.; Mangiaterra, G.; Cedraro, N.; Laudadio, E.; Sabbatini, G.; Cantarini, M.; Minnelli, C.; Mobbili, G.; Frangipani, E.; Biavasco, F.; et al. Berberine derivatives as Pseudomonas aeruginosa MexXY-OprM inhibitors: Activity and in silico insights. Molecules 2021, 26, 6644. [Google Scholar] [CrossRef] [PubMed]
- Dou, Q.; Zhu, Y.; Li, C.; Bian, Z.; Song, H.; Zhang, R.; Wang, Y.; Zhang, X.; Wang, Y. 4F-indole enhances the susceptibility of Pseudomonas aeruginosa to aminoglycoside antibiotics. Microbiol. Spectr. 2023, 11, e0451922. [Google Scholar] [CrossRef] [PubMed]
- Schnaars, C.; Kildahl-Andersen, G.; Prandina, A.; Popal, R.; Radix, S.; Le Borgne, M.; Gjøen, T.; Andresen, A.M.S.; Heikal, A.; Økstad, O.A.; et al. Synthesis and preclinical evaluation of TPA-based zinc chelators as metallo-β-lactamase inhibitors. ACS Infect. Dis. 2018, 4, 1407–1422. [Google Scholar] [CrossRef]
- Principe, L.; Vecchio, G.; Sheehan, G.; Kavanagh, K.; Morroni, G.; Viaggi, V.; di Masi, A.; Giacobbe, D.R.; Luzzaro, F.; Luzzati, R.; et al. Zinc chelators as carbapenem adjuvants for metallo-β-lactamase-producing bacteria: In vitro and in vivo evaluation. Microb. Drug Resist. 2020, 26, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Zeiser, E.T.; Becka, S.A.; Wilson, B.M.; Barnes, M.D.; LiPuma, J.J.; Papp-Wallace, K.M. “Switching Partners”: Piperacillin-avibactam is a highly potent combination against multidrug-resistant Burkholderia cepacia complex and Burkholderia gladioli cystic fibrosis isolates. J. Clin. Microbiol. 2019, 57, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Becka, S.A.; Zeiser, E.T.; LiPuma, J.J.; Papp-Wallace, K.M. Activity of imipenem-relebactam against multidrug- and extensively drug-resistant Burkholderia cepacia complex and Burkholderia gladioli. Antimicrob. Agents Chemother. 2021, 65, e0133221. [Google Scholar] [CrossRef]
- Sader, H.S.; Mendes, R.E.; Duncan, L.R.; Carvalhaes, C.G.; Castanheria, M. Antimicrobial activity of cefepime/zidebactam [WCK 5222], a β-lactam/β-lactam enhancer combination, against clinical isolates of Gram-negative bacteria collected worldwide [2018–19]. J. Antimicrob. Chemother 2022, 77, 2642–2649. [Google Scholar] [CrossRef]
- Domalaon, R.; De Silva, P.M.; Kumar, A.; Zhanel, G.G.; Schweizer, F. The anthelmintic drug niclosamide synergizes with colistin and reverses colistin resistance in Gram-negative bacilli. Antimicrob. Agents Chemother. 2019, 63, e02574-18. [Google Scholar] [CrossRef]
- Lu, T.; Zheng, X.; Mao, F.; Cao, Q.; Cao, Q.; Zhu, J.; Li, X.; Lan, L.; Li, B.; Li, J. Novel niclosamide-derived adjuvants elevating the efficacy of polymyxin B against MDR Pseudomonas aeruginosa DK2. Eur. J. Med. Chem. 2022, 236, 114318. [Google Scholar] [CrossRef]
- Silva, E.; Monteiro, R.; Grainha, T.; Alves, D.; Pereira, M.O.; Sousa, A.M. Fostering innovation in the treatment of chronic polymicrobial cystic fibrosis-associated infections exploring aspartic acid and succinic acid as ciprofloxacin adjuvants. Front. Cell Infect. Microbiol. 2020, 10, 441. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.M.; Valvano, M.A. Vitamin E increases antimicrobial sensitivity by inhibiting bacterial lipocalin antibiotic binding. mSphere 2018, 3, e00564-18. [Google Scholar] [CrossRef] [PubMed]
- Aiyer, A.; Manoharan, A.; Paino, D.; Farrell, J.; Whiteley, G.S.; Kriel, F.H.; Glasbey, T.O.; Manos, J.; Das, T. Disruption of biofilms and killing of Burkholderia cenocepacia from cystic fibrosis lung using an antioxidant-antibiotic combination therapy. Int. J. Antimicrob. Agents 2021, 58, 106372. [Google Scholar] [CrossRef]
- Ciacci, N.; Boncompagni, S.; Valzano, F.; Cariani, L.; Aliberti, S.; Blasi, F.; Pollini, S.; Rossolini, G.M.; Pallecchi, L. In vitro synergism of colistin and N-acetylcysteine against Stenotrophomonas maltophilia. Antibiotics 2019, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Aiyer, A.; Das, T.; Whiteley, G.S.; Glasbey, T.; Kriel, F.H.; Farrell, J.; Manos, J. The efficacy of an N-acetylcysteine-antibiotic combination therapy on Achromobacter xylosoxidans in a cystic fibrosis sputum/lung cell model. Biomedicines 2022, 10, 2886. [Google Scholar] [CrossRef]
- Guerini, M.; Condrò, G.; Friuli, V.; Maggi, L.; Perugini, P. N-acetylcysteine [NAC] and its role in clinical practice management of cystic fibrosis [CF]: A review. Pharmaceuticals 2022, 15, 217. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Hancock, R.E.; Nijnik, A.; Philpott, D.J. Modulating immunity as a therapy for bacterial infections. Nat. Rev. Microbiol. 2012, 10, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Ghimire, J.; Wu, E.; Wimley, W.C. Mechanistic landscape of membrane-permeabilizing peptides. Chem. Rev. 2019, 119, 6040–6085. [Google Scholar] [CrossRef]
- Rončević, T.; Puizina, J.; Tossi, A. Antimicrobial peptides as anti-infective agents in pre-post-antibiotic era? Int. J. Mol. Sci. 2019, 20, 5713. [Google Scholar] [CrossRef] [PubMed]
- Le, C.F.; Fang, C.M.; Sekaran, S.D. Intracellular targeting mechanisms by antimicrobial peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [PubMed]
- Beringer, P.M.; Bensman, T.J.; Ho, H.; Agnello, M.; Denovel, N.; Nguyen, A.; Wong-Beringer, A.; She, R.; Tran, D.Q.; Moskowitz, S.M.; et al. Rhesus θ-defensin-1 [RTD-1] exhibits in vitro and in vivo activity against cystic fibrosis strains of Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2016, 71, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Tai, K.P.; Kamdar, K.; Yamaki, J.; Le, V.V.; Tran, D.; Tran, P.; Selsted, M.E.; Ouellette, A.J.; Wong-Beringer, A. Microbicidal effects of α- and θ-defensins against antibiotic-resistant Staphylococcus aureus and Pseudomonas aeruginosa. Innate Immun. 2015, 21, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Dughbaj, M.A.; Jayne, J.G.; Park, A.Y.J.; Bensman, T.J.; Algorri, M.; Ouellette, A.J.; Selsted, M.E.; Beringer, P.M. Anti-inflammatory effects of RTD-1 in a murine model of chronic Pseudomonas aeruginosa lung infection: Inhibition of NF-κB, inflammasome gene expression, and pro-IL-1β biosynthesis. Antibiotics 2021, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Bensman, T.J.; Jayne, J.G.; Sun, M.; Kimura, E.; Meinert, J.; Wang, J.C.; Schaal, J.B.; Tran, D.; Rao, A.P.; Akbari, O.; et al. Efficacy of Rhesus theta-defensin-1 in experimental models of Pseudomonas aeruginosa lung infection and inflammation. Antimicrob. Agents Chemother. 2017, 61, e00154-17. [Google Scholar] [CrossRef] [PubMed]
- Jahnsen, R.D.; Haney, E.F.; Franzyk, H.; Hancock, R.E. Characterization of a proteolytically stable multifunctional host defense peptidomimetic. Chem. Biol. 2013, 20, 1286–1295. [Google Scholar] [CrossRef]
- Saiman, L.; Tabibi, S.; Starner, T.D.; San Gabriel, P.; Winokur, P.L.; Jia, H.P.; McCray, P.B., Jr.; Tack, B.F. Cathelicidin peptides inhibit multiply antibiotic-resistant pathogens from patients with cystic fibrosis. Antimicrob. Agents Chemother. 2001, 45, 2838–2844. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Scocchi, M.; Pomponio, S.; Guida, F.; Di Primio, A.; Fiscarelli, E.; Gennaro, R.; Di Bonaventura, G. Antibacterial and anti-biofilm effects of cathelicidin peptides against pathogens isolated from cystic fibrosis patients. Peptides 2011, 32, 1807–1814. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; Scocchi, M.; Pomponio, S.; Di Vincenzo, V.; Mardirossian, M.; Gherardi, G.; Fiscarelli, E.; Dicuonzo, G.; Gennaro, R.; et al. Potential novel therapeutic strategies in cystic fibrosis: Antimicrobial and anti-biofilm activity of natural and designed α-helical peptides against Staphylococcus aureus, Pseudomonas aeruginosa, and Stenotrophomonas maltophilia. BMC Microbiol. 2012, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Mardirossian, M.; Pompilio, A.; Degasperi, M.; Runti, G.; Pacor, S.; Di Bonaventura, G.; Scocchi, M. D-BMAP18 antimicrobial peptide is active in vitro, resists to pulmonary proteases but loses its activity in a murine model of Pseudomonas aeruginosa lung infection. Front. Chem. 2017, 5, 40. [Google Scholar] [CrossRef]
- Chen, C.; Mangoni, M.L.; Di, Y.P. In vivo therapeutic efficacy of frog skin-derived peptides against Pseudomonas aeruginosa-induced pulmonary infection. Sci. Rep. 2017, 7, 8548. [Google Scholar] [CrossRef]
- Loffredo, M.R.; Ghosh, A.; Harmouche, N.; Casciaro, B.; Luca, V.; Bortolotti, A.; Cappiello, F.; Stella, L.; Bhunia, A.; Bechinger, B.; et al. Membrane perturbing activities and structural properties of the frog-skin derived peptide Esculentin-1a [1-21]NH2 and its diastereomer Esc [1-21]-1c: Correlation with their antipseudomonal and cytotoxic activity. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2327–2339. [Google Scholar] [CrossRef] [PubMed]
- Degasperi, M.; Agostinis, C.; Mardirossian, M.; Maschio, M.; Taddio, A.; Bulla, R.; Scocchi, M. The anti-pseudomonal peptide D-BMAP18 is active in cystic fibrosis sputum and displays anti-inflammatory in vitro activity. Microorganisms 2020, 8, 1407. [Google Scholar] [CrossRef] [PubMed]
- Jahan, I.; Kumar, S.D.; Shin, S.Y.; Lee, C.W.; Shin, S.H.; Yang, S. Multifunctional properties of BMAP-18 and its aliphatic analog against drug-resistant bacteria. Pharmaceuticals 2023, 16, 1356. [Google Scholar] [CrossRef]
- Forde, E.; Humphreys, H.; Greene, C.M.; Fitzgerald-Hughes, D.; Devocelle, M. Potential of host defense peptide prodrugs as neutrophil elastase-dependent anti-infective agents for cystic fibrosis. Antimicrob. Agents Chemother. 2014, 58, 978–985. [Google Scholar] [CrossRef]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Zapotoczna, M.; Forde, É.; Hogan, S.; Humphreys, H.; O’Gara, J.P.; Fitzgerald-Hughes, D.; Devocelle, M.; O’Neill, E. Eradication of Staphylococcus aureus biofilm infections using synthetic antimicrobial peptides. J. Infect. Dis. 2017, 215, 975–983. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kang, J.H.; Lee, M.K.; Kim, S.Y.; Kim, Y.; Hahm, K.S. Cecropin A—Magainin 2 hybrid peptides having potent antimicrobial activity with low hemolytic effect. Biochem. Mol. Biol. Int. 1998, 44, 1119–1126. [Google Scholar] [CrossRef]
- Cantisani, M.; Leone, M.; Mignogna, E.; Kampanaraki, K.; Falanga, A.; Morelli, G.; Galdiero, M.; Galdiero, S. Structure-activity relations of myxinidin, an antibacterial peptide derived from the epidermal mucus of hagfish. Antimicrob. Agents Chemother. 2013, 57, 5665–5673. [Google Scholar] [CrossRef] [PubMed]
- Forde, É.; Schütte, A.; Reeves, E.; Greene, C.; Humphreys, H.; Mall, M.; Fitzgerald-Hughes, D.; Devocelle, M. Differential in vitro and in vivo toxicities of antimicrobial peptide prodrugs for potential use in cystic fibrosis. Antimicrob. Agents Chemother. 2016, 60, 2813–2821. [Google Scholar] [CrossRef] [PubMed]
- Degasperi, M.; Sgarra, R.; Mardirossian, M.; Pacor, S.; Maschio, M.; Scocchi, M. Elastase-activated antimicrobial peptide for a safer pulmonary treatment of cystic fibrosis infections. Antibiotics 2022, 11, 319. [Google Scholar] [CrossRef]
- Tam, J.P.; Lu, Y.A.; Yang, J.L. Antimicrobial dendrimeric peptides. Eur. J. Biochem. 2002, 269, 923–932. [Google Scholar] [CrossRef]
- Bracci, L.; Falciani, C.; Lelli, B.; Lozzi, L.; Runci, Y.; Pini, A.; De Montis, M.G.; Tagliamonte, A.; Neri, P. Synthetic peptides in the form of dendrimers become resistant to protease activity. J. Biol. Chem. 2003, 278, 46590–46595. [Google Scholar] [CrossRef]
- Pini, A.; Falciani, C.; Mantengoli, E.; Bindi, S.; Brunetti, J.; Iozzi, S.; Rossolini, G.M.; Bracci, L. A novel tetrabranched antimicrobial peptide that neutralizes bacterial lipopolysaccharide and prevents septic shock in vivo. FASEB J. 2010, 24, 1015–1022. [Google Scholar] [CrossRef]
- Falciani, C.; Zevolini, F.; Brunetti, J.; Riolo, G.; Gracia, R.; Marradi, M.; Loinaz, I.; Ziemann, C.; Cossío, U.; Llop, J.; et al. Antimicrobial Peptide-Loaded Nanoparticles as Inhalation Therapy for Pseudomonas aeruginosa Infections. Int. J. Nanomed. 2020, 15, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Quercini, L.; Brunetti, J.; Riolo, G.; Bindi, S.; Scali, S.; Lampronti, I.; D’Aversa, E.; Wronski, S.; Pollini, S.; Gentile, M.; et al. An antimicrobial molecule mitigates signs of sepsis in vivo and eradicates infections from lung tissue. FASEB J. 2020, 34, 192–207. [Google Scholar] [CrossRef]
- Han, X.; Liu, Y.; Ma, Y.; Zhang, M.; He, Z.; Siriwardena, T.N.; Xu, H.; Bai, Y.; Zhang, X.; Reymond, J.L.; et al. Peptide dendrimers G3KL and TNS18 inhibit Pseudomonas aeruginosa biofilms. Appl. Microbiol. Biotechnol. 2019, 103, 5821–5830. [Google Scholar] [CrossRef]
- Pompilio, A.; Geminiani, C.; Mantini, P.; Siriwardena, T.N.; Di Bonaventura, I.; Reymond, J.L.; Di Bonaventura, G. Peptide dendrimers as “lead compounds” for the treatment of chronic lung infections by Pseudomonas aeruginosa in cystic fibrosis patients: In vitro and in vivo studies. Infect. Drug Resist. 2018, 11, 1767–1782. [Google Scholar] [CrossRef]
- Deslouches, B.; Islam, K.; Craigo, J.K.; Paranjape, S.M.; Montelaro, R.C.; Mietzner, T.A. Activity of the de novo engineered antimicrobial peptide WLBU2 against Pseudomonas aeruginosa in human serum and whole blood: Implications for systemic applications. Antimicrob. Agents Chemother. 2005, 49, 3208–3216. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Deslouches, B.; Montelaro, R.C.; Di, Y.P. Enhanced efficacy of the engineered antimicrobial peptide WLBU2 via direct airway delivery in a murine model of Pseudomonas aeruginosa pneumonia. Clin. Microbiol. Infect. 2018, 24, 547.e1–547.e8. [Google Scholar] [CrossRef]
- Di, Y.P.; Lin, Q.; Chen, C.; Montelaro, R.C.; Doi, Y.; Deslouches, B. Enhanced therapeutic index of an antimicrobial peptide in mice by increasing safety and activity against multidrug-resistant bacteria. Sci. Adv. 2020, 6, eaay6817. [Google Scholar] [CrossRef] [PubMed]
- Sader, H.S.; Flamm, R.K.; Dale, G.E.; Rhomberg, P.R.; Castanheira, M. Murepavadin activity tested against contemporary [2016-17] clinical isolates of XDR Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2018, 73, 2400–2404. [Google Scholar] [CrossRef] [PubMed]
- Ekkelenkamp, M.B.; Cantón, R.; Díez-Aguilar, M.; Tunney, M.M.; Gilpin, D.F.; Bernardini, F.; Dale, G.E.; Elborn, J.S.; Bayjanov, J.R.; Fluit, A. Susceptibility of Pseudomonas aeruginosa recovered from cystic fibrosis patients to murepavadin and 13 comparator antibiotics. Antimicrob. Agents Chemother. 2020, 64, e01541-19. [Google Scholar] [CrossRef]
- Díez-Aguilar, M.; Ekkelenkamp, M.; Morosini, M.I.; Huertas, N.; Del Campo, R.; Zamora, J.; Fluit, A.C.; Tunney, M.M.; Obrecht, D.; Bernardini, F.; et al. Anti-biofilm activity of murepavadin against cystic fibrosis Pseudomonas aeruginosa isolates. J. Antimicrob. Chemother. 2021, 76, 2578–2585. [Google Scholar] [CrossRef]
- Mitragotri, S.; Stayton, P. Organic nanoparticles for drug delivery and imaging. MRS Bull. 2014, 39, 219–223. [Google Scholar] [CrossRef]
- Chen, M.; Shou, Z.; Jin, X.; Chen, Y. Emerging strategies in nanotechnology to treat respiratory tract infections: Realizing current trends for future clinical perspectives. Drug Deliv. 2022, 29, 2442–2458. [Google Scholar] [CrossRef]
- Wang, S.; Yu, S.; Lin, Y.; Zou, P.; Chai, G.; Yu, H.H.; Wickremasinghe, H.; Shetty, N.; Ling, J.; Li, J.; et al. Co-delivery of ciprofloxacin and colistin in liposomal formulations with enhanced in vitro antimicrobial activities against multidrug resistant Pseudomonas aeruginosa. Pharm. Res. 2018, 35, 187. [Google Scholar] [CrossRef]
- Pinto, R.M.; Monteiro, C.; Costa Lima, S.A.; Casal, S.; Van Dijck, P.; Martins, M.C.L.; Nunes, C.; Reis, S. N-Acetyl-l-cysteine-loaded nanosystems as a promising therapeutic approach toward the eradication of Pseudomonas aeruginosa biofilms. ACS Appl. Mater. Interfaces 2021, 13, 42329–42343. [Google Scholar] [CrossRef]
- Messiaen, A.S.; Forier, K.; Nelis, H.; Braeckmans, K.; Coenye, T. Transport of nanoparticles and tobramycin-loaded liposomes in Burkholderia cepacia complex biofilms. PLoS ONE 2013, 8, e79220. [Google Scholar] [CrossRef] [PubMed]
- Morais, D.; Tanoeiro, L.; Marques, A.T.; Gonçalves, T.; Duarte, A.; Matos, A.P.A.; Vital, J.S.; Cruz, M.E.M.; Carvalheiro, M.C.M.; Anes, E.; et al. Liposomal delivery of newly identified prophage lysins in a Pseudomonas aeruginosa model. Int. J. Mol. Sci. 2022, 23, 10143. [Google Scholar] [CrossRef] [PubMed]
- Ghasemiyeh, P.; Mohammadi-Samani, S. Solid lipid nanoparticles and nanostructured lipid carriers as novel drug delivery systems: Applications, advantages and disadvantages. Res. Pharm. Sci. 2018, 13, 288–303. [Google Scholar]
- Pastor, M.; Moreno-Sastre, M.; Esquisabel, A.; Sans, E.; Viñas, M.; Bachiller, D.; Asensio, V.J.; Pozo, A.D.; Gainza, E.; Pedraz, J.L. Sodium colistimethate loaded lipid nanocarriers for the treatment of Pseudomonas aeruginosa infections associated with cystic fibrosis. Int. J. Pharm. 2014, 477, 485–494. [Google Scholar] [CrossRef]
- Vairo, C.; Basas, J.; Pastor, M.; Palau, M.; Gomis, X.; Almirante, B.; Gainza, E.; Hernandez, R.M.; Igartua, M.; Gavaldà, J.; et al. In vitro and in vivo antimicrobial activity of sodium colistimethate and amikacin-loaded nanostructured lipid carriers [NLC]. Nanomedicine 2020, 29, 102259. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Hinojosa, E.I.; Salazar-Sesatty, H.A.; Alvizo-Baez, C.A.; Terrazas-Armendariz, L.D.; Luna-Cruz, I.E.; Alcocer-González, J.M.; Villarreal-Treviño, L.; Flores-Treviño, S. Antibiofilm and antimicrobial activity of curcumin-chitosan nanocomplexes and trimethoprim-sulfamethoxazole on Achromobacter, Burkholderia, and Stenotrophomonas isolates. Expert. Rev. Anti Infect. Ther. 2023, 21, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.K.; Tripathi, M.; Pandey, N.; Agrawal, A.K.; Gade, S.; Anjum, M.M.; Tilak, R.; Singh, S. Alginate lyase immobilized chitosan nanoparticles of ciprofloxacin for the improved antimicrobial activity against the biofilm associated mucoid P. aeruginosa infection in cystic fibrosis. Int. J. Pharm. 2019, 563, 30–42. [Google Scholar] [CrossRef]
- Cresti, L.; Conte, G.; Cappello, G.; Brunetti, J.; Falciani, C.; Bracci, L.; Quaglia, F.; Ungaro, F.; d’Angelo, I.; Pini, A. Inhalable polymeric nanoparticles for pulmonary delivery of antimicrobial peptide SET-M33: Antibacterial activity and toxicity in vitro and in vivo. Pharmaceutics 2023, 15, 3. [Google Scholar] [CrossRef]
- Thorn, C.R.; Carvalho-Wodarz, C.S.; Horstmann, J.C.; Lehr, C.M.; Prestidge, C.A.; Thomas, N. Tobramycin liquid crystal nanoparticles eradicate cystic fibrosis-related Pseudomonas aeruginosa biofilms. Small 2021, 17, e2100531. [Google Scholar] [CrossRef]
- Yu, S.; Pu, X.; Ahmed, M.U.; Yu, H.H.; Mutukuri, T.T.; Li, J.; Zhou, Q.T. Spray-freeze-dried inhalable composite microparticles containing nanoparticles of combinational drugs for potential treatment of lung infections caused by Pseudomonas aeruginosa. Int. J. Pharm. 2021, 610, 121160. [Google Scholar] [CrossRef]
- Cho, D.Y.; Lim, D.J.; Mackey, C.; Skinner, D.; Zhang, S.; McCormick, J.; Woodworth, B.A. Ivacaftor, a cystic fibrosis transmembrane conductance regulator potentiator, enhances ciprofloxacin activity against Pseudomonas aeruginosa. Am. J. Rhinol. Allergy 2019, 33, 129–136. [Google Scholar] [CrossRef]
- Costabile, G.; Provenzano, R.; Azzalin, A.; Scoffone, V.C.; Chiarelli, L.R.; Rondelli, V.; Grillo, I.; Zinn, T.; Lepioshkin, A.; Savina, S.; et al. PEGylated mucus-penetrating nanocrystals for lung delivery of a new FtsZ inhibitor against Burkholderia cenocepacia infection. Nanomedicine 2020, 23, 102–113. [Google Scholar] [CrossRef]
- Soni, K.S.; Desale, S.S.; Bronich, T.K. Nanogels: An overview of properties, biomedical applications and obstacles to clinical translation. J. Control Release 2016, 240, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Shan, C.L.; Zhou, Q.; Fang, Y.; Wang, Y.L.; Xu, F.; Han, L.R.; Ibrahim, M.; Guo, L.B.; Xie, G.L.; et al. Synthesis, characterization, and antibacterial activity of cross-linked chitosan-glutaraldehyde. Mar. Drugs 2013, 11, 1534–1552. [Google Scholar] [CrossRef] [PubMed]
- Balderrama-González, A.S.; Piñón-Castillo, H.A.; Ramírez-Valdespino, C.A.; Landeros-Martínez, L.L.; Orrantia-Borunda, E.; Esparza-Ponce, H.E. Antimicrobial resistance and inorganic nanoparticles. Int. J. Mol. Sci. 2021, 22, 12890. [Google Scholar] [CrossRef] [PubMed]
- Campo-Beleño, C.; Villamizar-Gallardo, R.A.; López-Jácome, L.E.; González, E.E.; Muñoz-Carranza, S.; Franco, B.; Morales-Espinosa, R.; Coria-Jimenez, R.; Franco-Cendejas, R.; Hernández-Durán, M.; et al. Biologically synthesized silver nanoparticles as potent antibacterial effective against multidrug-resistant Pseudomonas aeruginosa. Lett. Appl. Microbiol. 2022, 75, 680–688. [Google Scholar] [CrossRef]
- Pompilio, A.; Geminiani, C.; Bosco, D.; Rana, R.; Aceto, A.; Bucciarelli, T.; Scotti, L.; Di Bonaventura, G. electrochemically synthesized silver nanoparticles are active against planktonic and biofilm cells of Pseudomonas aeruginosa and other cystic fibrosis-associated bacterial pathogens. Front. Microbiol. 2018, 9, 1349. [Google Scholar] [CrossRef]
- Al-Momani, H.; Almasri, M.; Al Balawi, D.; Hamed, S.; Albiss, B.A.; Aldabaibeh, N.; Ibrahim, L.; Albalawi, H.; Al Haj Mahmoud, S.; Khasawneh, A.I.; et al. The efficacy of biosynthesized silver nanoparticles against Pseudomonas aeruginosa isolates from cystic fibrosis patients. Sci. Rep. 2023, 13, 8876. [Google Scholar] [CrossRef]
- Huang, Y.S.; Wang, J.T.; Tai, H.M.; Chang, P.C.; Huang, H.C.; Yang, P.C. Metal nanoparticles and nanoparticle composites are effective against Haemophilus influenzae, Streptococcus pneumoniae, and multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2022, 55, 708–715. [Google Scholar] [CrossRef]
- Rather, M.A.; Mandal, M. Attenuation of biofilm and quorum sensing regulated virulence factors of an opportunistic pathogen Pseudomonas aeruginosa by phytofabricated silver nanoparticles. Microb. Pathog. 2023, 185, 106433. [Google Scholar] [CrossRef]
- Cremonini, E.; Boaretti, M.; Vandecandelaere, I.; Zonaro, E.; Coenye, T.; Lleo, M.M.; Lampis, S.; Vallini, G. Biogenic selenium nanoparticles synthesized by Stenotrophomonas maltophilia SeITE02 loose antibacterial and antibiofilm efficacy as a result of the progressive alteration of their organic coating layer. Microb. Biotechnol. 2018, 11, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Lee, J.W.; Manivasagan, P.; Pham, D.T.N.; Oh, J.; Kim, Y.M. Synthesis and characterization of chitosan oligosaccharide-capped gold nanoparticles as an effective antibiofilm drug against the Pseudomonas aeruginosa PAO1. Microb. Pathog. 2019, 135, 103623. [Google Scholar] [CrossRef] [PubMed]
- Piktel, E.; Wnorowska, U.; Depciuch, J.; Łysik, D.; Cieśluk, M.; Fiedoruk, K.; Mystkowska, J.; Parlińska-Wojtan, M.; Janmey, P.A.; Bucki, R. N-Acetyl-cysteine increases activity of peanut-shaped gold nanoparticles against biofilms formed by clinical strains of Pseudomonas aeruginosa isolated from sputum of cystic fibrosis patients. Infect. Drug Resist. 2022, 15, 851–871. [Google Scholar] [CrossRef]
- Zahmatkesh, H.; Mirpour, M.; Zamani, H.; Rasti, B.; Rahmani, F.A.; Padasht, N. Effect of samarium oxide nanoparticles on virulence factors and motility of multi-drug resistant Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2022, 38, 209. [Google Scholar] [CrossRef] [PubMed]
- Iafisco, M.; Carella, F.; Degli Esposti, L.; Adamiano, A.; Catalucci, D.; Modica, J.; Bragonzi, A.; Vitali, A.; Torelli, R.; Sanguinetti, M.; et al. Biocompatible antimicrobial colistin loaded calcium phosphate nanoparticles for the counteraction of biofilm formation in cystic fibrosis related infections. J. Inorg. Biochem. 2022, 230, 111751. [Google Scholar] [CrossRef] [PubMed]
- Elborn, J.S.; Blasi, F.; Burgel, P.R.; Peckham, D. Role of inhaled antibiotics in the era of highly effective CFTR modulators. Eur. Respir. Rev. 2023, 32, 220154. [Google Scholar] [CrossRef] [PubMed]
- Fainardi, V.; Neglia, C.; Muscarà, M.; Spaggiari, C.; Tornesello, M.; Grandinetti, R.; Argentiero, A.; Calderaro, A.; Esposito, S.; Pisi, G. Multidrug-resistant bacteria in children and adolescents with cystic fibrosis. Children 2022, 9, 1330. [Google Scholar] [CrossRef]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage therap. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Podlacha, M.; Grabowski, Ł.; Kosznik-Kawśnicka, K.; Zdrojewska, K.; Stasiłojć, M.; Węgrzyn, G.; Węgrzyn, A. Interactions of bacteriophages with animal and human organisms—Safety issues in the light of phage therapy, Int. J. Mol. Sci. 2021, 22, 8937. [Google Scholar] [CrossRef]
- Liu, D.; Van Belleghem, J.D.; de Vries, C.R.; Burgener, E.; Chen, Q.; Manasherob, R.; Aronson, J.R.; Amanatullah, D.F.; Tamma, P.D.; Suh, G.A. The safety and toxicity of phage therapy: A review of animal and clinical studies. Viruses 2021, 13, 1268. [Google Scholar] [CrossRef]
- Bollenbach, T. Antimicrobial interactions: Mechanisms and implications for drug discovery and resistance evolution. Curr. Opin. Microbiol. 2015, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Predicting drug resistance evolution: Insights from antimicrobial peptides and antibiotics. Proc. Biol. Sci. 2018, 285, 20172687. [Google Scholar] [CrossRef] [PubMed]
- Geitani, R.; Ayoub Moubareck, C.; Touqui, L.; Karam Sarkis, D. Cationic antimicrobial peptides: Alternatives and/or adjuvants to antibiotics active against methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa. BMC Microbiol. 2019, 19, 54. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Son, H.; Park, S.C.; Lee, J.K.; Jang, M.K.; Lee, J.R. Anti-biofilm effects of rationally designed peptides against planktonic cells and pre-formed biofilm of Pseudomonas aeruginosa. Antibiotics 2023, 12, 349. [Google Scholar] [CrossRef]
- Bugli, F.; Martini, C.; Di Vito, M.; Cacaci, M.; Catalucci, D.; Gori, A.; Iafisco, M.; Sanguinetti, M.; Vitali, A. Antimicrobial peptides for tackling cystic fibrosis related bacterial infections: A review. Microbiol. Res. 2022, 263, 127152. [Google Scholar] [CrossRef]
- Loutet, S.A.; Valvano, M.A. Extreme antimicrobial peptide and polymyxin B resistance in the genus Burkholderia. Front. Microbiol. 2011, 2, 159. [Google Scholar] [CrossRef] [PubMed]
- Hetta, H.F.; Ramadan, Y.N.; Al-Harbi, A.I.A.; Ahmed, E.; Battah, B.M.; Abd Ellah, N.H.; Zanetti, S.; Donadu, M.G. Nanotechnology as a promising approach to combat multidrug resistant bacteria: A comprehensive review and future perspectives. Biomedicines 2023, 11, 413. [Google Scholar] [CrossRef]
- de la Harpe, K.M.; Kondiah, P.P.D.; Choonara, Y.E.; Marimuthu, T.; du Toit, L.C.; Pillay, V. The hemocompatibility of nanoparticles: A review of cell-nanoparticle interactions and hemostasis. Cells 2019, 8, 1209. [Google Scholar] [CrossRef]
- Rudramurthy, G.R.; Swamy, M.K.; Sinniah, U.R.; Ghasemzadeh, A. Nanoparticles: Alternatives against drug-resistant pathogenic microbes. Molecules 2016, 21, 836. [Google Scholar] [CrossRef]
- Sharma, S.; Sudhakara, P.; Singh, J.; Ilyas, R.A.; Asyraf, M.R.M.; Razman, M.R. Critical review of biodegradable and bioactive polymer composites for bone tissue engineering and drug delivery applications. Polymers 2021, 13, 2623. [Google Scholar] [CrossRef]
- Liu, Y.; Hardie, J.; Zhang, X.; Rotello, V.M. Effects of engineered nanoparticles on the innate immune system. Semin. Immunol. 2017, 34, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. The role of reactive oxygen species [ROS] in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]



| Synthetic Molecules | |||
|---|---|---|---|
| Quorum Sensing Inhibitors | Bacteria | Mechanism | Reference |
| AHL analogs | P. aeruginosa | 3O-C12-HSL competition | [7] |
| Halogenated furanone derivatives | P. aeruginosa | LasR and RhlR inhibition | [8] |
| Halogenated furanone derivatives with palladium-catalyzed coupling reactions | P. aeruginosa | LasR and RhlR inhibition | [9] |
| PqsR modulators | P. aeruginosa | PqsR inhibition | [10] |
| 3-hydroxypyridin-4[1H]-one derivatives | P. aeruginosa | pqs inhibition | [11] |
| Small QS inhibitor, squalene derived nanoparticles formulation | P. aeruginosa | unknown | [12] |
| Nitrofurazone and Erythromycin estolate | P. aeruginosa | unknown | [13] |
| Niclosamide | P. aeruginosa | 3OC12-HSL signaling process | [15] |
| Clofoctol | P. aeruginosa | PqsR competitive inhibitor | [15] |
| Tyramine | B. cenocepacia | CepI/R and CciI/R inhibition | [18] |
| Diketopiperazines | B. cenocepacia | CepI inhibition | [19,20] |
| Sulfonamide-based DSF bioisosteres | S. maltophilia and Bcc | DSF inhibition | [21] |
| Synthetic Molecules | |||
|---|---|---|---|
| Anti-Virulence Molecules | Bacteria | Target | Reference |
| Psammaplin A and bisaprasin | P. aeruginosa | LasB | [34] |
| Benzoxazolone derivatives | P. aeruginosa | Pyocyanin | [35] |
| Gallium nitrate | P. aeruginosa | Siderophores | [36] |
| MEDI3902 | P. aeruginosa | PcrV and Psl | [37] |
| Fluorothiazinon | P. aeruginosa | T3SS | [38] |
| Nonpeptidic inhibitors | H. influenzae | IgA1 proteases | [39] |
| FR90098 | B. cenocepacia | non-mevalonate pathway | [40] |
| Natural Compounds | ||
|---|---|---|
| Quorum Sensing Inhibitors | Bacteria | Reference |
| Coumarin and coumarin derivatives | P. aeruginosa | [41,42] |
| Baicalin | P. aeruginosa, Bcc | [43,44] |
| Oridonin | B. cenocepacia, Burkholderia species | [46] |
| Chromohalobacter sp. D23 | B. cepacia | [47] |
| Celastrol | S. maltophilia | [48] |
| Biofilm inhibitors | Bacteria | Reference |
| Dioon spinulosum extract | P. aeruginosa | [49] |
| Plant-derived triterpenes (analogs of oleanolic acid) | P. aeruginosa, B. cenocepacia | [50] |
| Plant essential oils | P. aeruginosa, H. influenzae | [51,52,53,54] |
| Ceragenins (CSAs) | A. xylosoxidans | [55] |
| Metalloproteases from stony coral | S. maltophilia | [56] |
| Vitamin C | S. maltophilia | [57] |
| Allium stipitatum extract | S. maltophilia | [58] |
| Glycosyl hydrolases PslG from P. fluorescens | Pseudomonas strains | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scoffone, V.C.; Barbieri, G.; Irudal, S.; Trespidi, G.; Buroni, S. New Antimicrobial Strategies to Treat Multi-Drug Resistant Infections Caused by Gram-Negatives in Cystic Fibrosis. Antibiotics 2024, 13, 71. https://doi.org/10.3390/antibiotics13010071
Scoffone VC, Barbieri G, Irudal S, Trespidi G, Buroni S. New Antimicrobial Strategies to Treat Multi-Drug Resistant Infections Caused by Gram-Negatives in Cystic Fibrosis. Antibiotics. 2024; 13(1):71. https://doi.org/10.3390/antibiotics13010071
Chicago/Turabian StyleScoffone, Viola Camilla, Giulia Barbieri, Samuele Irudal, Gabriele Trespidi, and Silvia Buroni. 2024. "New Antimicrobial Strategies to Treat Multi-Drug Resistant Infections Caused by Gram-Negatives in Cystic Fibrosis" Antibiotics 13, no. 1: 71. https://doi.org/10.3390/antibiotics13010071
APA StyleScoffone, V. C., Barbieri, G., Irudal, S., Trespidi, G., & Buroni, S. (2024). New Antimicrobial Strategies to Treat Multi-Drug Resistant Infections Caused by Gram-Negatives in Cystic Fibrosis. Antibiotics, 13(1), 71. https://doi.org/10.3390/antibiotics13010071