

Genome-Based Retrospective Analysis of a Providencia stuartii Outbreak in Rome, Italy: Broad Spectrum IncC Plasmids Spread the NDM Carbapenemase within the Hospital

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. An Outbreak of NDM-Producing Providencia Stuartii
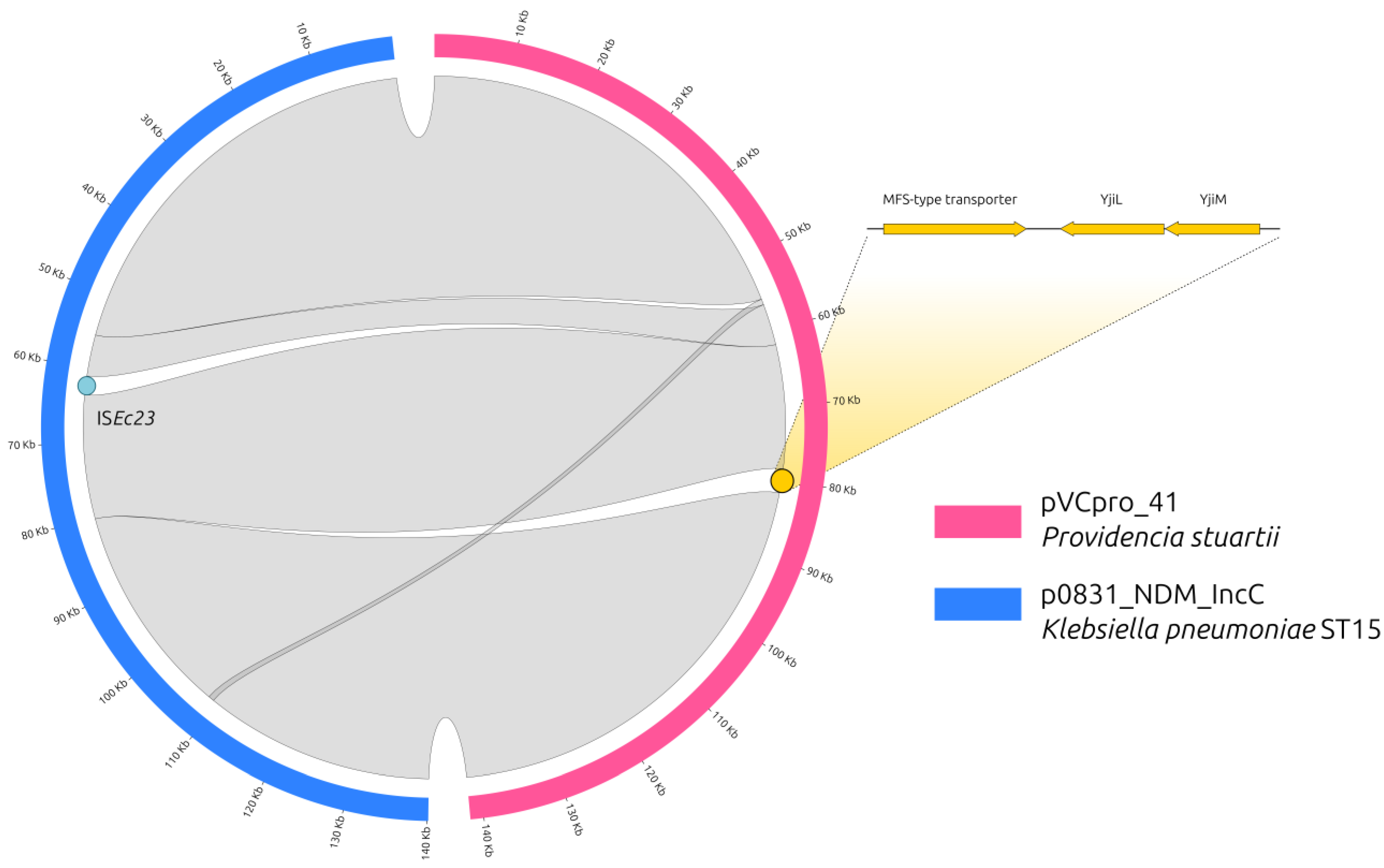
2.2. The NDM-Carrying IncC Plasmid
- p0831_NDM_IncC carried an Insertion Sequence (IS, ISEc23), which was not present in the P. stuartii plasmid;
- pVCpro_41 carried three genes, which were not present in the p0831_NDM_IncC plasmid, encoding for an uncharacterized MFS-type transporter, and two uncharacterized proteins named YjiL and YjiM.
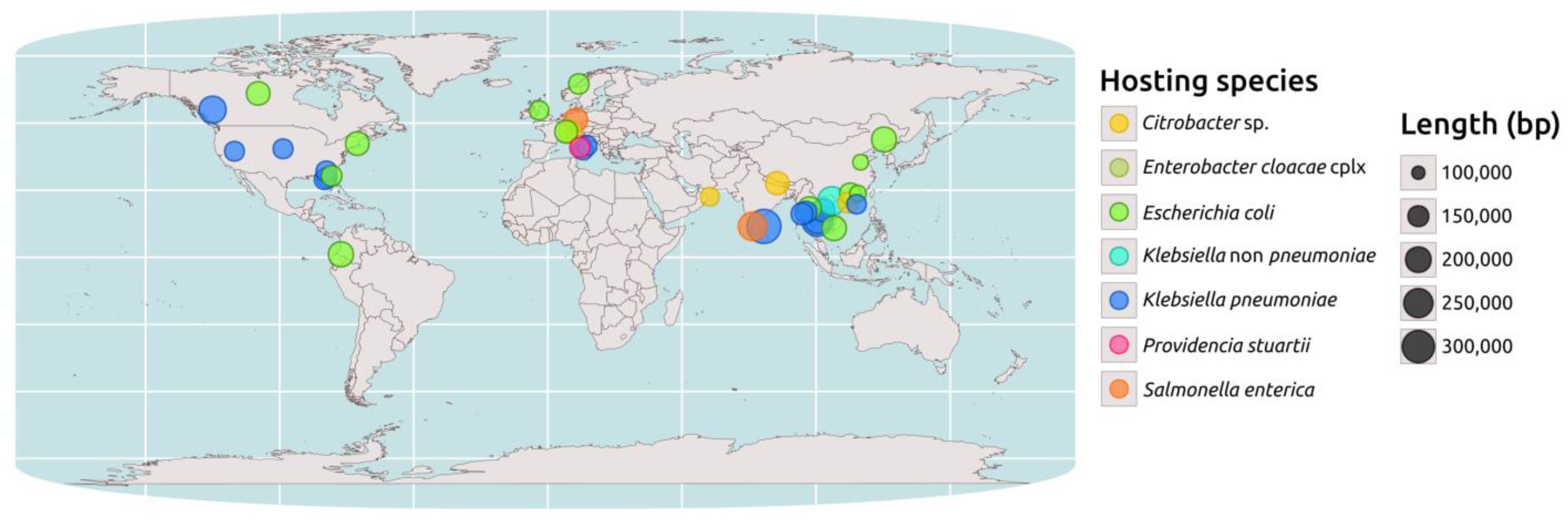
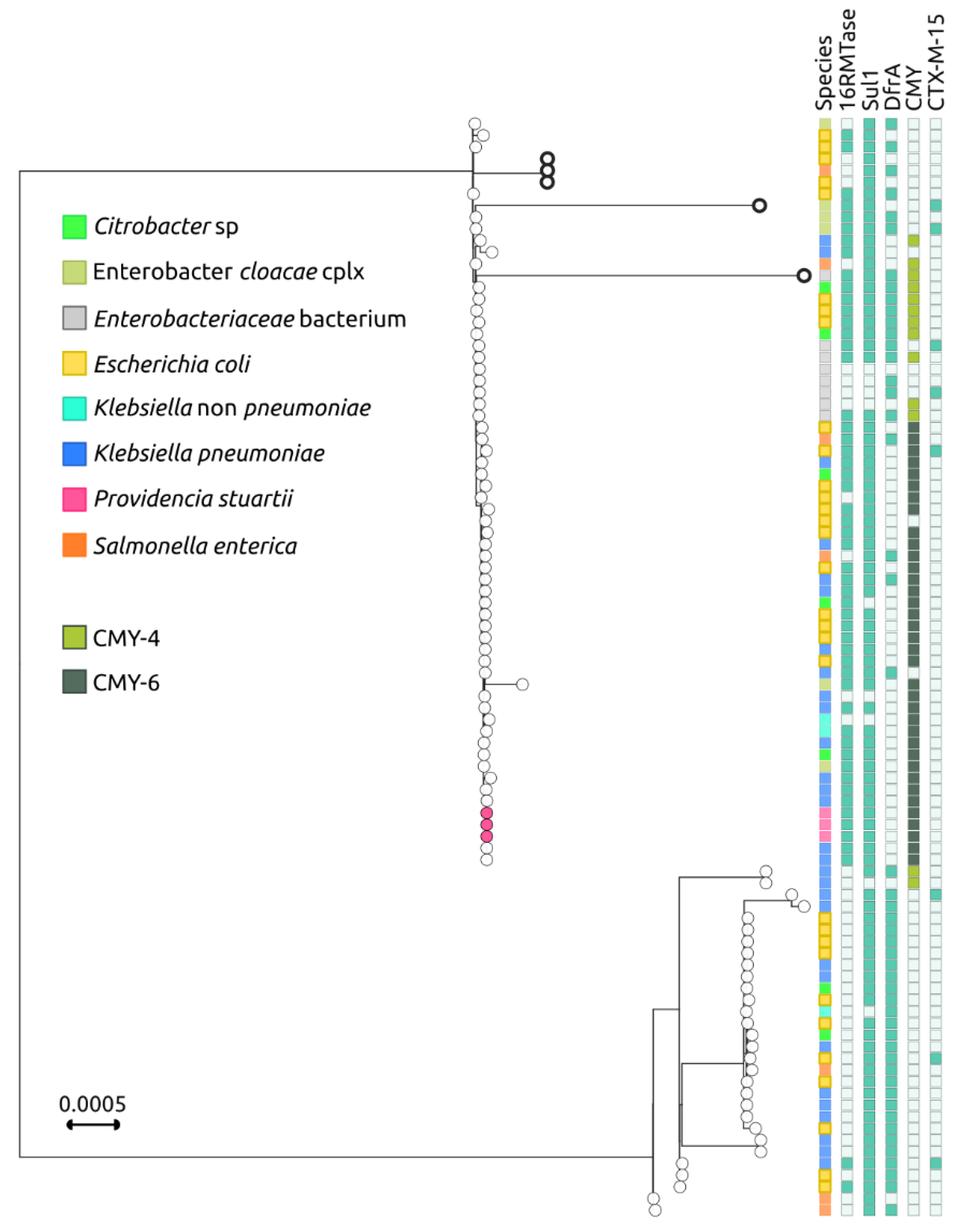
2.3. Genomic Epidemiology of blaNDM-Carrying IncC Plasmids
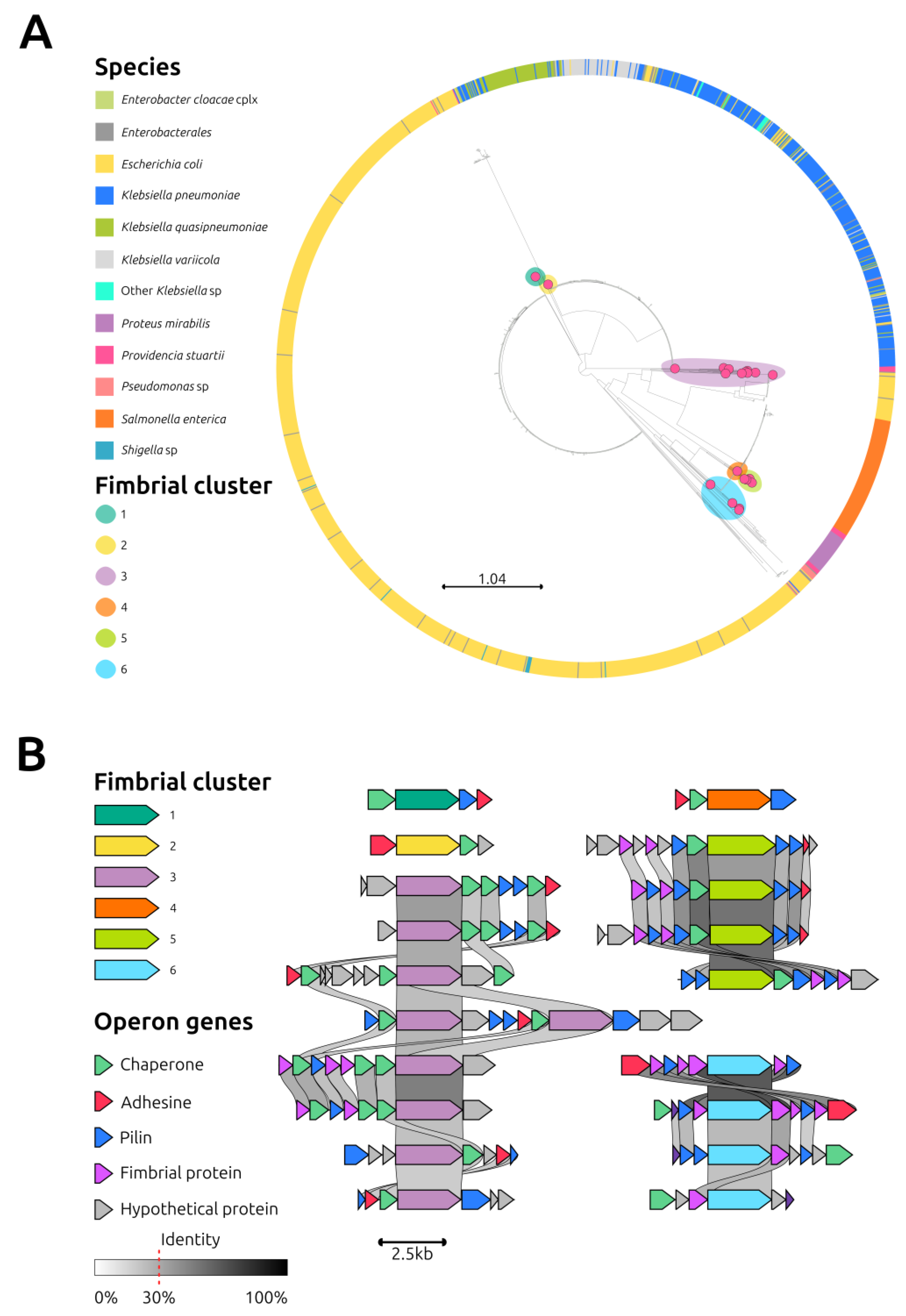
2.4. Anatomy of Three Providencia Stuartii Isolates Carrying the IncC Plasmid
3. Materials and Methods
3.1. Bacterial Isolation and Antimicrobial Susceptibility Testing
3.2. Whole-Genome Sequencing and Assembly
3.3. Genomic and Phylogenetic Analyses
3.4. IncC Plasmids Analysis
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ewing, W.H. The Tribe Proteeae: Its Nomenclature and Taxonomy. Int. Bull. Bacteriol. Nomencl. Taxon. 1962, 12, 93–102. [Google Scholar] [CrossRef]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-Based Phylogeny and Taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales Ord. Nov. Divided into the Families Enterobacteriaceae, Erwiniaceae Fam. Nov., Pectobacteriaceae Fam. Nov., Yersiniaceae Fam. Nov., Hafniaceae Fam. Nov., Morganellaceae Fam. Nov., and Budviciaceae Fam. Nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Molnár, S.; Flonta, M.M.M.; Almaş, A.; Buzea, M.; Licker, M.; Rus, M.; Földes, A.; Székely, E. Dissemination of NDM-1 Carbapenemase-Producer Providencia stuartii Strains in Romanian Hospitals: A Multicentre Study. J. Hosp. Infect. 2019, 103, 165–169. [Google Scholar] [CrossRef]
- Mnif, B.; Ktari, S.; Chaari, A.; Medhioub, F.; Rhimi, F.; Bouaziz, M.; Hammami, A. Nosocomial Dissemination of Providencia stuartii Isolates Carrying BlaOXA-48, BlaPER-1, BlaCMY-4 and QnrA6 in a Tunisian Hospital. J. Antimicrob. Chemother. 2013, 68, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Rakov, C.; Ben Porat, S.; Alkalay-Oren, S.; Yerushalmy, O.; Abdalrhman, M.; Gronovich, N.; Huang, L.; Pride, D.; Coppenhagen-Glazer, S.; Nir-Paz, R.; et al. Targeting Biofilm of MDR Providencia stuartii by Phages Using a Catheter Model. Antibiotics 2021, 10, 375. [Google Scholar] [CrossRef]
- Douka, E.; Perivolioti, E.; Kraniotaki, E.; Fountoulis, K.; Economidou, F.; Tsakris, A.; Skoutelis, A.; Routsi, C. Emergence of a Pandrug-Resistant VIM-1-Producing Providencia stuartii Clonal Strain Causing an Outbreak in a Greek Intensive Care Unit. Int. J. Antimicrob. Agents 2015, 45, 533–536. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q. Mohd.R. Current Update on Intrinsic and Acquired Colistin Resistance Mechanisms in Bacteria. Front. Med. 2021, 8, 677720. [Google Scholar] [CrossRef]
- Yaghoubi, S.; Zekiy, A.O.; Krutova, M.; Gholami, M.; Kouhsari, E.; Sholeh, M.; Ghafouri, Z.; Maleki, F. Tigecycline Antibacterial Activity, Clinical Effectiveness, and Mechanisms and Epidemiology of Resistance: Narrative Review. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1003–1022. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). CLSI Supplement M100: Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Hayakawa, K.; Marchaim, D.; Divine, G.W.; Pogue, J.M.; Kumar, S.; Lephart, P.; Risko, K.; Sobel, J.D.; Kaye, K.S. Growing Prevalence of Providencia stuartii Associated with the Increased Usage of Colistin at a Tertiary Health Care Center. Int. J. Infect. Dis. 2012, 16, e646–e648. [Google Scholar] [CrossRef] [PubMed]
- Moser, A.I.; Keller, P.M.; Campos-Madueno, E.I.; Poirel, L.; Nordmann, P.; Endimiani, A. A Patient With Multiple Carbapenemase Producers Including an Unusual Citrobacter sedlakii Hosting an IncC blaNDM-1- and ArmA-Carrying Plasmid. Pathog. Immun. 2021, 6, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Hoard, A.; Montaña, S.; Moriano, A.; Fernandez, J.S.; Traglia, G.M.; Quiroga, C.; Franchi, A.; Cohen, E.; Corigliano, C.; Almuzara, M.; et al. Genomic Analysis of Two NDM-1 Providencia stuartii Strains Recovered from a Single Patient. Curr. Microbiol. 2020, 77, 4029–4036. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.; Alhababi, R.; Alqudah, N.; Aldyyat, B.; Alharthy, A. First Report of Carbapenem-Resistant Providencia stuartii in Saudi Arabia. New Microbes New Infect. 2018, 26, 107–109. [Google Scholar] [CrossRef]
- Tavares, C.P.; Pereira, P.S.; Marques, E.d.A.; Faria, C.; de Souza, M.d.P.A.H.; de Almeida, R.; Alves, C.d.F.M.; Asensi, M.D.; Carvalho-Assef, A.P.D. Molecular Epidemiology of KPC-2-Producing Enterobacteriaceae (Non-Klebsiella pneumoniae) Isolated from Brazil. Diagn. Microbiol. Infect. Dis. 2015, 82, 326–330. [Google Scholar] [CrossRef]
- Mc Gann, P.; Hang, J.; Clifford, R.J.; Yang, Y.; Kwak, Y.I.; Kuschner, R.A.; Lesho, E.P.; Waterman, P.E. Complete Sequence of a Novel 178-Kilobase Plasmid Carrying BlaNDM-1 in a Providencia stuartii Strain Isolated in Afghanistan. Antimicrob. Agents Chemother. 2012, 56, 1673–1679. [Google Scholar] [CrossRef]
- Manageiro, V.; Sampaio, D.A.; Pereira, P.; Rodrigues, P.; Vieira, L.; Palos, C.; Caniça, M. Draft Genome Sequence of the First NDM-1-Producing Providencia stuartii Strain Isolated in Portugal. Genome Announc. 2015, 3, e01077-15. [Google Scholar] [CrossRef]
- Di Pilato, V.; Henrici De Angelis, L.; Aiezza, N.; Baccani, I.; Niccolai, C.; Parisio, E.M.; Giordano, C.; Camarlinghi, G.; Barnini, S.; Forni, S.; et al. Resistome and Virulome Accretion in an NDM-1-Producing ST147 Sublineage of Klebsiella pneumoniae Associated with an Outbreak in Tuscany, Italy: A Genotypic and Phenotypic Characterisation. Lancet Microbe 2022, 3, e224–e234. [Google Scholar] [CrossRef]
- Martin, M.J.; Corey, B.W.; Sannio, F.; Hall, L.R.; MacDonald, U.; Jones, B.T.; Mills, E.G.; Harless, C.; Stam, J.; Maybank, R.; et al. Anatomy of an Extensively Drug-Resistant Klebsiella pneumoniae Outbreak in Tuscany, Italy. Proc. Natl. Acad. Sci. USA 2021, 118, e2110227118. [Google Scholar] [CrossRef]
- Di Pilato, V.; Errico, G.; Monaco, M.; Giani, T.; Del Grosso, M.; Antonelli, A.; David, S.; Lindh, E.; Camilli, R.; Aanensen, D.M.; et al. The Changing Epidemiology of Carbapenemase-Producing Klebsiella pneumoniae in Italy: Toward Polyclonal Evolution with Emergence of High-Risk Lineages. J. Antimicrob. Chemother. 2021, 76, 355–361. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, S.J.; Harmer, C.J.; Hall, R.M. Evolution and Typing of IncC Plasmids Contributing to Antibiotic Resistance in Gram-Negative Bacteria. Plasmid 2018, 99, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Sacco, F.; Raponi, G.; Oliva, A.; Bibbolino, G.; Mauro, V.; Di Lella, F.M.; Volpicelli, L.; Antonelli, G.; Venditti, M.; Carattoli, A.; et al. An Outbreak Sustained by ST15 Klebsiella pneumoniae Carrying 16S RRNA Methyltransferases and BlaNDM: Evaluation of the Global Dissemination of These Resistance Determinants. Int. J. Antimicrob. Agents 2022, 60, 106615. [Google Scholar] [CrossRef]
- Galata, V.; Fehlmann, T.; Backes, C.; Keller, A. PLSDB: A Resource of Complete Bacterial Plasmids. Nucleic Acids Res. 2019, 47, D195–D202. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A Genomic Surveillance Framework and Genotyping Tool for Klebsiella pneumoniae and Its Related Species Complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Octavia, S.; Chew, K.L.; Chew, K.L.; Lin, R.T.P.; Teo, J.W.P. Multidrug-Resistant Salmonella enterica Serovar London Carrying blaNDM-1 Encoding Plasmid from Singapore. Clin. Microbiol. Infect. 2020, 26, 963–966. [Google Scholar] [CrossRef]
- Weber, R.E.; Pietsch, M.; Frühauf, A.; Pfeifer, Y.; Martin, M.; Luft, D.; Gatermann, S.; Pfennigwerth, N.; Kaase, M.; Werner, G.; et al. IS26-Mediated Transfer of blaNDM–1 as the Main Route of Resistance Transmission During a Polyclonal, Multispecies Outbreak in a German Hospital. Front. Microbiol. 2019, 10, 2817. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Poirel, L.; Bonnin, R.A.; Nordmann, P. Evolution of IncA/C blaCMY-2-Carrying Plasmids by Acquisition of the blaNDM-1 Carbapenemase Gene. Antimicrob. Agents Chemother. 2012, 56, 783–786. [Google Scholar] [CrossRef]
- Schweizer, H.P. Efflux as a Mechanism of Resistance to Antimicrobials in Pseudomonas aeruginosa and Related Bacteria: Unanswered Questions. Genet. Mol. Res. 2003, 2, 48–62. [Google Scholar]
- Kolenda, R.; Ugorski, M.; Grzymajlo, K. Everything You Always Wanted to Know About Salmonella Type 1 Fimbriae, but Were Afraid to Ask. Front. Microbiol. 2019, 10, 1017. [Google Scholar] [CrossRef]
- Carattoli, A.; Arcari, G.; Bibbolino, G.; Sacco, F.; Tomolillo, D.; Di Lella, F.M.; Trancassini, M.; Faino, L.; Venditti, M.; Antonelli, G.; et al. Evolutionary Trajectories toward Ceftazidime-Avibactam Resistance in Klebsiella pneumoniae Clinical Isolates. Antimicrob. Agents Chemother. 2021, 65, e0057421. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an Update of CRISRFinder, Includes a Portable Version, Enhanced Performance and Integrates Search for Cas Proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Starikova, E.V.; Tikhonova, P.O.; Prianichnikov, N.A.; Rands, C.M.; Zdobnov, E.M.; Ilina, E.N.; Govorun, V.M. Phigaro: High-Throughput Prophage Sequence Annotation. Bioinformatics 2020, 36, 3882–3884. [Google Scholar] [CrossRef]
- Dieckmann, M.A.; Beyvers, S.; Nkouamedjo-Fankep, R.C.; Hanel, P.H.G.; Jelonek, L.; Blom, J.; Goesmann, A. EDGAR3.0: Comparative Genomics and Phylogenomics on a Scalable Infrastructure. Nucleic Acids Res. 2021, 49, W185–W192. [Google Scholar] [CrossRef]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and Standardized Annotation of Bacterial Genomes via Alignment-Free Sequence Identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef]
- Nuccio, S.-P.; Bäumler, A.J. Evolution of the Chaperone/Usher Assembly Pathway: Fimbrial Classification Goes Greek. Microbiol. Mol. Biol. Rev. 2007, 71, 551–575. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap. Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef] [PubMed]
- Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis|Bioinformatics|Oxford Academic. Available online: https://academic.oup.com/bioinformatics/article/31/22/3691/240757?login=true (accessed on 9 September 2022).
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PT | ID | SS | MIC mg/L | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AK | P/T | CAZ | FOX | CRM | CZA | C/T | IMI | MEM | CIP | SXT | AZT | FOS | |||
| Pt#1 | 41 * | RS | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 80 | ≤1 | >64 |
| Pt#2 | 38 | RS | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | 2 | >64 |
| Pt#3 | 883 * | TBA | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | ≤1 | >64 |
| 919 | UR | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | ≤1 | >64 | |
| Pt#4 | 65 * | UR | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | ≤1 | >64 |
| 607 | TBA | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | 2 | >64 | |
| 317 | TBA | 32 | >16 | >32 | >16 | >8 | >16 | >32 | >16 | >16 | >4 | 40 | ≤1 | >64 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capitani, V.; Arcari, G.; Oliva, A.; Sacco, F.; Menichincheri, G.; Fenske, L.; Polani, R.; Raponi, G.; Antonelli, G.; Carattoli, A. Genome-Based Retrospective Analysis of a Providencia stuartii Outbreak in Rome, Italy: Broad Spectrum IncC Plasmids Spread the NDM Carbapenemase within the Hospital. Antibiotics 2023, 12, 943. https://doi.org/10.3390/antibiotics12050943
Capitani V, Arcari G, Oliva A, Sacco F, Menichincheri G, Fenske L, Polani R, Raponi G, Antonelli G, Carattoli A. Genome-Based Retrospective Analysis of a Providencia stuartii Outbreak in Rome, Italy: Broad Spectrum IncC Plasmids Spread the NDM Carbapenemase within the Hospital. Antibiotics. 2023; 12(5):943. https://doi.org/10.3390/antibiotics12050943
Chicago/Turabian StyleCapitani, Valerio, Gabriele Arcari, Alessandra Oliva, Federica Sacco, Gaia Menichincheri, Linda Fenske, Riccardo Polani, Giammarco Raponi, Guido Antonelli, and Alessandra Carattoli. 2023. "Genome-Based Retrospective Analysis of a Providencia stuartii Outbreak in Rome, Italy: Broad Spectrum IncC Plasmids Spread the NDM Carbapenemase within the Hospital" Antibiotics 12, no. 5: 943. https://doi.org/10.3390/antibiotics12050943
APA StyleCapitani, V., Arcari, G., Oliva, A., Sacco, F., Menichincheri, G., Fenske, L., Polani, R., Raponi, G., Antonelli, G., & Carattoli, A. (2023). Genome-Based Retrospective Analysis of a Providencia stuartii Outbreak in Rome, Italy: Broad Spectrum IncC Plasmids Spread the NDM Carbapenemase within the Hospital. Antibiotics, 12(5), 943. https://doi.org/10.3390/antibiotics12050943