
Rapid Screening and Comparison of Chimeric Lysins for Antibacterial Activity against Staphylococcus aureus Strains

 , , and
, , and
Abstract
1. Introduction
2. Results
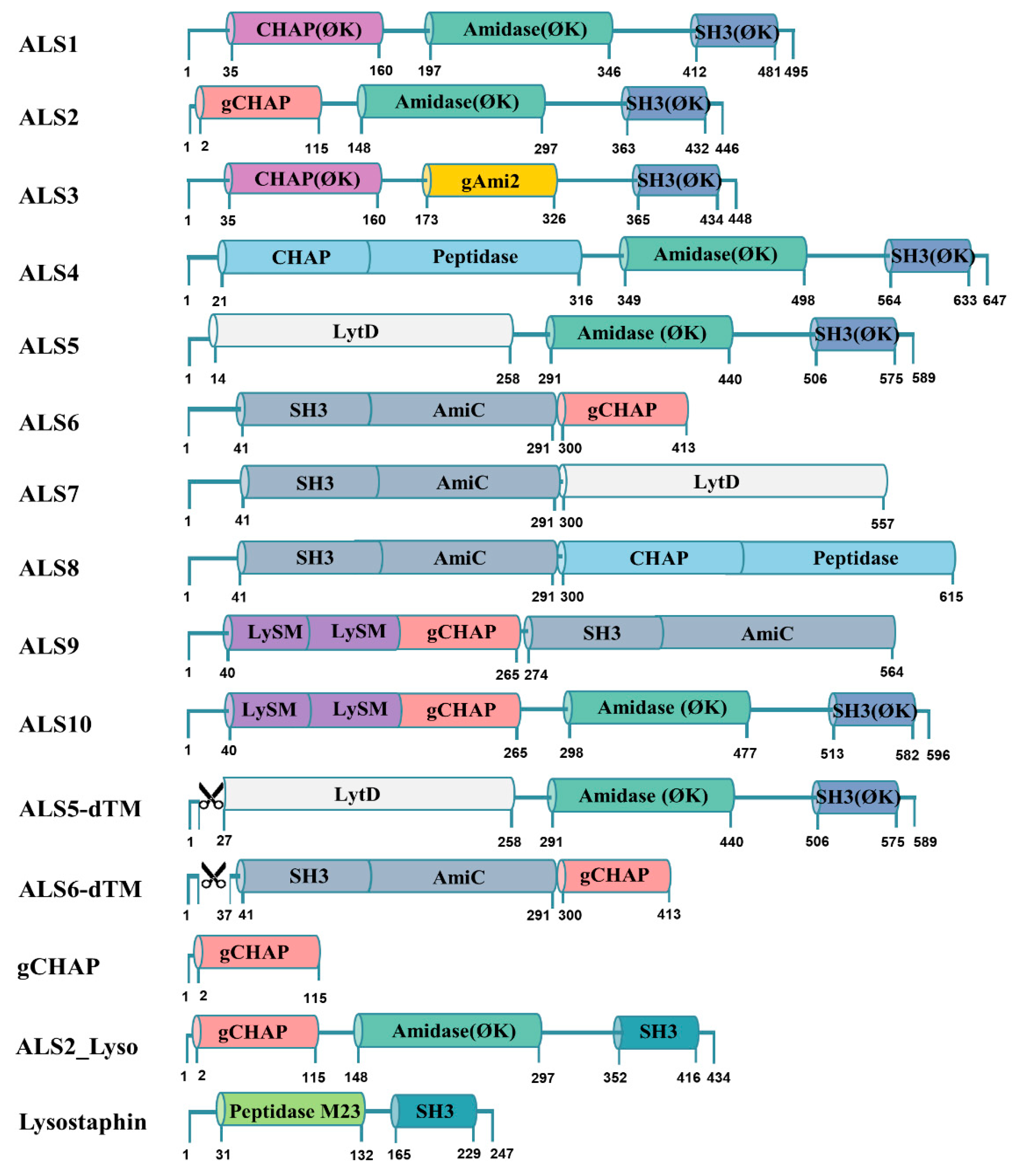
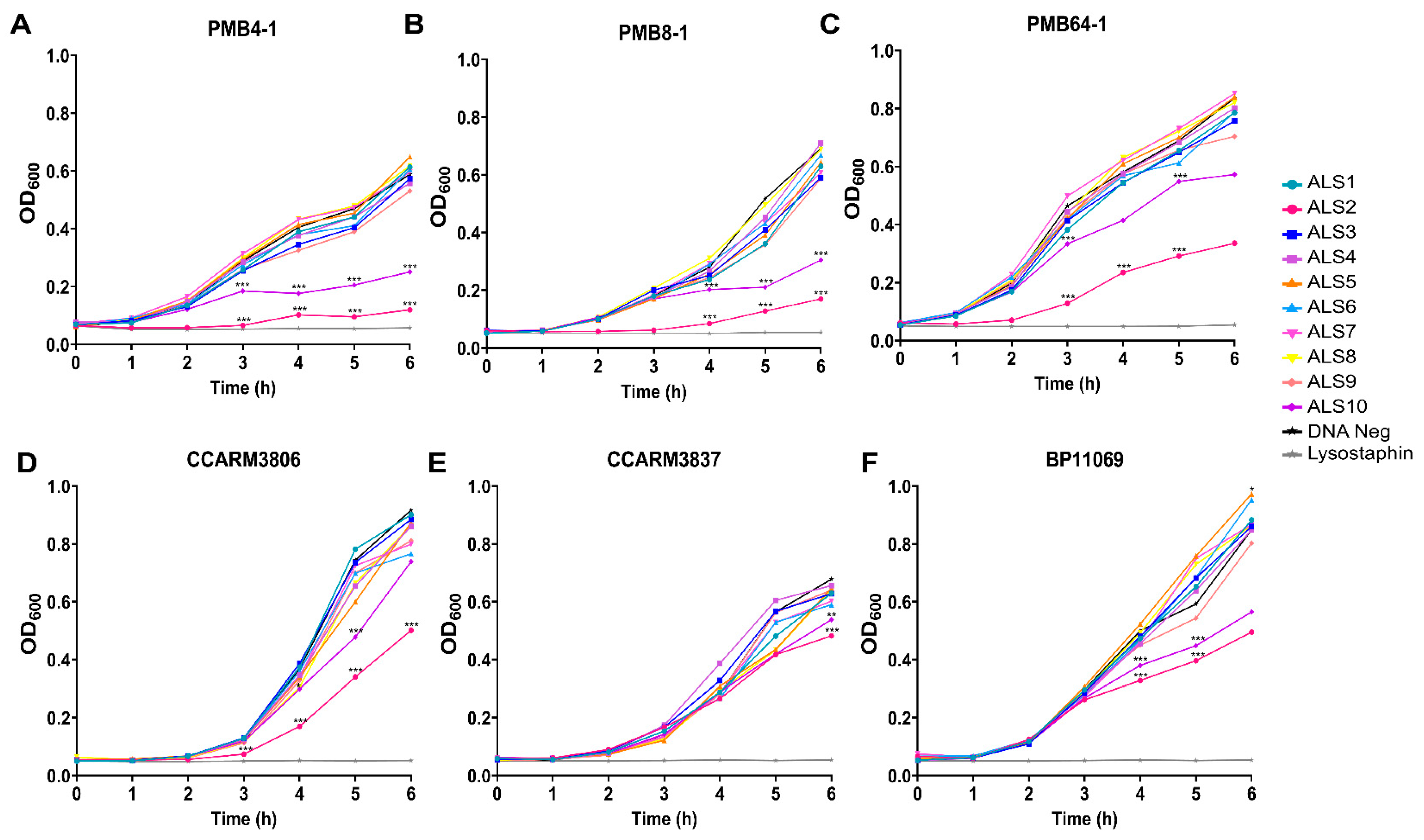
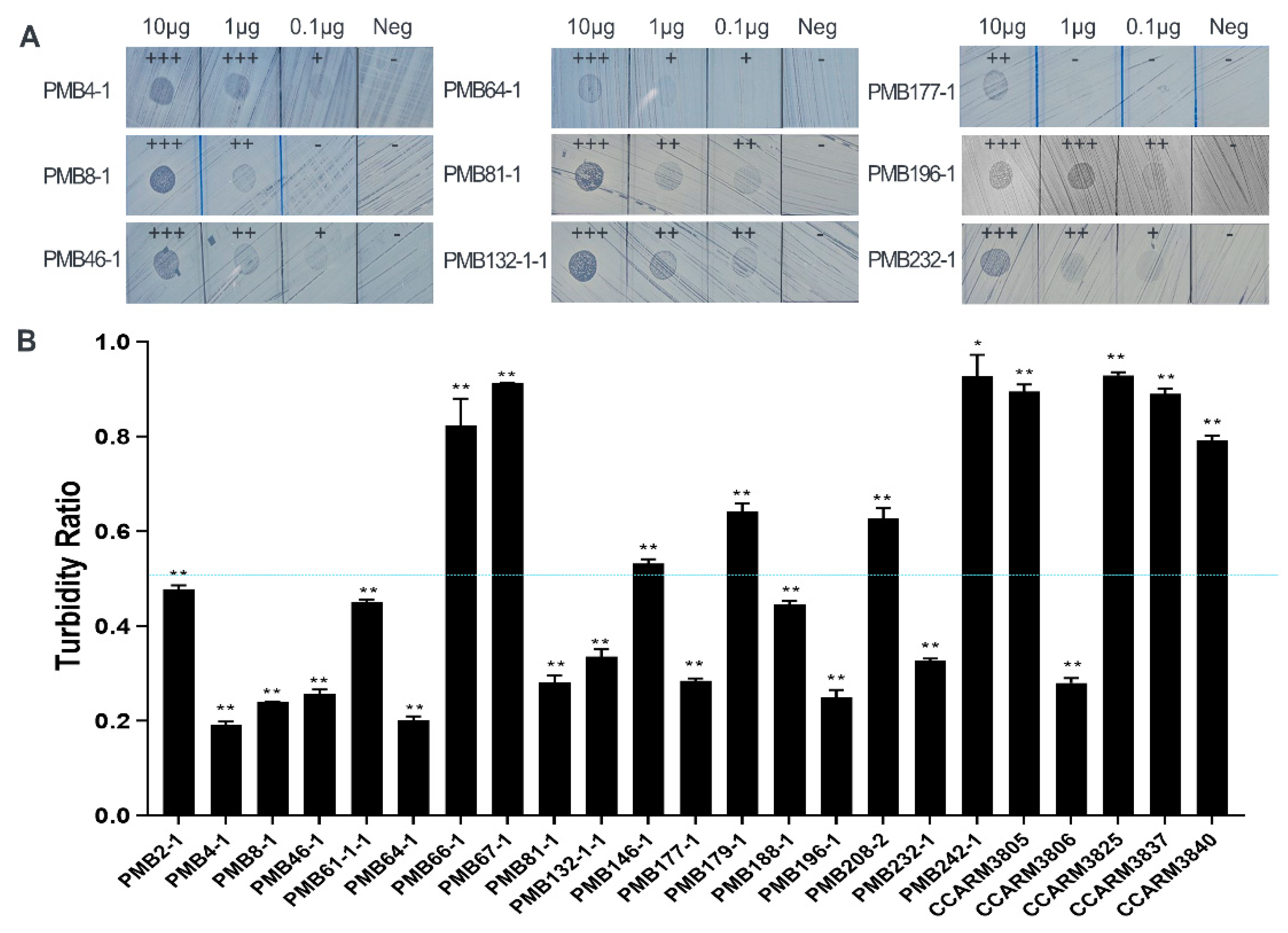
2.1. Rapid Antibacterial Activity Screening and Comparison of Chimeric Lysin Candidates
2.2. Comparison of Susceptibility of Various S. aureus Strains to ALS2 Using Cell-Free Expression System
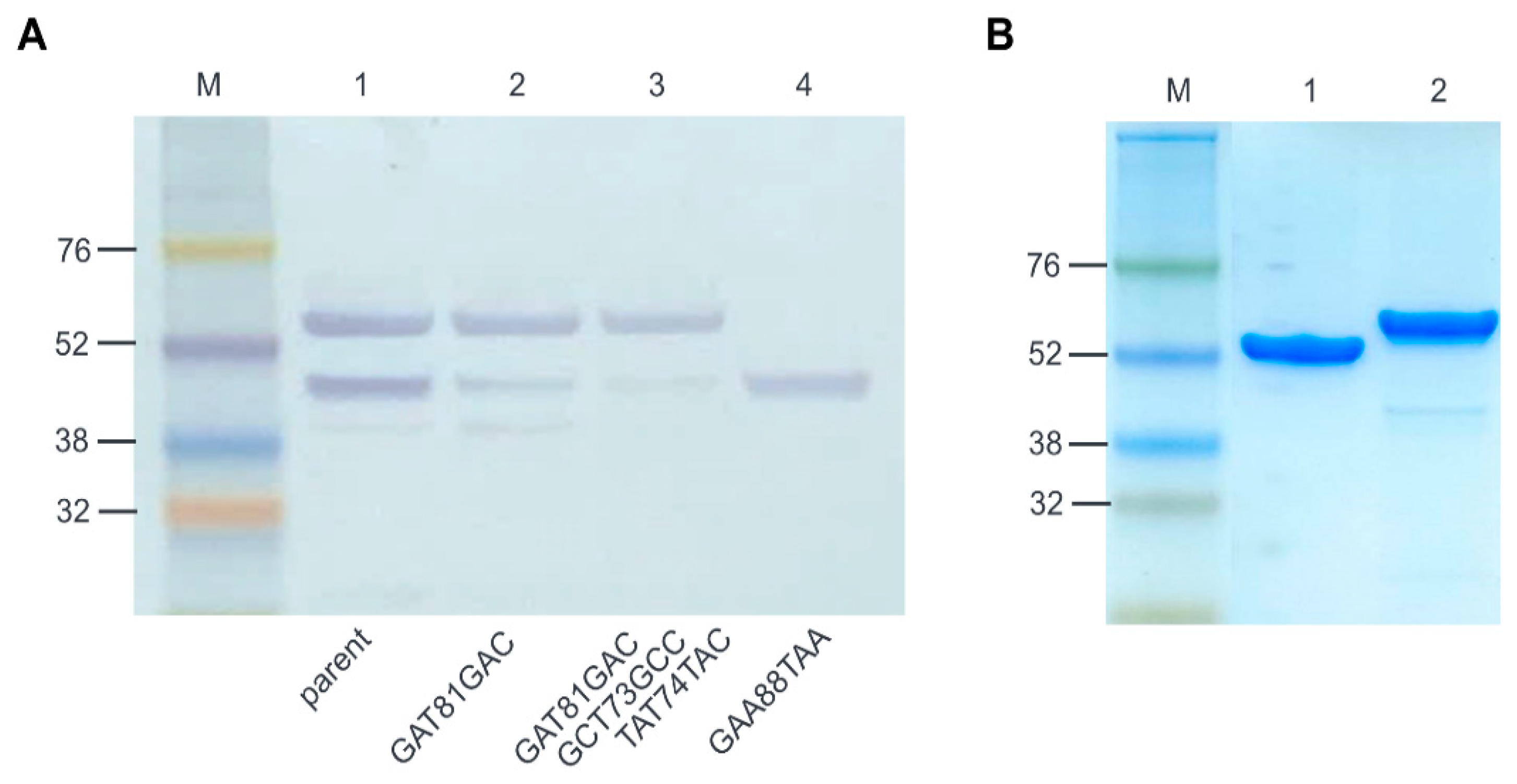
2.3. Optimization of E. coli Expression of ALS2
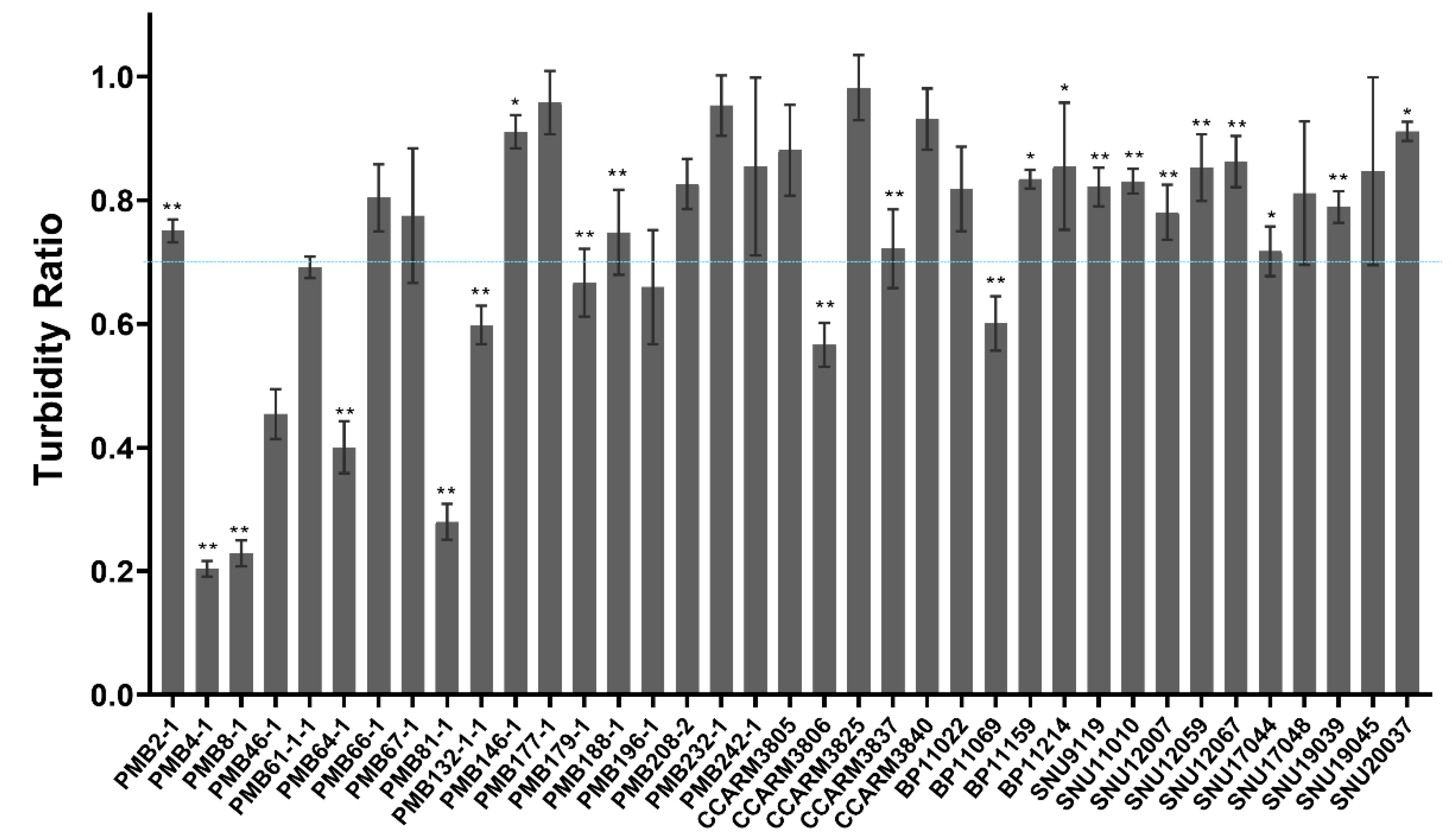
2.4. Comparison of Susceptibility of Bovine and Human S. aureus Strains to E. coli-Expressed and Highly Purified ALS2
2.5. Accordance of Bacterial Susceptibility to ALS2 Expressed in Cell-Free and E. coli Expression Systems
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Bioinformatics
4.3. Design and Construction of Chimeric Lysin Gene for Cell-Free Expression
4.4. Site-Directed Mutagenesis
4.5. Cell-Free Protein Synthesis
4.6. Antibacterial Activity Tests of Chimeric Lysins
4.7. E. coli Expression and Protein Purification
4.8. SDS-PAGE and Western Blotting
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burton, J.L.; Erskine, R.J. Immunity and mastitis Some new ideas for an old disease. Vet. Clin. NorthAm. Food Anim. Pract. 2003, 19, 1–45. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar] [PubMed]
- McNamee, P.T.; Smyth, J.A. Bacterial chondronecrosis with osteomyelitis (‘femoral head necrosis’) of broiler chickens: A review. Avian Pathol. 2000, 29, 477–495. [Google Scholar] [CrossRef] [PubMed]
- Jevons, M.P. “Celbenin”-resistant staphylococci. Br. Med. J. 1961, 1, 124. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Cui, L.; Kuroda, M.; Ito, T. The emergence and evolution of methicillin-resistant Staphylococcus aureus. Trends Microbiol. 2001, 9, 486–493. [Google Scholar] [CrossRef]
- Ko, D.-S.; Kim, N.-H.; Kim, E.-K.; Ha, E.-J.; Ro, Y.-H.; Kim, D.; Choi, K.-S.; Kwon, H.-J. Comparative genomics of bovine mastitis-origin Staphylococcus aureus strains classified into prevalent human genotypes. Res. Vet. Sci. 2021, 139, 67–77. [Google Scholar] [CrossRef]
- Guinane, C.M.; Ben Zakour, N.L.; Tormo-Mas, M.A.; Weinert, L.A.; Lowder, B.V.; Cartwright, R.A.; Smyth, D.S.; Smyth, C.J.; Lindsay, J.A.; Gould, K.A. Evolutionary genomics of Staphylococcus aureus reveals insights into the origin and molecular basis of ruminant host adaptation. Genome Biol. Evol. 2010, 2, 454–466. [Google Scholar] [CrossRef]
- Matuszewska, M.; Murray, G.G.R.; Harrison, E.M.; Holmes, M.A.; Weinert, L.A. The Evolutionary Genomics of Host Specificity in Staphylococcus aureus. Trends Microbiol. 2020, 28, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef]
- Weinert, L.A.; Welch, J.J.; Suchard, M.A.; Lemey, P.; Rambaut, A.; Fitzgerald, J.R. Molecular dating of human-to-bovid host jumps by Staphylococcus aureus reveals an association with the spread of domestication. Biol. Lett. 2012, 8, 829–832. [Google Scholar] [CrossRef]
- Seong, W.-J.; Kim, J.-H.; Kwon, H.-J. Comparison of complete rpoB gene sequence typing and multi-locus sequence typing for phylogenetic analysis of Staphylococcus aureus. J. Gen. Appl. Microbiol. 2013, 59, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.-S.; Seong, W.-J.; Kim, D.; Kim, E.-K.; Kim, N.-H.; Lee, C.-Y.; Kim, J.-H.; Kwon, H.-J. Molecular prophage typing of Staphylococcus aureus isolates from bovine mastitis. J. Vet. Sci. 2018, 19, 771–781. [Google Scholar] [CrossRef]
- Ko, D.-S.; Kim, D.; Kim, E.-K.; Kim, J.-H.; Kwon, H.-J. Evolution of a major bovine mastitic genotype (rpoB sequence type 10-2) of Staphylococcus aureus in cows. J. Microbiol. 2019, 57, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M. Bacterial responses to osmotic challenges. J. Gen. Physiol. 2015, 145, 381–388. [Google Scholar] [CrossRef]
- Loskill, P.; Pereira, P.M.; Jung, P.; Bischoff, M.; Herrmann, M.; Pinho, M.G.; Jacobs, K. Reduction of the peptidoglycan crosslinking causes a decrease in stiffness of the Staphylococcus aureus cell envelope. Biophys. J. 2014, 107, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Matias, V.R.; Beveridge, T.J. Native cell wall organization shown by cryo-electron microscopy confirms the existence of a periplasmic space in Staphylococcus aureus. J. Bacteriol. 2006, 188, 1011–1021. [Google Scholar] [CrossRef]
- Vollmer, W.; Blanot, D.; De Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef]
- Tyagi, J.L.; Sharma, M.; Gulati, K.; Kairamkonda, M.; Kumar, D.; Poluri, K.M. Engineering of a T7 Bacteriophage Endolysin Variant with Enhanced Amidase Activity. Biochemistry 2023, 62, 330–344. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol 2012, 7, 1147–1171. [Google Scholar] [CrossRef]
- Ho, M.K.Y.; Zhang, P.; Chen, X.; Xia, J.; Leung, S.S.Y. Bacteriophage endolysins against gram-positive bacteria, an overview on the clinical development and recent advances on the delivery and formulation strategies. Crit. Rev. Microbiol. 2022, 48, 303–326. [Google Scholar] [CrossRef]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef]
- O’flaherty, S.; Coffey, A.; Meaney, W.; Fitzgerald, G.; Ross, R.P. The recombinant phage lysin LysK has a broad spectrum of lytic activity against clinically relevant staphylococci, including methicillin-resistant Staphylococcus aureus. J. Bacteriol. 2005, 187, 7161–7164. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Coffey, A.; Edwards, R.; Meaney, W.; Fitzgerald, G.F.; Ross, R.P. Genome of staphylococcal phage K: A new lineage of Myoviridae infecting gram-positive bacteria with a low G+C content. J. Bacteriol. 2004, 186, 2862–2871. [Google Scholar] [CrossRef] [PubMed]
- Low, L.Y.; Yang, C.; Perego, M.; Osterman, A.; Liddington, R. Role of net charge on catalytic domain and influence of cell wall binding domain on bactericidal activity, specificity, and host range of phage lysins. J. Biol. Chem. 2011, 286, 34391–34403. [Google Scholar] [CrossRef]
- Becker, S.C.; Foster-Frey, J.; Donovan, D.M. The phage K lytic enzyme LysK and lysostaphin act synergistically to kill MRSA. FEMS Microbiol. Lett. 2008, 287, 185–191. [Google Scholar] [CrossRef]
- Leonard, A.C.; Goncheva, M.I.; Gilbert, S.E.; Shareefdeen, H.; Petrie, L.E.; Thompson, L.K.; Khursigara, C.M.; Heinrichs, D.E.; Cox, G. Autolysin-mediated peptidoglycan hydrolysis is required for the surface display of Staphylococcus aureus cell wall-anchored proteins. Proc. Natl. Acad. Sci. USA 2023, 120, e2301414120. [Google Scholar] [CrossRef]
- Rupp, M.E.; Fey, P.D.; Heilmann, C.; Götz, F. Characterization of the importance of Staphylococcus epidermidis autolysin and polysaccharide intercellular adhesin in the pathogenesis of intravascular catheter-associated infection in a rat model. J. Infect. Dis. 2001, 183, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, S.X.; St. John, A.; Torres, V.J. The Major Autolysin Atl Regulates the Virulence of Staphylococcus aureus by Controlling the Sorting of LukAB. Infect. Immun. 2022, 90, e0005622. [Google Scholar] [CrossRef] [PubMed]
- Farrand, A.J.; Haley, K.P.; Lareau, N.M.; Heilbronner, S.; McLean, J.A.; Foster, T.; Skaar, E.P. An iron-regulated autolysin remodels the cell wall to facilitate heme acquisition in Staphylococcus lugdunensis. Infect. Immun. 2015, 83, 3578–3589. [Google Scholar] [CrossRef]
- Layec, S.; Decaris, B.; Leblond-Bourget, N. Diversity of Firmicutes peptidoglycan hydrolases and specificities of those involved in daughter cell separation. Res. Microbiol. 2008, 159, 507–515. [Google Scholar] [CrossRef]
- Kajimura, J.; Fujiwara, T.; Yamada, S.; Suzawa, Y.; Nishida, T.; Oyamada, Y.; Hayashi, I.; Yamagishi, J.i.; Komatsuzawa, H.; Sugai, M. Identification and molecular characterization of an N-acetylmuramyl-l-alanine amidase Sle1 involved in cell separation of Staphylococcus aureus. Mol. Microbiol. 2005, 58, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Biswas, R.; Voggu, L.; Simon, U.K.; Hentschel, P.; Thumm, G.; Götz, F. Activity of the major staphylococcal autolysin Atl. FEMS Microbiol. Lett. 2006, 259, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Murakami, K. Staphylococcus aureus clinical isolate with high-level methicillin resistance with an lytH mutation caused by IS 1182 insertion. Antimicrob. Agents Chemother. 2008, 52, 643–647. [Google Scholar] [CrossRef]
- Osipovitch, D.C.; Therrien, S.; Griswold, K.E. Discovery of novel S. aureus autolysins and molecular engineering to enhance bacteriolytic activity. Appl. Microbiol. Biotechnol. 2015, 99, 6315–6326. [Google Scholar] [CrossRef]
- Ning, H.; Lin, H.; Wang, J.; He, X.; Lv, X.; Ju, L. Characterizations of the endolysin Lys84 and its domains from phage qdsa002 with high activities against Staphylococcus aureus and its biofilms. Enzym. Microb. Technol. 2021, 148, 109809. [Google Scholar] [CrossRef]
- Son, B.; Kong, M.; Cha, Y.; Bai, J.; Ryu, S. Simultaneous Control of Staphylococcus aureus and Bacillus cereus Using a Hybrid Endolysin LysB4EAD-LysSA11. Antibiotics 2020, 9, 906. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, J.; Nakamura, T.; Furusawa, T.; Ohno, H.; Takahashi, H.; Kitana, J.; Usui, M.; Higuchi, H.; Tanji, Y.; Tamura, Y.; et al. Characterization of the Lytic Capability of a LysK-Like Endolysin, Lys-phiSA012, Derived from a Polyvalent Staphylococcus aureus Bacteriophage. Pharmaceuticals 2018, 11, 25. [Google Scholar] [CrossRef]
- Park, J.-M. Development of a Novel Chimeric Lysin(gCHAP-LysK) against Staphylococcus aureus. Master’s Thesis, Seoul National University, Seoul, Republic of Korea, 2019. [Google Scholar]
- Seong, W.-J.; Kim, D.; Kim, E.-K.; Ko, D.-S.; Ro, Y.; Kim, J.-H.; Kwon, H.-J. Molecular identification of coagulase-negative staphylococci by rpoB sequence typing. Korean J. Vet. Res. 2018, 58, 51–55. [Google Scholar] [CrossRef]
- Tang, G.-Q.; Bandwar, R.P.; Patel, S.S. Extended upstream AT sequence increases T7 promoter strength. J. Biol. Chem. 2005, 280, 40707–40713. [Google Scholar] [CrossRef]
- Calvopina-Chavez, D.G.; Gardner, M.A.; Griffitts, J.S. Engineering efficient termination of bacteriophage T7 RNA polymerase transcription. G3 2022, 12, jkac070. [Google Scholar] [CrossRef]
- Komarova, E.S.; Chervontseva, Z.S.; Osterman, I.A.; Evfratov, S.A.; Rubtsova, M.P.; Zatsepin, T.S.; Semashko, T.A.; Kostryukova, E.S.; Bogdanov, A.A.; Gelfand, M.S. Influence of the spacer region between the Shine–Dalgarno box and the start codon for fine-tuning of the translation efficiency in Escherichia coli. Microb. Biotechnol. 2020, 13, 1254–1261. [Google Scholar] [CrossRef]
- Horgan, M.; O’Flynn, G.; Garry, J.; Cooney, J.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P.; McAuliffe, O. Phage lysin LysK can be truncated to its CHAP domain and retain lytic activity against live antibiotic-resistant staphylococci. Appl. Environ. Microbiol. 2009, 75, 872–874. [Google Scholar] [CrossRef]
- Hecht, A.; Glasgow, J.; Jaschke, P.R.; Bawazer, L.A.; Munson, M.S.; Cochran, J.R.; Endy, D.; Salit, M. Measurements of translation initiation from all 64 codons in E. coli. Nucleic Acids Res. 2017, 45, 3615–3626. [Google Scholar] [CrossRef]
- Sanz-Gaitero, M.; Keary, R.; Garcia-Doval, C.; Coffey, A.; Van Raaij, M.J. Crystal structure of the lytic CHAPK domain of the endolysin LysK from Staphylococcus aureus bacteriophage K. Virol. J. 2014, 11, 133. [Google Scholar] [CrossRef]
- Zou, Y.; Hou, C. Systematic analysis of an amidase domain CHAP in 12 Staphylococcus aureus genomes and 44 staphylococcal phage genomes. Comput. Biol. Chem. 2010, 34, 251–257. [Google Scholar] [CrossRef]
- Jayakumar, J.; Kumar, V.A.; Biswas, L.; Biswas, R. Therapeutic applications of lysostaphin against Staphylococcus aureus. J. Appl. Microbiol. 2021, 131, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, J.; Son, B.; Ryu, S. Development of Advanced Chimeric Endolysin to Control Multidrug-Resistant Staphylococcus aureus through Domain Shuffling. ACS Infect. Dis. 2021, 7, 2081–2092. [Google Scholar] [CrossRef] [PubMed]
- Tossavainen, H.; Raulinaitis, V.; Kauppinen, L.; Pentikäinen, U.; Maaheimo, H.; Permi, P. Structural and functional insights into lysostaphin–substrate interaction. Front. Mol. Biosci. 2018, 5, 60. [Google Scholar] [CrossRef]
- Sharif, S.; Singh, M.; Kim, S.J.; Schaefer, J. Staphylococcus aureus peptidoglycan tertiary structure from carbon-13 spin diffusion. J. Am. Chem. Soc. 2009, 131, 7023–7030. [Google Scholar] [CrossRef]
- Donovan, D.M.; Foster-Frey, J. LambdaSa2 prophage endolysin requires Cpl-7-binding domains and amidase-5 domain for antimicrobial lysis of streptococci. FEMS Microbiol. Lett. 2008, 287, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Louzoun Zada, S.; Green, K.D.; Shrestha, S.K.; Herzog, I.M.; Garneau-Tsodikova, S.; Fridman, M. Derivatives of Ribosome-Inhibiting Antibiotic Chloramphenicol Inhibit the Biosynthesis of Bacterial Cell Wall. ACS Infect. Dis. 2018, 4, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Espaillat, A.; Cava, F. Bacterial strategies to preserve cell wall integrity against environmental threats. Front. Microbiol. 2018, 9, 2064. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Dong, S.; Baker, J.R.; Foster-Frey, J.; Pritchard, D.G.; Donovan, D.M. LysK CHAP endopeptidase domain is required for lysis of live staphylococcal cells. FEMS Microbiol. Lett. 2009, 294, 52–60. [Google Scholar] [CrossRef]
- Park, S.; Jun, S.Y.; Kim, C.-H.; Jung, G.M.; Son, J.S.; Jeong, S.T.; Yoon, S.J.; Lee, S.Y.; Kang, S.H. Characterisation of the antibacterial properties of the recombinant phage endolysins AP50-31 and LysB4 as potent bactericidal agents against Bacillus anthracis. Sci. Rep. 2018, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Duman, Z.E.; Ünlü, A.; Çakar, M.M.; Ünal, H.; Binay, B. Enhanced production of recombinant Staphylococcus simulans lysostaphin using medium engineering. Prep. Biochem. Biotechnol. 2019, 49, 521–528. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Szweda, P.; Kotłowski, R.; Kur, J. New effective sources of the Staphylococcus simulans lysostaphin. J. Biotechnol. 2005, 117, 203–213. [Google Scholar] [CrossRef]
- Heckman, K.L.; Pease, L.R. Gene splicing and mutagenesis by PCR-driven overlap extension. Nat. Protoc. 2007, 2, 924–932. [Google Scholar] [CrossRef]
- Donovan, D.M.; Dong, S.; Garrett, W.; Rousseau, G.M.; Moineau, S.; Pritchard, D.G. Peptidoglycan hydrolase fusions maintain their parental specificities. Appl. Environ. Microbiol. 2006, 72, 2988–2996. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Origin a | RST b | MRSA Gene | Plate Lysis Test c | Turbidity Reduction Test | Ref | |||
|---|---|---|---|---|---|---|---|---|---|
| 10 µg | 1 µg | 0.1 µg | E. coli d | Cell-Free e | |||||
| PMB2-1 | BM | 10-2 | - | ++ c | + | - | + | - f | [14] |
| PMB4-1 | BM | 22-1 | - | +++ b | +++ | + | + | + f | |
| PMB8-1 | BM | 8-1 | - | +++ | ++ | - | + | + f | |
| PMB46-1 | BM | 10-2 | - | +++ | ++ | + | + | + | |
| PMB61-1-1 | BM | 10-2 | - | ++ | + | + | + | + | |
| PMB64-1 | BM | 10-2 | - | +++ | +++ | ++ | + | + f | |
| PMB66-1 | BM | 14-3 | - | - | - | - | - | - g | |
| PMB67-1 | BM | 5-2 | - | + | - | - | - | - f | |
| PMB81-1 | BM | 10-2 | - | +++ | ++ | ++ | + | + | |
| PMB132-1-1 | BM | 10-2 | - | +++ | ++ | ++ | + | + | |
| PMB146-1 | BM | 11-5 | - | +++ | ++ | - | + | + | |
| PMB177-1 | BM | 14-2 | - | ++ | + | - | + | + | |
| PMB179-1 | BM | 4-1 | - | + | - | - | - | + f | |
| PMB188-1 | BM | 10-3 | - | ++ | + | - | + | - | |
| PMB196-1 | BM | 2-1 | - | +++ | ++ | ++ | + | + g | |
| PMB208-2 | BM | 11-6 | - | - | - | - | - | - | |
| PMB232-1 | BM | 10-2 | - | +++ | ++ | + | + | - | |
| PMB242-1 | BM | 11-4 | - | nt f | nt | nt | - | - | |
| CCARM3805 | HI | 2-1 | + | - | - | - | - | - | |
| CCARM3806 | HI | 10-2 | + | nt | nt | nt | + | + f | |
| CCARM3825 | HI | 2-1 | + | nt | nt | nt | - | - | |
| CCARM3837 | HI | 4-1 | + | - | - | - | - | - g | |
| CCARM3840 | HI | 4-1 | + | nt | nt | nt | - | - g | |
| BP11022 | CI | 3-1 | - | nt | nt | nt | nt | - | [13] |
| BP11069 | CI | 6-3 | + | nt | nt | nt | nt | + g | |
| BP11159 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| BP11214 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU9119 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU11010 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU12007 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU12059 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU12067 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU17044 | CI | 3-1 | - | nt | nt | nt | nt | - | This study |
| SNU17048 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU19039 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU19045 | CI | 3-1 | - | nt | nt | nt | nt | - | |
| SNU20037 | CI | 3-1 | - | nt | nt | nt | nt | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-M.; Ko, D.-S.; Kim, H.-S.; Kim, N.-H.; Kim, E.-K.; Roh, Y.-H.; Kim, D.; Kim, J.-H.; Choi, K.-S.; Kwon, H.-J. Rapid Screening and Comparison of Chimeric Lysins for Antibacterial Activity against Staphylococcus aureus Strains. Antibiotics 2023, 12, 667. https://doi.org/10.3390/antibiotics12040667
Park J-M, Ko D-S, Kim H-S, Kim N-H, Kim E-K, Roh Y-H, Kim D, Kim J-H, Choi K-S, Kwon H-J. Rapid Screening and Comparison of Chimeric Lysins for Antibacterial Activity against Staphylococcus aureus Strains. Antibiotics. 2023; 12(4):667. https://doi.org/10.3390/antibiotics12040667
Chicago/Turabian StylePark, Jin-Mi, Dae-Sung Ko, Hee-Soo Kim, Nam-Hyung Kim, Eun-Kyoung Kim, Young-Hye Roh, Danil Kim, Jae-Hong Kim, Kang-Seuk Choi, and Hyuk-Joon Kwon. 2023. "Rapid Screening and Comparison of Chimeric Lysins for Antibacterial Activity against Staphylococcus aureus Strains" Antibiotics 12, no. 4: 667. https://doi.org/10.3390/antibiotics12040667
APA StylePark, J.-M., Ko, D.-S., Kim, H.-S., Kim, N.-H., Kim, E.-K., Roh, Y.-H., Kim, D., Kim, J.-H., Choi, K.-S., & Kwon, H.-J. (2023). Rapid Screening and Comparison of Chimeric Lysins for Antibacterial Activity against Staphylococcus aureus Strains. Antibiotics, 12(4), 667. https://doi.org/10.3390/antibiotics12040667