
Effect of Quorum Sensing Molecule Farnesol on Mixed Biofilms of Candida albicans and Staphylococcus aureus

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Characterization of Strains in Terms of Identification, Resistance and Biofilm Formation
2.2. Effect of FAR and the Combination of FAR with Antibiotics to S. aureus Isolates
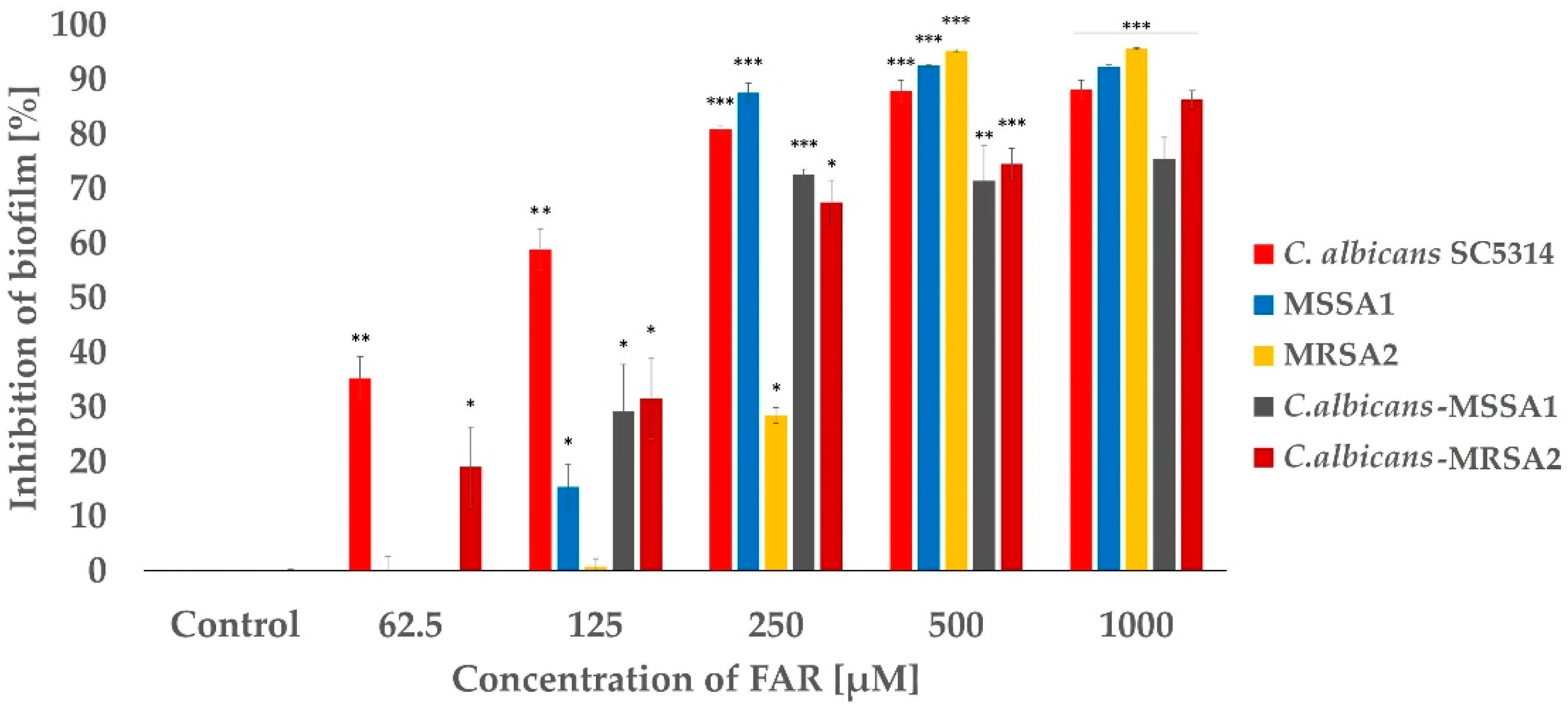
2.3. Study of the Effect of FAR and the Combination of FAR and OXA on Mono-Species and Mixed Biofilms Formed by Candida albicans-MSSA or MRSA
2.4. Microscopic Evaluation of the Effect of FAR and the Combination of FAR with OXA on Mixed Biofilms
3. Discussion
4. Materials and Methods
4.1. Characterization of Microbial Strains
4.2. Antimicrobial Susceptibilities and Resistance Signatures of S. aureus Strains
4.3. Biofilm Assay
4.4. Susceptibility Testing of FAR to Planktonic S. aureus
4.5. Susceptibility Testing of FAR with Antibiotics on Planktonic S. aureus
4.6. Susceptibility Testing of FAR on Mono-Species and Mixed Biofilms
4.7. Susceptibility Testing of FAR/OXA Combination on Mono-Species and Mixed Biofilms
4.8. Microscopic Analysis of Mixed Biofilms
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peters, B.M.; Ovchinnikova, E.S.; Krom, B.P.; Schlecht, L.M.; Zhou, H.; Hoyer, L.L.; Busscher, H.J.; van der Mei, H.C.; Jabra-Rizk, M.A.; Shirtliff, M.E. Staphylococcus aureus adherence to Candida albicans hyphae is mediated by the hyphal adhesin Als3p. Microbiology 2012, 158, 2975–2986. [Google Scholar] [CrossRef] [PubMed]
- Pinto, H.; Simões, M.; Borges, A. Prevalence and Impact of Biofilms on Bloodstream and Urinary Tract Infections: A Systematic Review and Meta-Analysis. Antibiotics 2021, 10, 825. [Google Scholar] [CrossRef] [PubMed]
- Edmiston, C.E.; McBain, A.J.; Roberts, C.; Leaper, D. Clinical and Microbiological Aspects of Biofilm-Associated Surgical Site Infections. In Biofilm-Based Healthcare-Associated Infections; Donelli, G., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2015; pp. 47–67. ISBN 978-3-319-11038-7. [Google Scholar]
- Vandecandelaere, I.; Matthijs, N.; van Nieuwerburgh, F.; Deforce, D.; Vosters, P.; de Bus, L.; Nelis, H.J.; Depuydt, P.; Coenye, T. Assessment of Microbial Diversity in Biofilms Recovered from Endotracheal Tubes Using Culture Dependent and Independent Approaches. PLoS ONE 2012, 7, e38401. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Surveillance of Healthcare-Associated Infections and Prevention Indicators in European Intensive Care Units Year: Hai-Net Icu Protocol; Version 2.2; ECDC: Stockholm, Sweden, 2017; Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-healthcare-associated-infections-and-prevention-indicators-european (accessed on 23 September 2021).
- Carolus, H.; Van Dyck, K.; Van Dijck, P. Candida albicans and Staphylococcus species: A threatening twosome. Front. Microbiol. 2019, 10, 2162. [Google Scholar] [CrossRef] [PubMed]
- Todd, O.A.; Peters, B.M. Candida albicans and Staphylococcus aureus pathogenicity and polymicrobial interactions: Lessons beyond koch’s postulates. J. Fungi 2019, 5, 81. [Google Scholar] [CrossRef]
- Kean, R.; Rajendran, R.; Haggarty, J.; Townsend, E.M.; Short, B.; Burgess, K.E.; Lang, S.; Millington, O.; Mackay, W.G.; Williams, C.; et al. Candida albicans Mycofilms Support Staphylococcus aureus Colonization and Enhances Miconazole Resistance in Dual-Species Interactions. Front. Microbiol. 2017, 23, 258. [Google Scholar] [CrossRef]
- Harriott, M.M.; Noverr, M.C. Candida albicans and Staphylococcus aureus form polymicrobial biofilms: Effects on antimicrobial resistance. Antimicrob. Agents Chemother. 2009, 53, 3914–3922. [Google Scholar] [CrossRef]
- Harriott, M.M.; Noverr, M.C. Ability of Candida albicans mutants to induce Staphylococcus aureus vancomycin resistance during polymicrobial biofilm formation. Antimicrob Agents Chemother. 2010, 54, 3746–3755. [Google Scholar] [CrossRef]
- Kong, E.F.; Tsui, C.; Kucharíková, S.; Andes, D.; Van Dijck, P.; Jabra-Rizk, M.A. Commensal Protection of Staphylococcus aureus against Antimicrobials by Candida albicans Biofilm Matrix. MBio 2016, 7, e01365-16. [Google Scholar] [CrossRef]
- Adam, B.; Baillie, G.S.; Douglas, L.J. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J. Med. Microbiol. 2002, 51, 344–349. [Google Scholar] [CrossRef]
- Tuon, F.F.; Suss, P.H.; Telles, J.P.; Dantas, L.R.; Borges, N.H.; Ribeiro, V.S.T. Antimicrobial Treatment of Staphylococcus aureus Biofilms. Antibiotics 2023, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, X. Reevaluation of lock solutions for Central venous catheters in hemodialysis: A narrative review. Ren. Fail. 2022, 44, 1501–1518. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Rodriguez, J.A.; Shaqour, B.; Guarch-Pérez, C.; Choińska, E.; Riool, M.; Verleije, B.; Beyers, K.; Costantini, V.J.A.; Świeszkowski, W.; Zaat, S.A.J.; et al. A Niclosamide-releasing hot-melt extruded catheter prevents Staphylococcus aureus experimental biomaterial-associated infection. Sci. Rep. 2022, 12, 12329. [Google Scholar] [CrossRef] [PubMed]
- Aldilla, V.R.; Chen, R.; Kuppusamy, R.; Chakraborty, S.; Willcox, M.D.P.; Black, D.S.; Thordarson, P.; Martin, A.D.; Kumar, N. Hydrogels with intrinsic antibacterial activity prepared from naphthyl anthranilamide (NaA) capped peptide mimics. Sci. Rep. 2022, 12, 22259. [Google Scholar] [CrossRef]
- Griesser, S.S.; Jasieniak, M.; Vasilev, K.; Griesser, H.J. Antimicrobial Peptides Grafted onto a Plasma Polymer Interlayer Platform: Performance upon Extended Bacterial Challenge. Coatings 2021, 11, 68. [Google Scholar] [CrossRef]
- Shalom, Y.; Perelshtein, I.; Perkas, N.; Gedanken, A.; Banin, E. Catheters Coated with Zn-Doped CuO Nanoparticles Delay the Onset of Catheter-Associated Urinary Tract Infections. Nano Res. 2017, 10, 520–533. [Google Scholar] [CrossRef]
- Kuppusamy, R.; Browne, K.; Suresh, D.; Do Rosario, R.M.; Chakraborty, S.; Yang, S.; Willcox, M.D.; Black, D.S.; Chen, R.; Kumar, N. Transition Towards Antibiotic Hybrid Vehicles: The Next Generation Antibacterials. Curr. Med. Chem. 2022, 30, 104–125. [Google Scholar] [CrossRef]
- Nowacka, M.; Kowalewska, A.; Kręgiel, D. Farnesol-Containing Macromolecular Systems for Antibiofilm Strategies. Surfaces 2020, 3, 197–210. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, J.; Yang, C.; Yin, Y.; Yao, K. Quorum Sensing: A Prospective Therapeutic Target for Bacterial Diseases. Biomed Res. Int. 2019, 2019, 2015978. [Google Scholar] [CrossRef]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial quorum sensing and microbial community interactions. MBio 2018, 9, e02331-17. [Google Scholar] [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal Quorum-Sensing Molecules and Inhibitors with Potential Antifungal Activity: A Review. Molecules 2019, 24, 1950. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Saville, S.P.; Wickes, B.L.; López-Ribot, J.L. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl. Environ. Microbiol. 2002, 68, 5459–5463. [Google Scholar] [CrossRef] [PubMed]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Shiraishi, A.; Hada, T.; Hirose, K.; Hamashima, H.; Shimada, J. The antibacterial effects of terpene alcohols on Staphylococcus aureus and their mode of action. FEMS Microbiol. Lett. 2004, 237, 325–331. [Google Scholar] [CrossRef]
- Akiyama, H.; Oono, T.; Huh, W.K.; Yamasaki, O.; Ogawa, S.; Katsuyama, M.; Ichikawa, H.; Iwatsuki, K. Actions of farnesol and xylitol against Staphylococcus aureus. Chemotherapy 2002, 48, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Dekkerová, J.; Černáková, L.; Kendra, S.; Borghi, E.; Ottaviano, E.; Willinger, B.; Bujdáková, H. Farnesol Boosts the Antifungal Effect of Fluconazole and Modulates Resistance in Candida auris through Regulation of the CDR1 and ERG11 Genes. J. Fungi 2022, 8, 783. [Google Scholar] [CrossRef]
- Sharma, M.; Prasad, R. The Quorum-Sensing Molecule Farnesol Is a Modulator of Drug Efflux Mediated by ABC Multidrug Transporters and Synergizes with Drugs in Candida Albicans. Antimicrob. Agents Chemother. 2011, 55, 4834–4843. [Google Scholar] [CrossRef]
- Kim, C.; Hesek, D.; Lee, M.; Mobashery, S. Potentiation of the activity of β-lactam antibiotics by farnesol and its derivatives. Bioorg. Med. Chem. Lett. 2018, 28, 642–645. [Google Scholar] [CrossRef]
- Jabra-Rizk, M.A.; Meiller, T.F.; James, C.E.; Shirtliff, M.E. Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob. Agents Chemother. 2006, 50, 1463–1469. [Google Scholar] [CrossRef]
- Kong, E.F.; Tsui, C.; Kucharíková, S.; Van Dijck, P.; Jabra-Rizk, M.A. Modulation of Staphylococcus aureus Response to Antimicrobials by the Candida albicans Quorum Sensing Molecule Farnesol. Antimicrob. Agents Chemother. 2017, 61, e01573-17. [Google Scholar] [CrossRef]
- Gillum, A.M.; Tsay, E.Y.; Kirsch, D.R. Isolation of the Candida albicans gene for orotidine-5’-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 1984, 198, 179–182. [Google Scholar] [CrossRef]
- Vannuffel, P.; Heusterspreute, M.; Bouyer, M.; Vandercam, B.; Philippe, M.; Gala, J.L. Molecular characterization of femA from Staphylococcus hominis and Staphylococcus saprophyticus, and femA-based discrimination of staphylococcal species. Res. Microbiol. 1999, 150, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Černáková, L.; Dižová, S.; Gášková, D.; Jančíková, I.; Bujdáková, H. Impact of Farnesol as a Modulator of Efflux Pumps in a Fluconazole-Resistant Strain of Candida albicans. Microb. Drug Resist. 2019, 25, 805–812. [Google Scholar] [CrossRef]
- Kucharíková, S.; Tournu, H.; Lagrou, K.; Van Dijck, P.; Bujdáková, H. Detailed comparison of Candida albicans and Candida glabrata biofilms under different conditions and their susceptibility to caspofungin and anidulafungin. J. Med. Microbiol. 2011, 60, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- European Committee On Antimicrobial Susceptibility Testing (EUCAST, Version 12.0, 2022), Clinical Breakpoints—Breakpoints and Guidance. Available online: https://www.eucast.org/eucast_news/news_singleview/?tx_ttnews%5Btt_news%5D=459&cHash=160a5b91371e598957e10178fb3aa143 (accessed on 2 September 2022).
- Schoenrath, F.; Kursawe, L.; Nersesian, G.; Kikhney, J.; Schmidt, J.; Barthel, F.; Kaufmann, F.; Knierim, J.; Knosalla, C.; Hennig, F.; et al. Fluorescence In Situ Hybridization and Polymerase Chain Reaction to Detect Infections in Patients With Left Ventricular Assist Devices. ASAIO J. 2021, 67, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Trebesius, K.; Leitritz, L.; Adler, K.; Schubert, S.; Autenrieth, I.B.; Heesemann, J. Culture independent and rapid identification of bacterial pathogens in necrotising fasciitis and streptococcal toxic shock syndrome by fluorescence in situ hybridisation. Med. Microbiol. Immunol. 2000, 188, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef]
- Suetens, C.; Latour, K.; Kärki, T.; Ricchizzi, E.; Kinross, P.; Moro, M.L.; Jans, B.; Hopkins, S.; Hansen, S.; Lyytikäinen, O.; et al. Healthcare-Associated Infections Prevalence Study Group. Prevalence of healthcare-associated infections, estimated incidence and composite antimicrobial resistance index in acute care hospitals and long-term care facilities: Results from two European point prevalence surveys, 2016 to 2017. EuroSurveillance 2018, 23, 1800516. [Google Scholar] [CrossRef]
- Černáková, L.; Líšková, A.; Lengyelová, L.; Rodrigues, C.F. Prevalence and Antifungal Susceptibility Profile of Oral Candida spp. Isolates from a Hospital in Slovakia. Medicina 2022, 58, 576. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P.; Kathju, S.; Høiby, N.; Moser, C.; Costerton, J.W.; Moter, A.; Bjarnsholt, T. Towards diagnostic guidelines for biofilm-associated infections. FEMS Immunol. Med. Microbiol. 2012, 65, 127–145. [Google Scholar] [CrossRef]
- Dadi, N.C.T.; Radochová, B.; Vargová, J.; Bujdáková, H. Impact of Healthcare-Associated Infections Connected to Medical Devices-An Update. Microorganisms 2021, 9, 2332. [Google Scholar] [CrossRef] [PubMed]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum Sensing vs. Quorum Quenching: A Battle With No End in Sight; Springer: New Delhi, India, 2015; pp. 23–37. [Google Scholar]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Kateete, D.P.; Bwanga, F.; Seni, J.; Mayanja, R.; Kigozi, E.; Mujuni, B.; Ashaba, F.K.; Baluku, H.; Najjuka, C.F.; Källander, K.; et al. CA-MRSA and HA-MRSA coexist in community and hospital settings in Uganda. Antimicrob. Resist. Infect. Control 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Okuma, K.; Ma, X.X.; Yuzawa, H.; Hiramatsu, K. Insights on antibiotic resistance of Staphylococcus aureus from its whole genome: Genomic island SCC. Drug Resist. Updates 2003, 6, 41–52. [Google Scholar] [CrossRef]
- Kong, E.F.; Johnson, J.K.; Jabra-Rizk, M.A. Community-Associated Methicillin-Resistant Staphylococcus aureus: An Enemy amidst Us. PLoS Pathog. 2016, 12, e1005837. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Community-associated methicillin-resistant Staphylococcus aureus: Epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef]
- Pardo, L.; Machado, V.; Cuello, D.; Aguerrebere, P.; Seija, V.; Braga, V.; Varela, G. Macrolide-lincosamide-streptogramin B resistance phenotypes and their associated genotypes in Staphylococcus aureus isolates from a tertiary level public hospital of Uruguay. Rev. Argent. Microbiol. 2020, 52, 202–210. [Google Scholar] [CrossRef]
- Voss, A.; Loeffen, F.; Bakker, J.; Klaassen, C.; Wulf, M. Methicillin-resistant Staphylococcus aureus in Pig Farming. Emerg. Infect Dis. 2005, 11, 1965–1966. [Google Scholar] [CrossRef]
- Clarebout, G.; Nativelle, E.; Bozdogan, B.; Villers, C.; Leclercq, R. Bactericidal activity of quinupristin-dalfopristin against strains of Staphylococcus aureus with the MLS(B) phenotype of resistance according to the erm gene type. Int. J. Antimicrob. Agents 2004, 24, 444–449. [Google Scholar] [CrossRef]
- Thapa, D.; Pyakurel, S.; Thapa, S.; Lamsal, S.; Chaudhari, M.; Adhikari, N.; Shrestha, D. Staphylococcus aureus with inducible clindamycin resistance and methicillin resistance in a tertiary hospital in Nepal. Trop. Med. Health 2021, 49, 99. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Archer, G.L. Staphylococcus aureus: A well–armed pathogen. Clin. Infect. Dis. 1998, 26, 1179–1181. [Google Scholar] [CrossRef] [PubMed]
- Leshem, T.; Schnall, B.S.; Azrad, M.; Baum, M.; Rokney, A.; Peretz, A. Incidence of biofilm formation among MRSA and MSSA clinical isolates from hospitalized patients in Israel. J. Appl. Microbiol. 2022, 133, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Senobar Tahaei, S.A.; Stájer, A.; Barrak, I.; Ostorházi, E.; Szabó, D.; Gajdács, M. Correlation Between Biofilm-Formation and the Antibiotic Resistant Phenotype in Staphylococcus aureus Isolates: A Laboratory-Based Study in Hungary and a Review of the Literature. Infect. Drug Resist. 2021, 14, 1155–1168. [Google Scholar] [CrossRef]
- Ghasemian, A.; Najar Peerayeh, S.; Bakhshi, B.; Mirzaee, M. Comparison of Biofilm Formation between Methicillin-Resistant and Methicillin-Susceptible Isolates of Staphylococcus aureus. Iran Biomed. J. 2016, 20, 175–181. [Google Scholar] [CrossRef]
- Demuyser, L.; Jabra-Rizk, M.A.; Van Dijck, P. Microbial cell surface proteins and secreted metabolites involved in multispecies biofilms. Pathog. Dis. 2014, 70, 219–230. [Google Scholar] [CrossRef]
- Keller, L.; Surette, M.G. Communication in bacteria: An ecological and evolutionary perspective. Nat. Rev. Microbiol. 2006, 4, 249–258. [Google Scholar] [CrossRef]
- Kuroda, M.; Nagasaki, S.; Ohta, T. Sesquiterpene farnesol inhibits recycling of the C55 lipid carrier of the murein monomer precursor contributing to increased susceptibility to β-lactams in methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2007, 59, 425–432. [Google Scholar] [CrossRef]
- Kaneko, M.; Togashi, N.; Hamashima, H.; Hirohara, M.; Inoue, Y. Effect of farnesol on mevalonate pathway of Staphylococcus aureus. J. Antibiot. 2011, 64, 547–549. [Google Scholar] [CrossRef]
- Hada, T.; Shiraishi, A.; Furuse, S.; Inoue, Y.; Hamashima, H.; Matsumoto, Y.; Masuda, K.; Shiojima, K.; Shimada, J. Inhibitory effects of terpenes on the growth of Staphylococcus aureus. Nat. Med. 2003, 57, 64–67. [Google Scholar]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Hayacibara, M.F.; Schobel, B.D.; Cury, J.A.; Rosalen, P.L.; Park, Y.K.; Vacca-Smith, A.M.; Bowen, W.H. Inhibition of Streptococcus mutans biofilm accumulation and polysaccharide production by apigenin and tt-farnesol. J. Antimicrob. Chemother. 2003, 52, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Jabra-Rizk, M.A.; Scheper, M.A.; Leid, J.G.; Costerton, J.W.; Shirtliff, M.E. Microbial interactions and differential protein expression in Staphylococcus aureus–Candida albicans dual-species biofilms. FEMS Immunol. Med. Microbiol. 2010, 59, 493–503. [Google Scholar] [CrossRef]
- Décanis, N.; Tazi, N.; Correia, A.; Vilanova, M.; Rouabhia, M. Farnesol, a fungal quorum-sensing molecule triggers Candida albicans morphological changes by downregulating the expression of different secreted aspartyl proteinase genes. Open Microbiol. J. 2011, 5, 119–126. [Google Scholar] [CrossRef]
- Santos Ferreira, I.; Kikhney, J.; Kursawe, L.; Kasper, S.; Gonçalves, L.M.D.; Trampuz, A.; Moter, A.; Bettencourt, A.F.; Almeida, A.J. Encapsulation in polymeric microparticles improves daptomycin activity against mature Staphylococci biofilms-a thermal and imaging study. AAPS Pharm. Sci. Tech. 2018, 19, 1625–1636. [Google Scholar] [CrossRef]
- Sutrave, S.; Kikhney, J.; Schmidt, J.; Petrich, A.; Wiessner, A.; Kursawe, L.; Gebhardt, M.; Kertzscher, U.; Gabel, G.; Goubergrits, L.; et al. Effect of daptomycin and vancomycin on Staphylococcus epidermidis biofilms: An in vitro assessment using fluorescence in situ hybridization. PLoS ONE 2019, 14, e0221786. [Google Scholar] [CrossRef]
- Martineau, F.; Picard, F.J.; Lansac, N.; Ménard, C.; Roy, P.H.; Ouellette, M.; Bergeron, M.G. Correlation between the resistance genotype determined by multiplex PCR assays and the antibiotic susceptibility patterns of Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2000, 44, 231–238. [Google Scholar] [CrossRef]
- Choi, S.M.; Kim, S.H.; Kim, H.J.; Lee, D.G.; Choi, J.H.; Yoo, J.H.; Kang, J.H.; Shin, W.S.; Kang, M.W. Multiplex PCR for the detection of genes encoding aminoglycoside modifying enzymes and methicillin resistance among Staphylococcus species. J. Korean Med. Sci. 2003, 18, 631–636. [Google Scholar] [CrossRef]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef]
- Ojo, K.K.; Striplin, M.J.; Ulep, C.C.; Close, N.S.; Zittle, J.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M.C. Staphylococcus efflux msr(A) gene characterized in Streptococcus, Enterococcus, Corynebacterium, and Pseudomonas isolates. Antimicrob. Agents Chemother. 2006, 50, 1089–1091. [Google Scholar] [CrossRef]
- Rossato, A.M.; Primon-Barros, M.; Rocha, L.D.L.; Reiter, K.C.; Dias, C.A.G.; d’Azevedo, P.A. Resistance profile to antimicrobials agents in methicillin-resistant Staphylococcus aureus isolated from hospitals in South Brazil between 2014–2019. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200431. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Van de Walle, K.; Wickes, B.L.; Lopez-Ribot, J.L. Standardized method for in vitro antifungal susceptibility testing of Candida albicans. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M.A. Modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Dadi, N.C.T.; Bujdák, J.; Medvecká, V.; Pálková, H.; Barlog, M.; Bujdáková, H. Surface Characterization and Anti-Biofilm Effectiveness of Hybrid Films of Polyurethane Functionalized with Saponite and Phloxine B. Materials 2021, 14, 7583. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Group | Antibiotic/Dose | Strain of S. aureus | |||
|---|---|---|---|---|---|
| MSSA1 | MSSA2 | MRSA1 | MRSA2 | ||
| Penicillins | Oxacillin 10 µg | S | S | R | R |
| Ampicillin 10 µg | S | R | R | R | |
| Cephalosporins | Cefotaxime 30 µg | S | S | R | R |
| Cefoxitin 30 µg | S | S | R | R | |
| Carbapenems | Ertapenem 10 µg | S | S | R | R |
| Glycopeptides | Vancomycin 30 µg | S | S | S | S |
| Fluoroquinolones | Ciprofloxacin 5 µg | S | S | R | R |
| Ofloxacin 5 µg | S | R | R | R | |
| Aminoglycosides | Gentamicin 10 µg | S | S | S | S |
| Tobramycin 10 µg | S | S | R | R | |
| Macrolides | Erythromycin 15 µg | S | R | R | R |
| Lincosamides | Clindamycin 2 µg | S | R | R | R |
| Streptogramines | Quinupristin/dalphopristin 15 µg | S | S | S | S |
| Tetracyclines | Tetracycline 30 µg | S | S | S | S |
| Antibiotic Group | Resistance Signatures | S. aureus Isolates | |||
|---|---|---|---|---|---|
| MSSA1 | MSSA2 | MRSA1 | MRSA2 | ||
| Beta-lactams | mecA | − | − | + | + |
| Beta-lactamases production | − | + | + | + | |
| Fluoroquinolones | norA | + | + | + | + |
| norB | + | + | + | + | |
| norC | + | + | + | + | |
| Aminoglycosides | ant(4′)-Ia | − | − | + | + |
| aph(3′)-III | − | − | − | − | |
| aac(6′)-aph(2″) | − | − | − | − | |
| Macrolides, lincosamides and streptogramin B | ermA | − | − | + | + |
| ermB | − | − | − | − | |
| ermC | − | − | − | − | |
| msrA | − | − | − | − | |
| msrB | − | − | − | − | |
| Microbial Strains | OD570 | SD | Biofilm Intensity |
|---|---|---|---|
| MSSA1 | 0.324 | 0.034 | moderate |
| MSSA2 | 0.337 | 0.012 | moderate |
| MRSA1 | 0.228 | 0.01 | moderate |
| MRSA2 | 0.291 | 0.013 | moderate |
| C. albicans | 0.363 | 0.03 | moderate |
| C. albicans-MSSA1 | 0.413 | 0.093 | strong |
| C. albinans-MSSA2 | 0.476 | 0.036 | strong |
| C. albicans-MRSA1 | 0.34 | 0.03 | moderate |
| C. albicans-MRSA2 | 0.376 | 0.094 | moderate |
| MSSA1 | MSSA2 | MRSA1 | MRSA2 | ||
|---|---|---|---|---|---|
| MIC (mg/mL) | |||||
| OXA | Control | 0.38 | 0.75 | 48 | 32 |
| 150 µM FAR | 0.19 | 0.5 | 24 | 12 | |
| 300 µM FAR | 0.19 | 0.38 | 4 | 6 | |
| FOX | Control MIC | 2 | 3 | 256 | 256 |
| 150 µM FAR | 1.5 | 2 | 96 | 256 | |
| 300 µM FAR | 1 | 2 | 16 | 2 | |
| K | Control MIC | 1.5 | 3 | 256 | 256 |
| 150 µM FAR | 1 | 3 | 96 | 256 | |
| 300 µM FAR | 0.5 | 0.75 | 24 | 48 | |
| CIP | Control MIC | 0.25 | 0.19 | 32 | 32 |
| 150 µM FAR | 0.094 | 0.094 | 32 | 32 | |
| 300 µM FAR | 0.012 | 0.032 | 32 | 32 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaálová-Radochová, B.; Kendra, S.; Jordao, L.; Kursawe, L.; Kikhney, J.; Moter, A.; Bujdáková, H. Effect of Quorum Sensing Molecule Farnesol on Mixed Biofilms of Candida albicans and Staphylococcus aureus. Antibiotics 2023, 12, 441. https://doi.org/10.3390/antibiotics12030441
Gaálová-Radochová B, Kendra S, Jordao L, Kursawe L, Kikhney J, Moter A, Bujdáková H. Effect of Quorum Sensing Molecule Farnesol on Mixed Biofilms of Candida albicans and Staphylococcus aureus. Antibiotics. 2023; 12(3):441. https://doi.org/10.3390/antibiotics12030441
Chicago/Turabian StyleGaálová-Radochová, Barbora, Samuel Kendra, Luisa Jordao, Laura Kursawe, Judith Kikhney, Annette Moter, and Helena Bujdáková. 2023. "Effect of Quorum Sensing Molecule Farnesol on Mixed Biofilms of Candida albicans and Staphylococcus aureus" Antibiotics 12, no. 3: 441. https://doi.org/10.3390/antibiotics12030441
APA StyleGaálová-Radochová, B., Kendra, S., Jordao, L., Kursawe, L., Kikhney, J., Moter, A., & Bujdáková, H. (2023). Effect of Quorum Sensing Molecule Farnesol on Mixed Biofilms of Candida albicans and Staphylococcus aureus. Antibiotics, 12(3), 441. https://doi.org/10.3390/antibiotics12030441