
Exposure of Paracoccidioides brasiliensis to Mebendazole Leads to Inhibition of Fungal Energy Production

 , ,
, ,  , , and
, , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
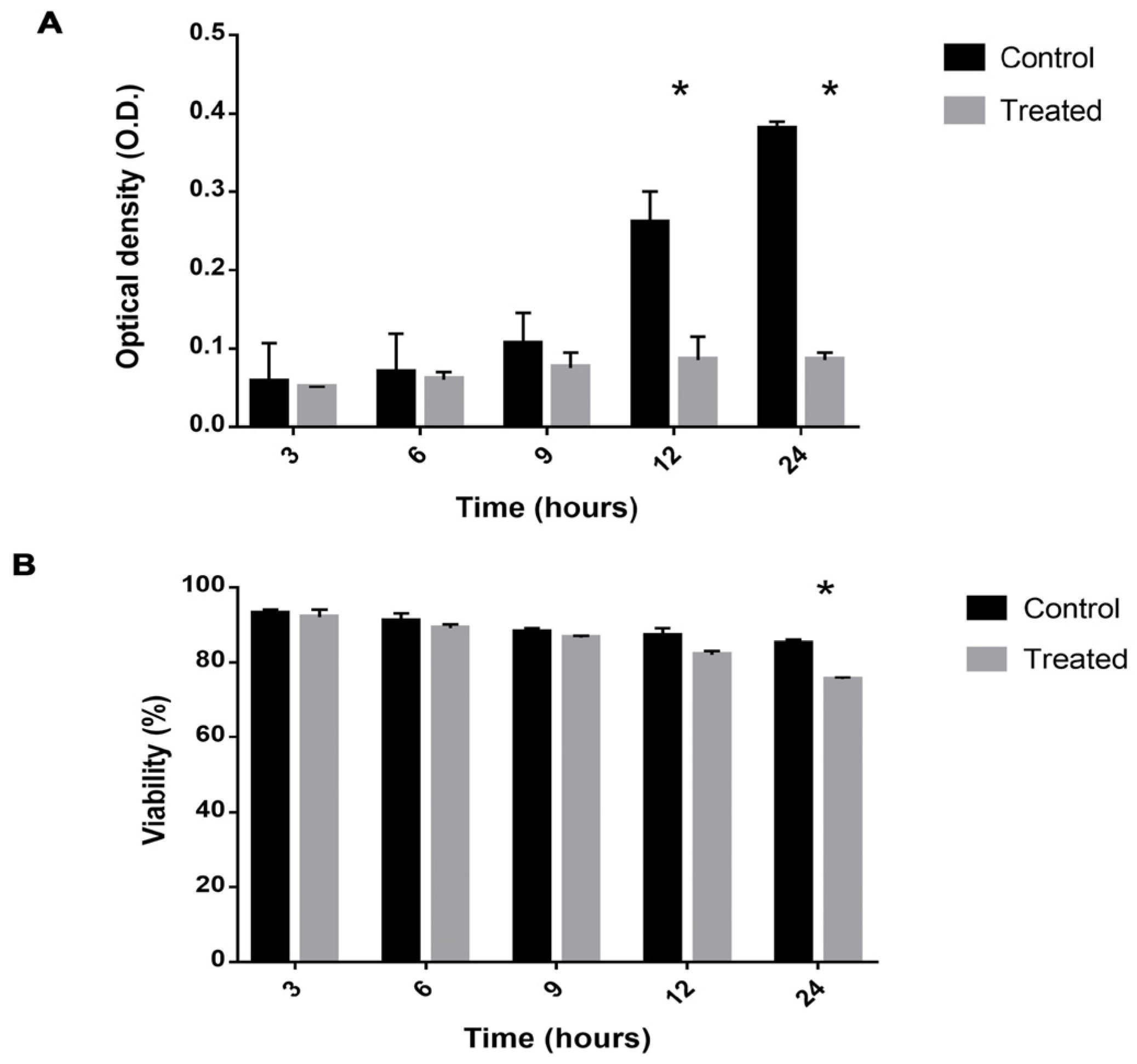
2.1. Viability Is Decreased and Growth Reduced in Cells Exposed to Mebendazole
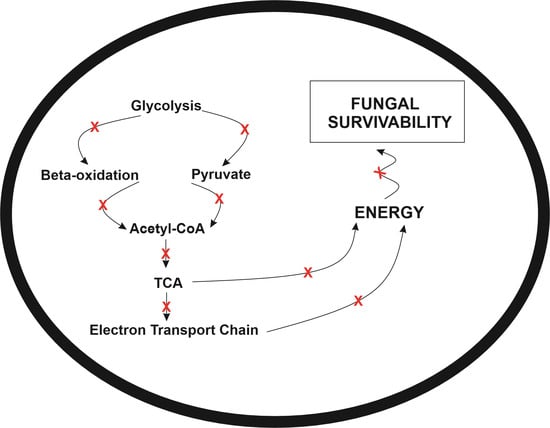
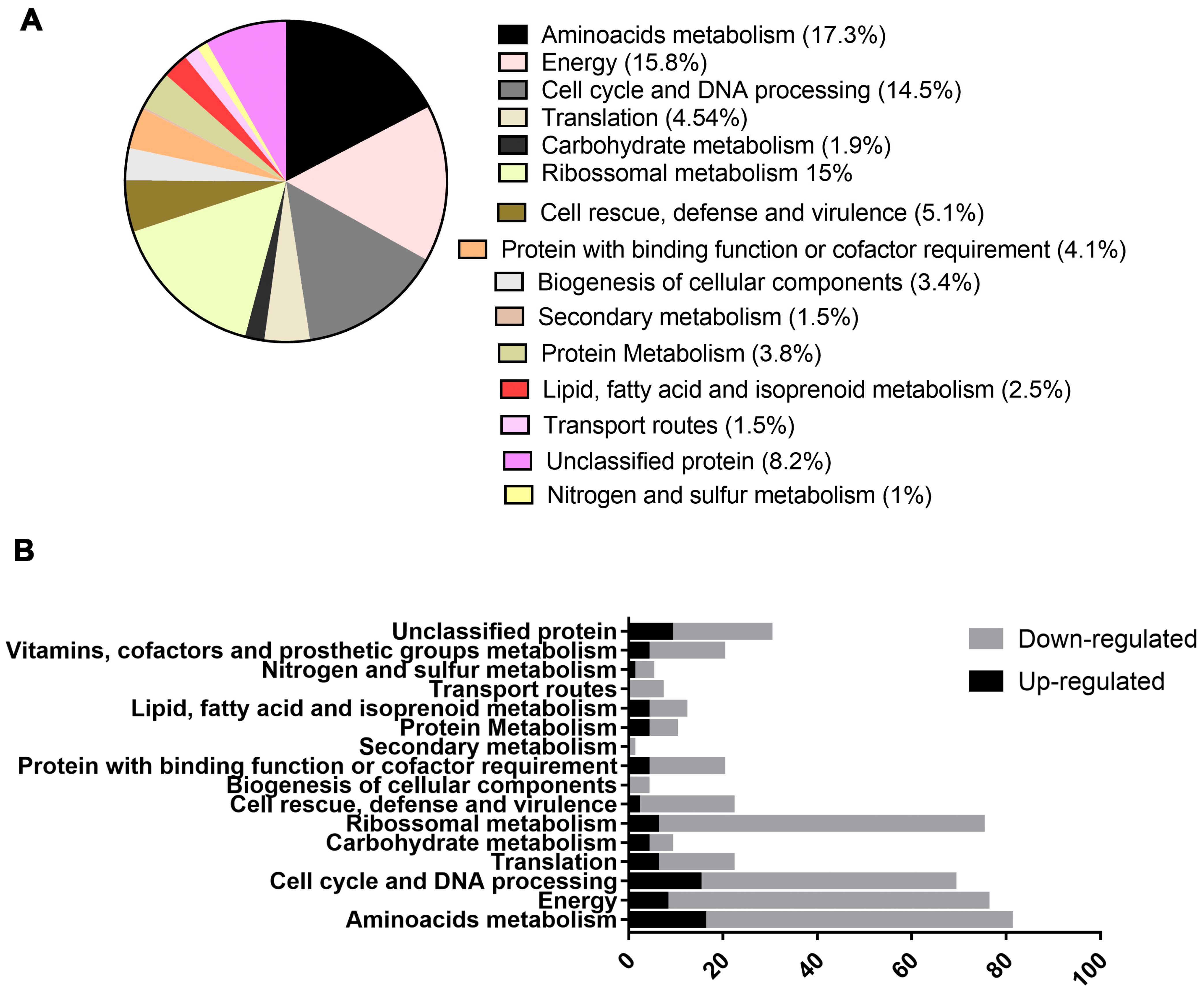
2.2. Exposure to Mebendazole Alters the Energy Metabolism of Cells
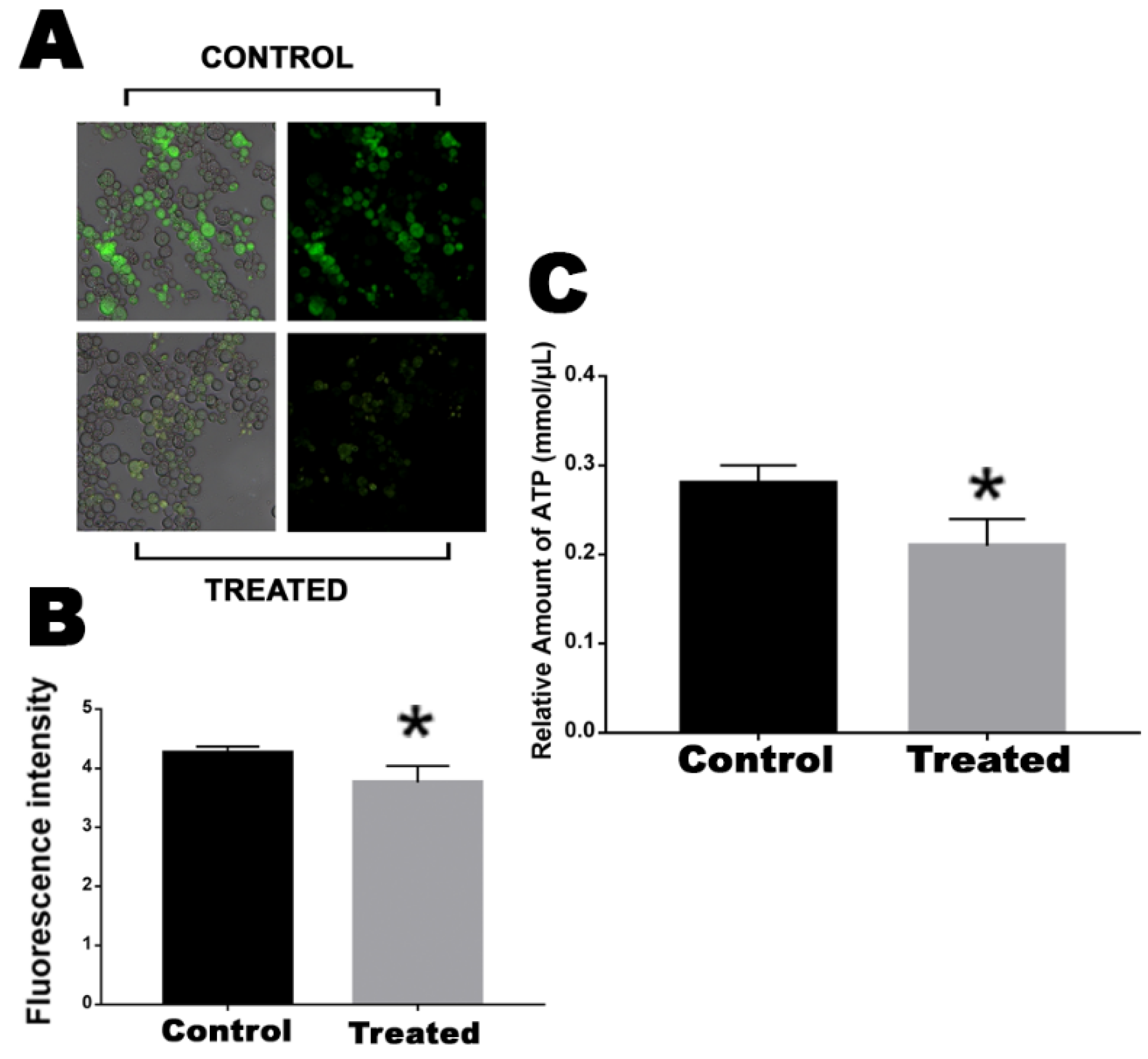
2.3. Mitochondrial Activity and ATP Production Is Decreased by Exposure to the Mebendazole
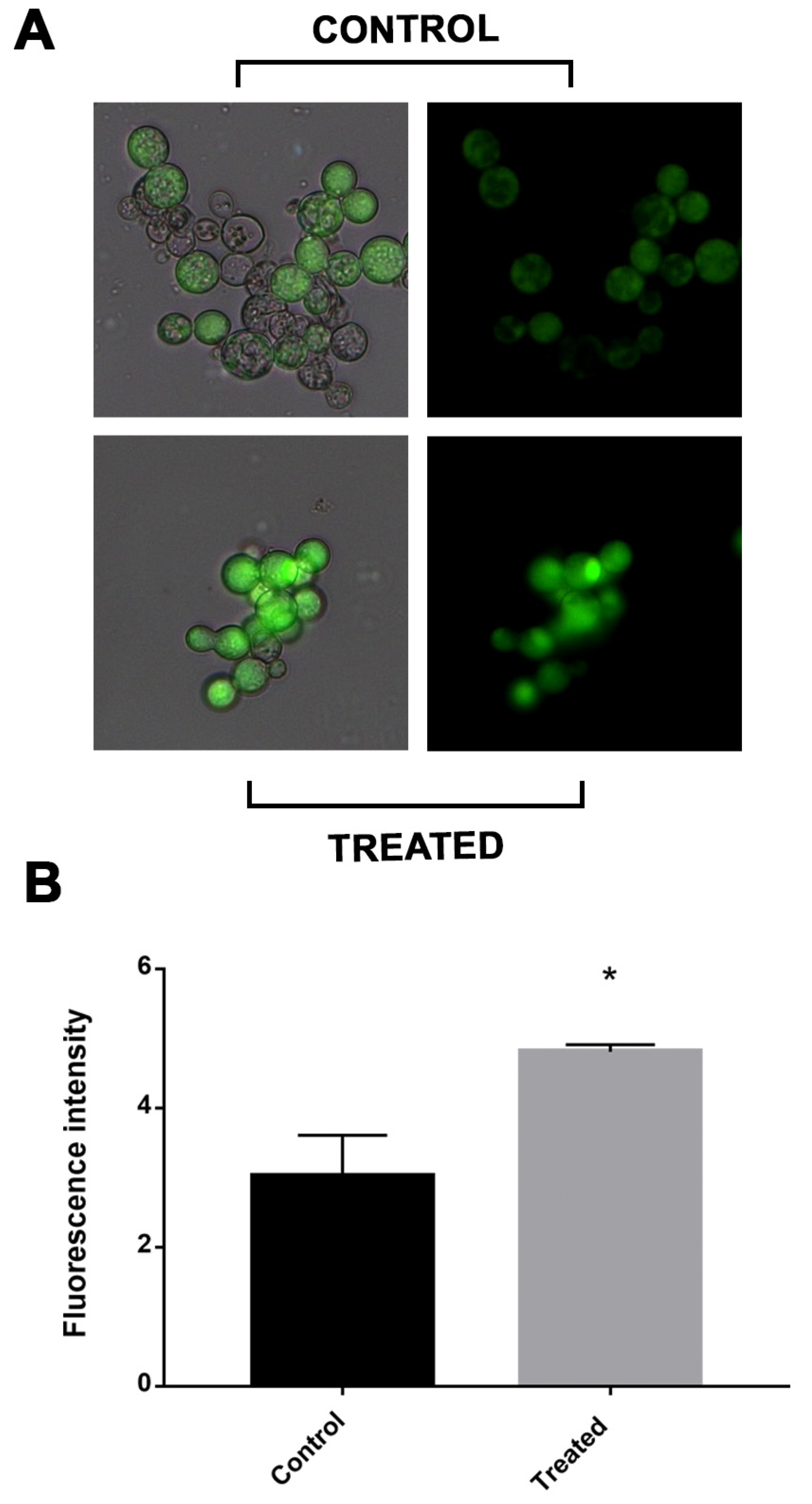
2.4. Oxidative Stress in Cells Is Increased after Incubation with Mebendazole
2.5. SOD and Cytochrome c Oxidase Enzymatic Activities Are Affected by Exposure to Mebendazole
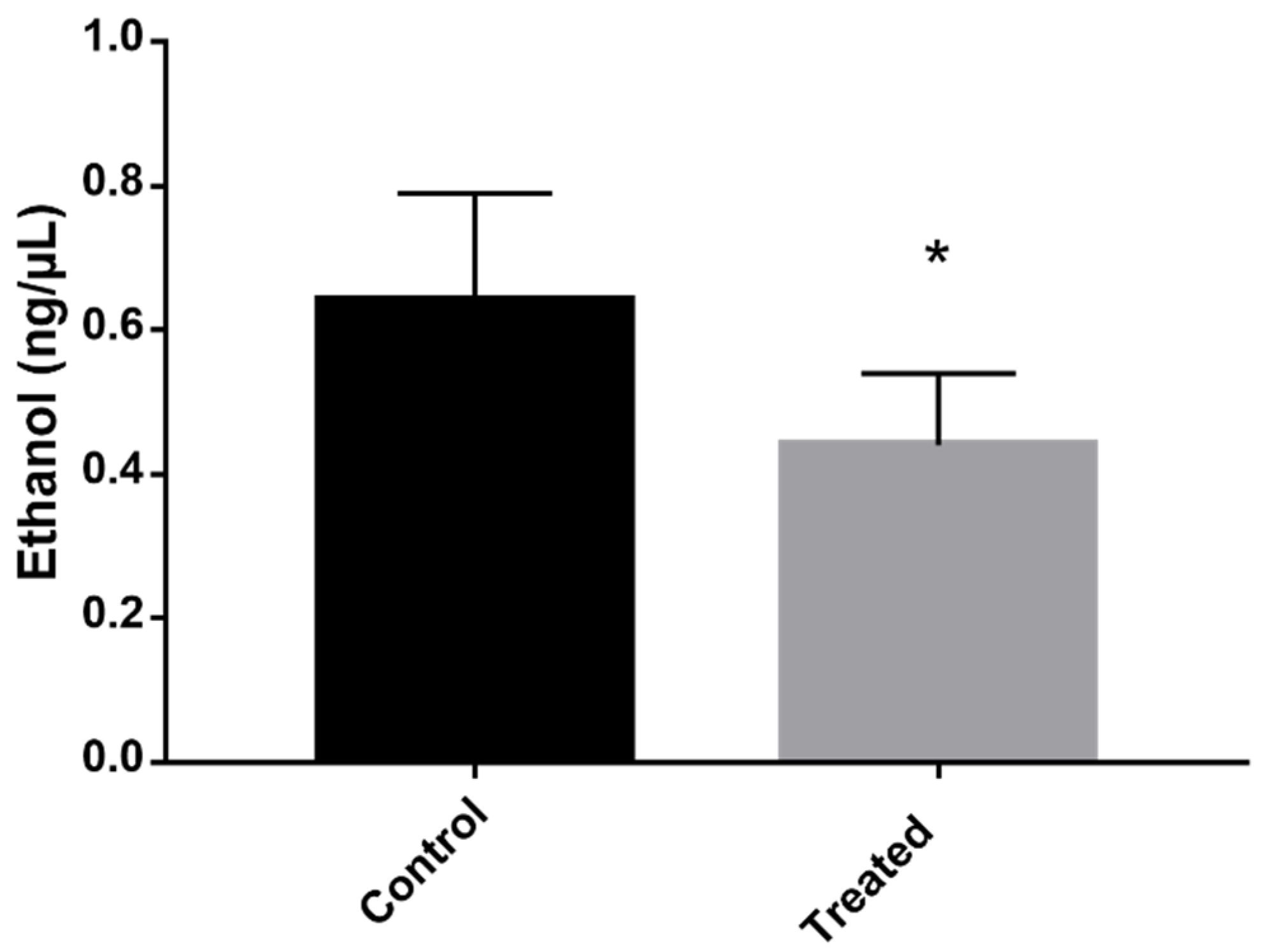
2.6. Energy Production by Fermentation Is Compromised after Exposure to the Mebendazole
3. Discussion
4. Materials and Methods
4.1. Microorganism and Culture Conditions
4.2. Viability and Growth Curve
4.3. Protein Extraction
4.4. Proteins Enzymatic Digestion
4.5. Liquid Chromatography and Mass Spectrometry
4.6. Mitochondrial Activity
4.7. ATP Quantification
4.8. Reactive Oxygen Species Quantification
4.9. Superoxide Dismutase Activity
4.10. Ethanol Quantification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santos, L.A.; Grisolia, J.C.; Burger, E.; de Araujo Paula, F.B.; Dias, A.L.T.; Malaquias, L.C.C. Virulence Factors of Paracoccidioides Brasiliensis as Therapeutic Targets: A Review. Antonie Van Leeuwenhoek 2020, 113, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S.S. Paracoccidioides Species Complex: Ecology, Phylogeny, Sexual Reproduction, and Virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Shikanai-Yasuda, M.A.; Soares Felipe, M.S. Paracoccidioidomycosis: Eco-Epidemiology, Taxonomy and Clinical and Therapeutic Issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian Guidelines for the Clinical Management of Paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Do Carmo Silva, L.; de Oliveira, A.A.; de Souza, D.R.; Barbosa, K.L.B.; Freitas e Silva, K.S.; Carvalho Júnior, M.A.B.; Rocha, O.B.; Lima, R.M.; Santos, T.G.; Soares, C.M.d.A.; et al. Overview of Antifungal Drugs against Paracoccidioidomycosis: How Do We Start, Where Are We, and Where Are We Going? J. Fungi 2020, 6, 300. [Google Scholar] [CrossRef]
- Guerini, A.E.; Triggiani, L.; Maddalo, M.; Bonù, M.L.; Frassine, F.; Baiguini, A.; Alghisi, A.; Tomasini, D.; Borghetti, P.; Pasinetti, N.; et al. Mebendazole as a Candidate for Drug Repurposing in Oncology: An Extensive Review of Current Literature. Cancers 2019, 11, 1284. [Google Scholar] [CrossRef] [PubMed]
- Joffe, L.S.; Schneider, R.; Lopes, W.; Azevedo, R.; Staats, C.C.; Kmetzsch, L.; Schrank, A.; Del Poeta, M.; Vainstein, M.H.; Rodrigues, M.L. The Anti-Helminthic Compound Mebendazole Has Multiple Antifungal Effects against Cryptococcus Neoformans. Front. Microbiol. 2017, 8, 535. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.-Y.; Jung, B.-K.; Hong, S.-J. Albendazole and Mebendazole as Anti-Parasitic and Anti-Cancer Agents: An Update. Korean J. Parasitol. 2021, 59, 189–225. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; Neves, B.J.; Silva, L.d.C.; Soares, C.M.d.A.; Andrade, C.H.; Pereira, M. Drug Repurposing for Paracoccidioidomycosis Through a Computational Chemogenomics Framework. Front. Microbiol. 2019, 10, 1301. [Google Scholar] [CrossRef]
- MacDonald, L.M.; Armson, A.; Thompson, A.R.C.; Reynoldson, J.A. Characterisation of Benzimidazole Binding with Recombinant Tubulin from Giardia Duodenalis, Encephalitozoon Intestinalis, and Cryptosporidium Parvum. Mol. Biochem. Parasitol. 2004, 138, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.C.; Bartlett, M.S.; Edlind, T.D. In Vitro Susceptibility of the Opportunistic Fungus Cryptococcus Neoformans to Anthelmintic Benzimidazoles. Antimicrob. Agents Chemother. 1994, 38, 378–380. [Google Scholar] [CrossRef]
- Rocha, O.B.; Freitas, E.; Silva, K.S.; de Carvalho Junior, M.A.B.; Moraes, D.; Alonso, A.; Alonso, L.; do Carmo Silva, L.; Soares, C.M.A.; Pereira, M. Proteomic Alterations in Paracoccidioides Brasiliensis Caused by Exposure to Curcumin. J. Proteom. 2022, 266, 104683. [Google Scholar] [CrossRef]
- Prado, R.S.; Bailão, A.M.; Silva, L.C.; de Oliveira, C.M.A.; Marques, M.F.; Silva, L.P.; Silveira-Lacerda, E.P.; Lima, A.P.; Soares, C.M.; Pereira, M. Proteomic Profile Response of Paracoccidioides Lutzii to the Antifungal Argentilactone. Front. Microbiol. 2015, 6, 616. [Google Scholar] [CrossRef]
- E Silva, K.S.; da S Neto, B.R.; Zambuzzi-Carvalho, P.F.; de Oliveira, C.M.; Pires, L.B.; Kato, L.; Bailão, A.M.; Parente-Rocha, J.A.; Hernández, O.; Ochoa, J.G.; et al. Response of Paracoccidioides Lutzii to the Antifungal Camphene Thiosemicarbazide Determined by Proteomic Analysis. Future Microbiol. 2018, 13, 1473–1496. [Google Scholar] [CrossRef]
- Lacerda Pigosso, L.; Baeza, L.C.; Vieira Tomazett, M.; Batista Rodrigues Faleiro, M.; Brianezi Dignani de Moura, V.M.; Melo Bailão, A.; Borges, C.L.; Alves Parente Rocha, J.; Rocha Fernandes, G.; Gauthier, G.M.; et al. Paracoccidioides Brasiliensis Presents Metabolic Reprogramming and Secretes a Serine Proteinase during Murine Infection. Virulence 2017, 8, 1417–1434. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Lima, P.; Casaletti, L.; Bailão, A.M.; de Vasconcelos, A.T.R.; da Rocha Fernandes, G.; de Almeida Soares, C.M. Transcriptional and Proteomic Responses to Carbon Starvation in Paracoccidioides. PLoS Negl. Trop. Dis. 2014, 8, e2855. [Google Scholar] [CrossRef]
- Delgado, J.; Owens, R.A.; Doyle, S.; Asensio, M.A.; Núñez, F. Impact of the Antifungal Protein PgAFP from Penicillium Chrysogenum on the Protein Profile in Aspergillus Flavus. Appl. Microbiol. Biotechnol. 2015, 99, 8701–8715. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, V.P.; Harsch, M.; Lampen, J.O.Y. Action of the Polyene Antibiotics Filipin, Nystatin and n-Acetylcandidin on the Yeast Cell Membrane. Microbiology 1964, 35, 249–259. [Google Scholar] [CrossRef]
- Katragkou, A.; Alexander, E.L.; Eoh, H.; Raheem, S.K.; Roilides, E.; Walsh, T.J. Effects of Fluconazole on the Metabolomic Profile of Candida Albicans. J. Antimicrob. Chemother. 2016, 71, 635–640. [Google Scholar] [CrossRef]
- Askew, C.; Sellam, A.; Epp, E.; Hogues, H.; Mullick, A.; Nantel, A.; Whiteway, M. Transcriptional Regulation of Carbohydrate Metabolism in the Human Pathogen Candida Albicans. PLoS Pathog. 2009, 5, e1000612. [Google Scholar] [CrossRef]
- Lima, P.d.S.; Chung, D.; Bailão, A.M.; Cramer, R.A.; Soares, C.M.d.A. Characterization of the Paracoccidioides Hypoxia Response Reveals New Insights into Pathogenesis Mechanisms of This Important Human Pathogenic Fungus. PLoS Negl. Trop. Dis. 2015, 9, e0004282. [Google Scholar] [CrossRef] [PubMed]
- Parente, A.F.A.; de Rezende, T.C.V.; de Castro, K.P.; Bailão, A.M.; Parente, J.A.; Borges, C.L.; Silva, L.P.; Soares, C.M.d.A. A Proteomic View of the Response of Paracoccidioides Yeast Cells to Zinc Deprivation. Fungal Biol. 2013, 117, 399–410. [Google Scholar] [CrossRef] [PubMed]
- De Arruda Grossklaus, D.; Bailão, A.M.; Vieira Rezende, T.C.; Borges, C.L.; de Oliveira, M.A.P.; Parente, J.A.; de Almeida Soares, C.M. Response to Oxidative Stress in Paracoccidioides Yeast Cells as Determined by Proteomic Analysis. Microbes Infect. 2013, 15, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, J.; Kong, W.; Zhao, G.; Yang, M. Mechanisms of Antifungal and Anti-Aflatoxigenic Properties of Essential Oil Derived from Turmeric (Curcuma longa L.) on Aspergillus Flavus. Food Chem. 2017, 220, 1–8. [Google Scholar] [CrossRef]
- Chambergo, F.S.; Bonaccorsi, E.D.; Ferreira, A.J.S.; Ramos, A.S.P.; Ferreira Júnior, J.R.; Abrahão-Neto, J.; Farah, J.P.S.; El-Dorry, H. Elucidation of the Metabolic Fate of Glucose in the Filamentous Fungus Trichoderma Reesei Using Expressed Sequence Tag (EST) Analysis and CDNA Microarrays. J. Biol. Chem. 2002, 277, 13983–13988. [Google Scholar] [CrossRef]
- Maeda, H.; Sano, M.; Maruyama, Y.; Tanno, T.; Akao, T.; Totsuka, Y.; Endo, M.; Sakurada, R.; Yamagata, Y.; Machida, M.; et al. Transcriptional Analysis of Genes for Energy Catabolism and Hydrolytic Enzymes in the Filamentous Fungus Aspergillus Oryzae Using CDNA Microarrays and Expressed Sequence Tags. Appl. Microbiol. Biotechnol. 2004, 65, 74–83. [Google Scholar] [CrossRef]
- Johnston, M.; Feasting, Fasting and Fermenting. Glucose Sensing in Yeast and Other Cells. Trends Genet. 1999, 15, 29–33. [Google Scholar] [CrossRef]
- Li, D.; Calderone, R. Exploiting Mitochondria as Targets for the Development of New Antifungals. Virulence 2017, 8, 159–168. [Google Scholar] [CrossRef]
- OuYang, Q.; Tao, N.; Zhang, M. A Damaged Oxidative Phosphorylation Mechanism Is Involved in the Antifungal Activity of Citral against Penicillium Digitatum. Front. Microbiol. 2018, 9, 239. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Zhang, Y.; Zhang, Y.; Zhang, Z.; Tang, C.; Weng, L.; Chen, X.; Zhang, G.; Zhang, H. The δ Subunit of F1Fo-ATP Synthase Is Required for Pathogenicity of Candida Albicans. Nat. Commun. 2021, 12, 6041. [Google Scholar] [CrossRef]
- Vestergaard, M.; Bald, D.; Ingmer, H. Targeting the ATP Synthase in Bacterial and Fungal Pathogens: Beyond Mycobacterium Tuberculosis. J. Glob. Antimicrob. Resist. 2022, 29, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Chaves, E.G.A.; Parente-Rocha, J.A.; Baeza, L.C.; Araújo, D.S.; Borges, C.L.; de Oliveira, M.A.P.; Soares, C.M.d.A. Proteomic Analysis of Paracoccidioides Brasiliensis During Infection of Alveolar Macrophages Primed or Not by Interferon-Gamma. Front. Microbiol. 2019, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Parente-Rocha, J.A.; Parente, A.F.A.; Baeza, L.C.; Bonfim, S.M.R.C.; Hernandez, O.; McEwen, J.G.; Bailão, A.M.; Taborda, C.P.; Borges, C.L.; de Almeida Soares, C.M. Macrophage Interaction with Paracoccidioides Brasiliensis Yeast Cells Modulates Fungal Metabolism and Generates a Response to Oxidative Stress. PLoS ONE 2015, 10, e0137619. [Google Scholar] [CrossRef] [PubMed]
- Castilho, D.G.; Navarro, M.V.; Chaves, A.F.A.; Xander, P.; Batista, W.L. Recovery of the Paracoccidioides Brasiliensis Virulence after Animal Passage Promotes Changes in the Antioxidant Repertoire of the Fungus. FEMS Yeast Res. 2018, 18, foy007. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, O.B.; e Silva, K.S.F.; Moraes, D.; Borges, C.L.; Soares, C.M.d.A.; Pereira, M. Exposure of Paracoccidioides brasiliensis to Mebendazole Leads to Inhibition of Fungal Energy Production. Antibiotics 2023, 12, 206. https://doi.org/10.3390/antibiotics12020206
Rocha OB, e Silva KSF, Moraes D, Borges CL, Soares CMdA, Pereira M. Exposure of Paracoccidioides brasiliensis to Mebendazole Leads to Inhibition of Fungal Energy Production. Antibiotics. 2023; 12(2):206. https://doi.org/10.3390/antibiotics12020206
Chicago/Turabian StyleRocha, Olivia Basso, Kleber Santiago Freitas e Silva, Dayane Moraes, Clayton Luiz Borges, Célia Maria de Almeida Soares, and Maristela Pereira. 2023. "Exposure of Paracoccidioides brasiliensis to Mebendazole Leads to Inhibition of Fungal Energy Production" Antibiotics 12, no. 2: 206. https://doi.org/10.3390/antibiotics12020206
APA StyleRocha, O. B., e Silva, K. S. F., Moraes, D., Borges, C. L., Soares, C. M. d. A., & Pereira, M. (2023). Exposure of Paracoccidioides brasiliensis to Mebendazole Leads to Inhibition of Fungal Energy Production. Antibiotics, 12(2), 206. https://doi.org/10.3390/antibiotics12020206