

Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
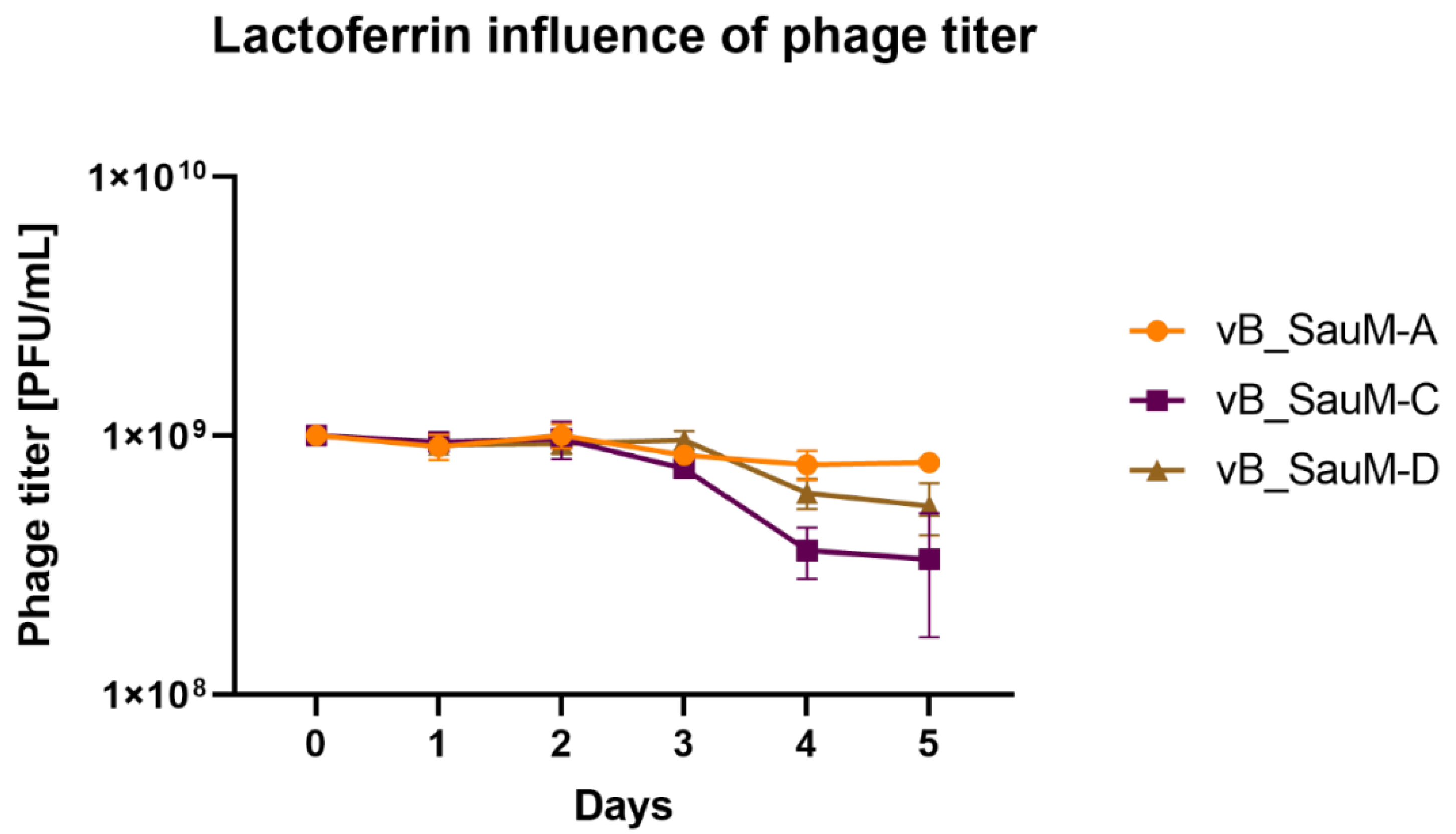
2.1. Lactoferrin Influence on Phage Activity
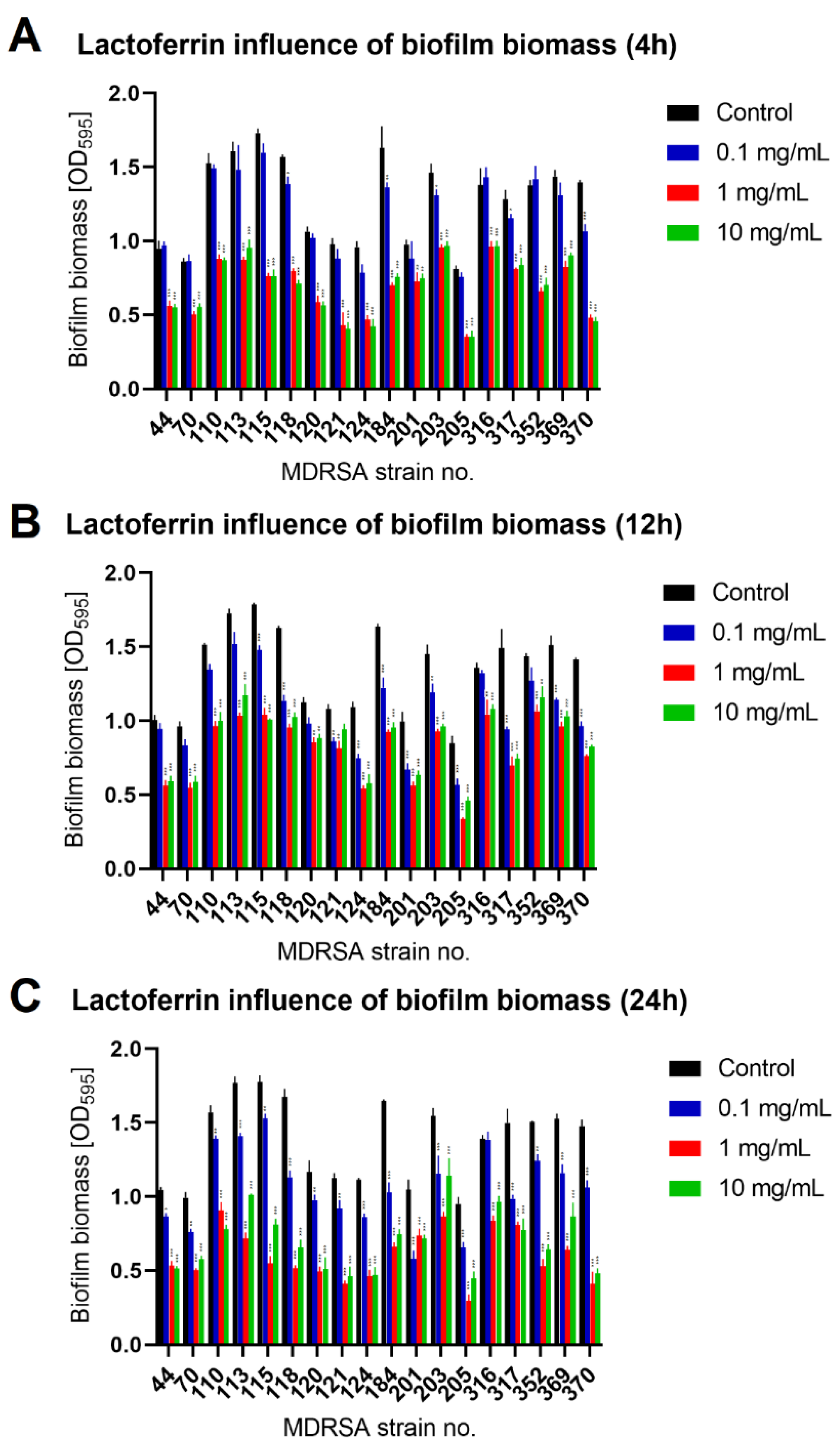
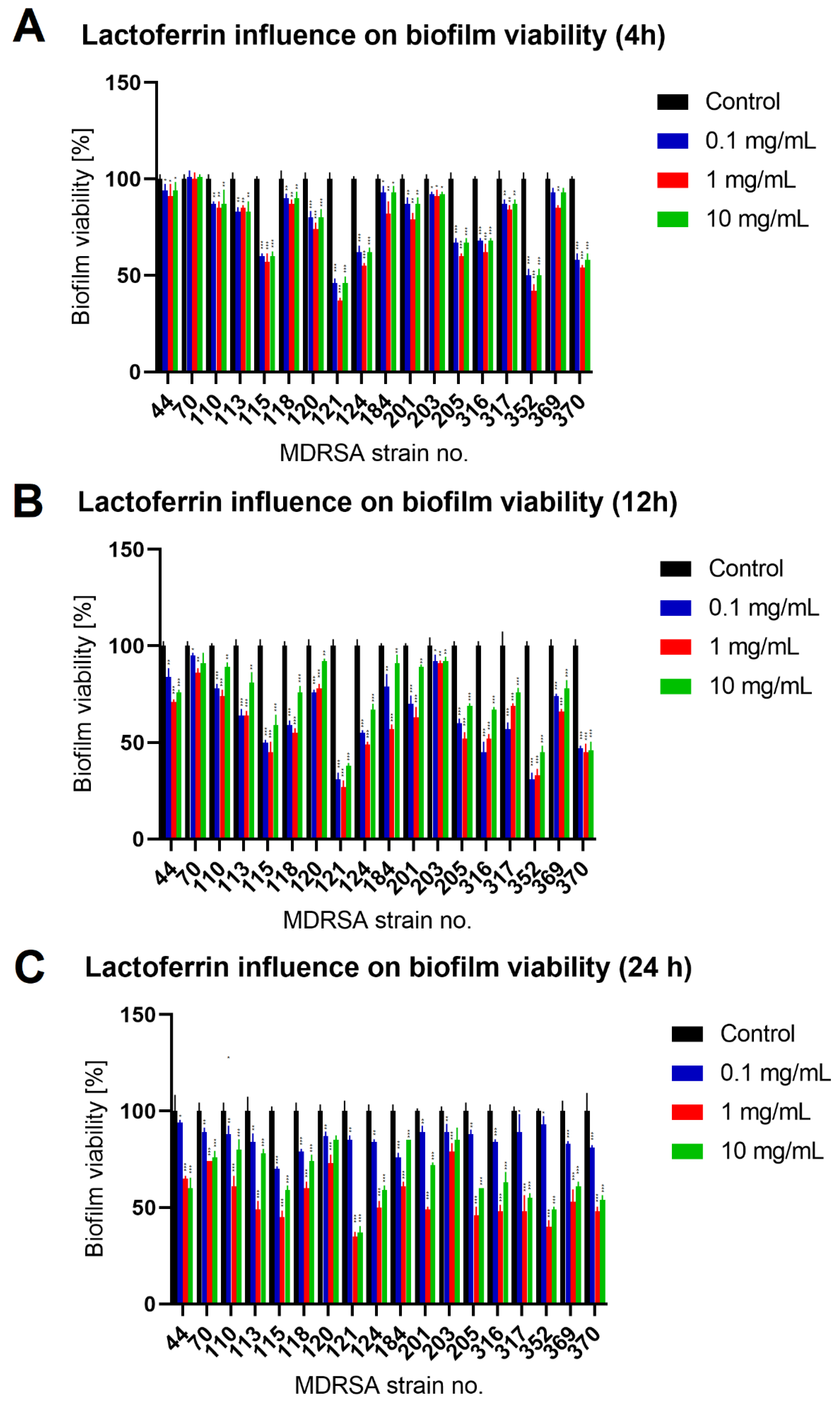
2.2. Lactoferrin Influence on Biofilm Formed by MDRSA Strains
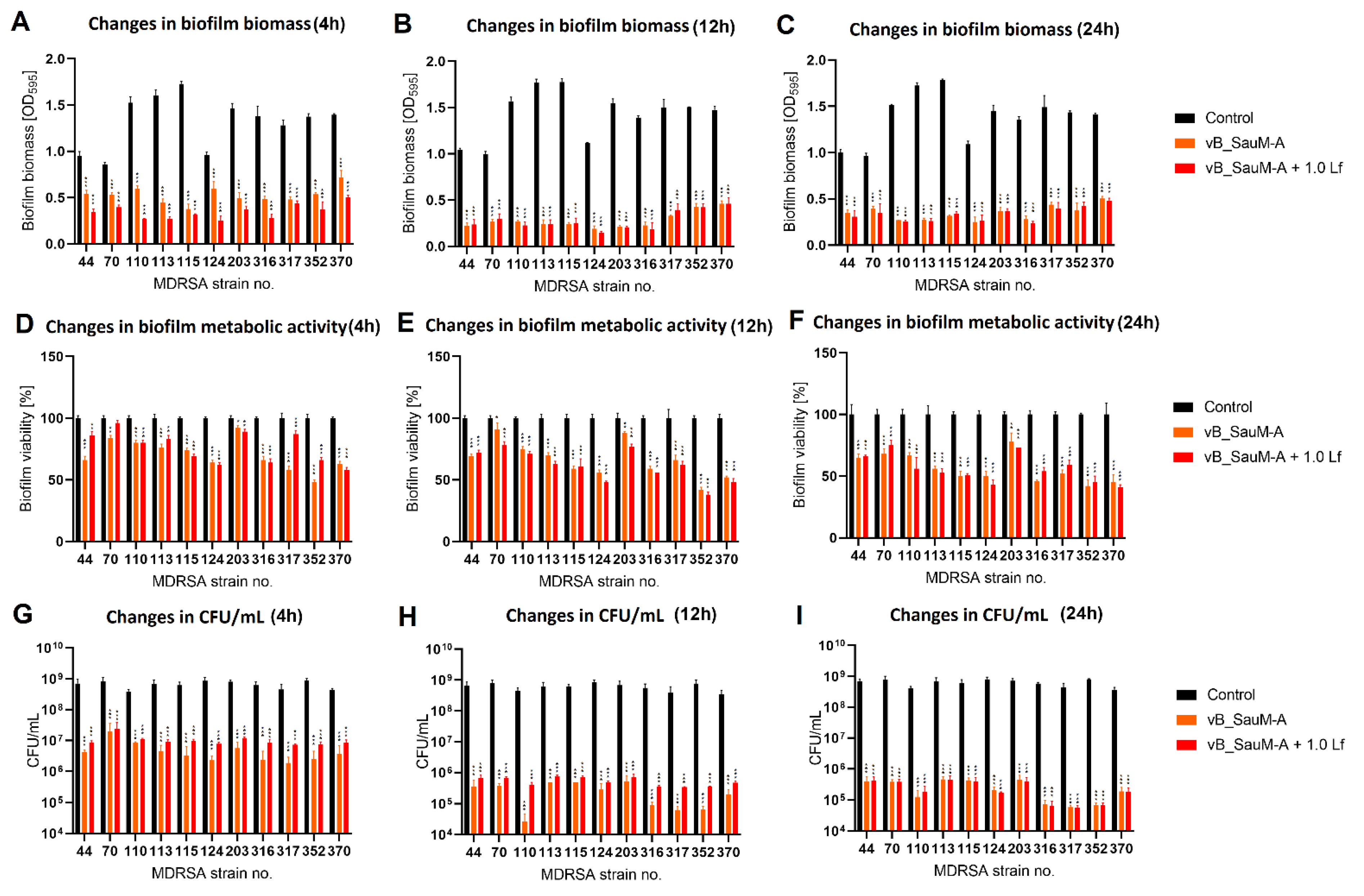
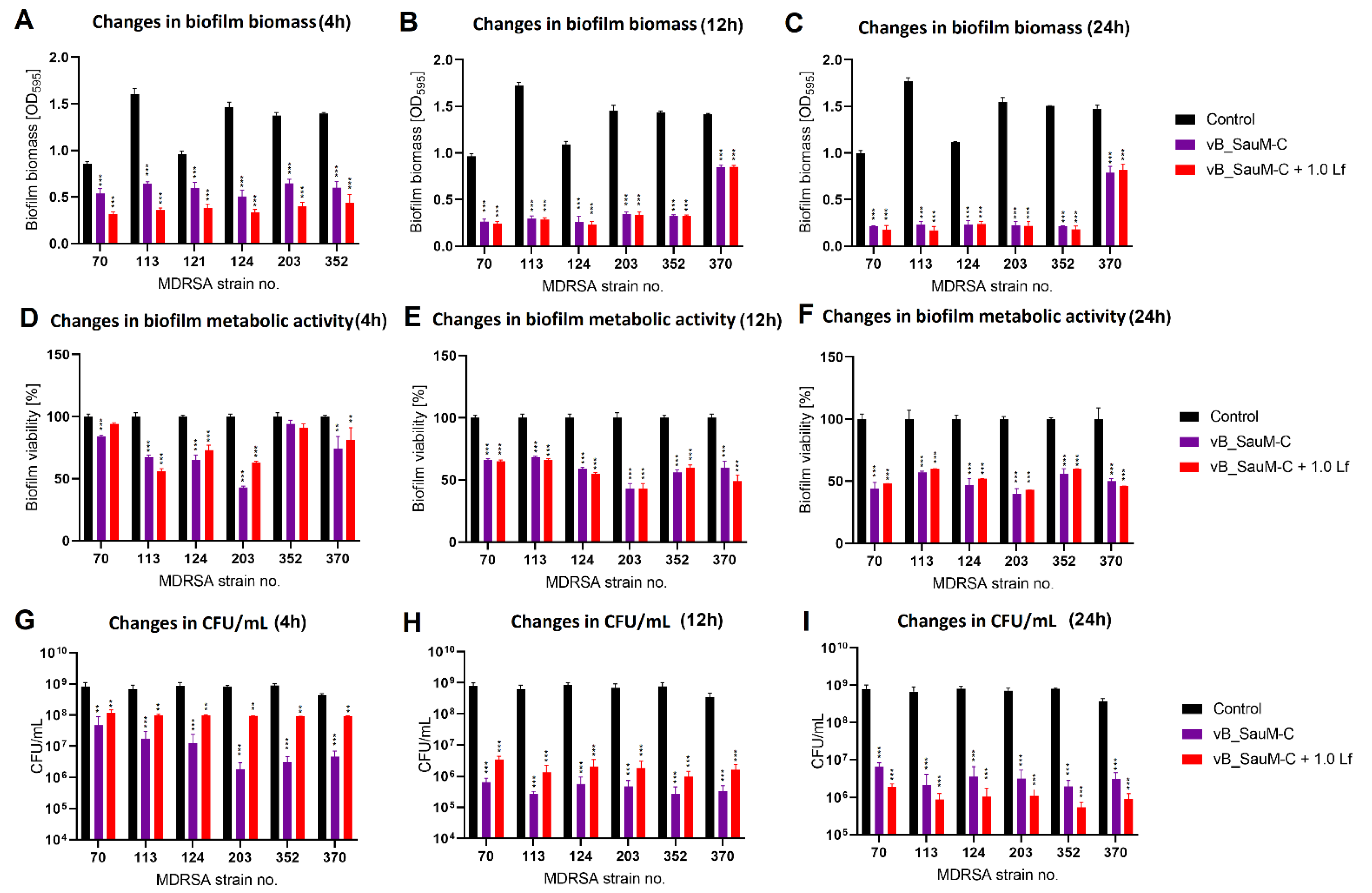
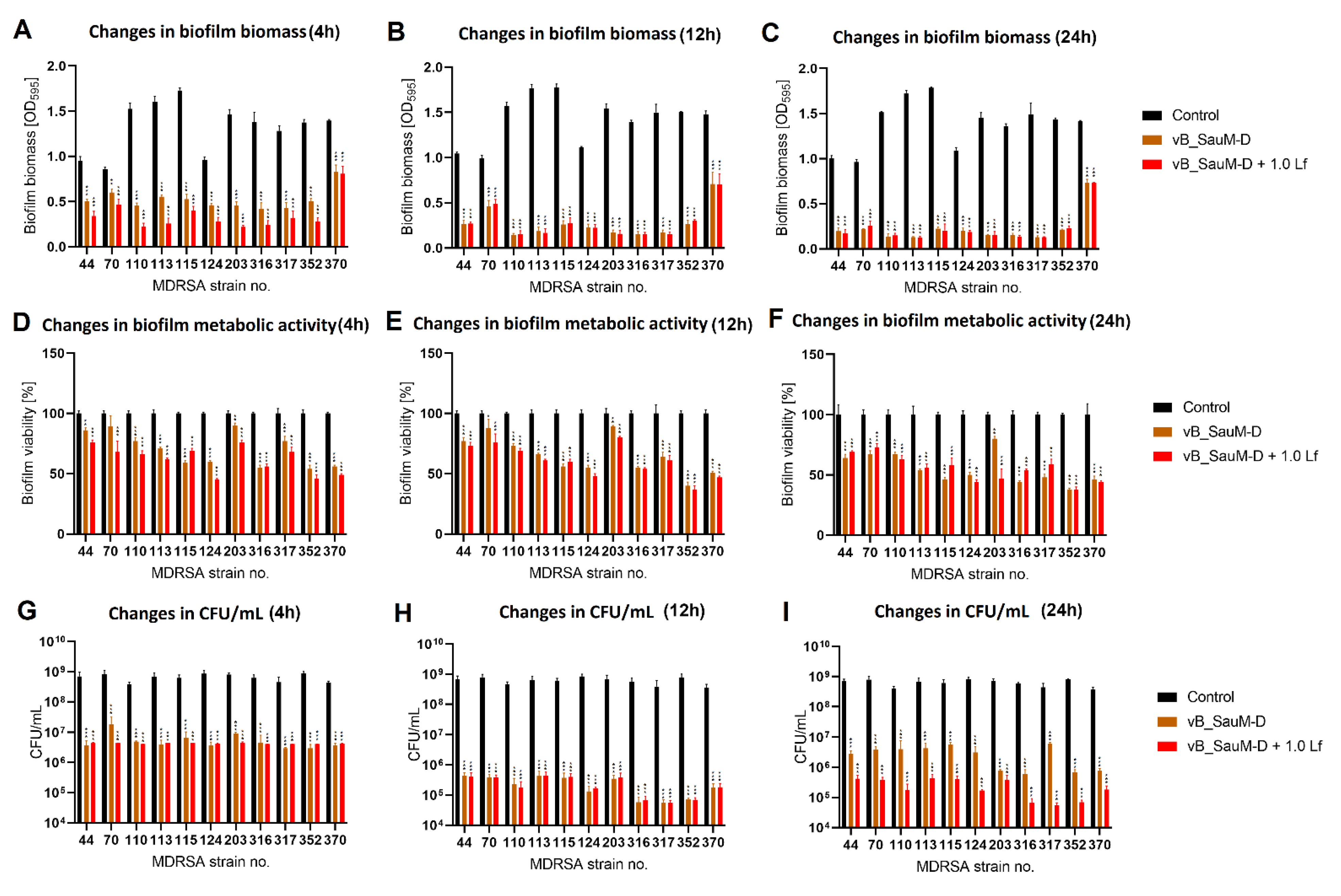
2.3. Biofilm Eradication by Phage–Lactoferrin Mixture
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Bacteriophages
4.3. Lactoferrin
4.4. Lactoferrin Influence on Phage Activity
4.5. Assessment of Biofilm Biomass Using Crystal Violet Staining
4.6. Assessment of Biofilm Metabolic Activity Using MTT
4.7. Enumeration of Cells in Biofilm Using CFU/mL Count
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, R.L.; Cabezudo, I.; Wenzel, R.P. Epidemiology of Nosocomial Infections Caused by Methicillin-Resistant Staphylococcus aureus. Ann. Intern. Med. 1982, 97, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.L. Staphylococcus aureus. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 379–380. ISBN 978-0-12-386455-0. [Google Scholar]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and Virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Pokhrel, B.; Mohapatra, T. Study of Nosocomial Isolates of Staphylococcus aureus with Special Reference to Methicillin Resistant S. Aureus in a Tertiary Care Hospital in Nepal. Nepal Med. Coll. J. NMCJ 2009, 11, 123–126. [Google Scholar] [PubMed]
- Stefani, S.; Chung, D.R.; Lindsay, J.A.; Friedrich, A.W.; Kearns, A.M.; Westh, H.; MacKenzie, F.M. Meticillin-Resistant Staphylococcus aureus (MRSA): Global Epidemiology and Harmonisation of Typing Methods. Int. J. Antimicrob. Agents 2012, 39, 273–282. [Google Scholar] [CrossRef]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus Biofilms: Recent Developments in Biofilm Dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef]
- Kaźmierczak, N.; Grygorcewicz, B.; Roszak, M.; Bochentyn, B.; Piechowicz, L. Comparative Assessment of Bacteriophage and Antibiotic Activity against Multidrug-Resistant Staphylococcus aureus Biofilms. Int. J. Mol. Sci. 2022, 23, 1274. [Google Scholar] [CrossRef]
- Kifelew, L.G.; Warner, M.S.; Morales, S.; Thomas, N.; Gordon, D.L.; Mitchell, J.G.; Speck, P.G. Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses 2020, 12, 559. [Google Scholar] [CrossRef]
- Chang, C.; Yu, X.; Guo, W.; Guo, C.; Guo, X.; Li, Q.; Zhu, Y. Bacteriophage-Mediated Control of Biofilm: A Promising New Dawn for the Future. Front. Microbiol. 2022, 13, 825828. [Google Scholar] [CrossRef]
- Moons, P.; Faster, D.; Aertsen, A. Lysogenic Conversion and Phage Resistance Development in Phage Exposed Escherichia coli Biofilms. Viruses 2013, 5, 150–161. [Google Scholar] [CrossRef]
- Simmons, E.L.; Bond, M.C.; Koskella, B.; Drescher, K.; Bucci, V.; Nadell, C.D. Biofilm Structure Promotes Coexistence of Phage-Resistant and Phage-Susceptible Bacteria. mSystems 2020, 5, e00877-19. [Google Scholar] [CrossRef] [PubMed]
- Ferriol-González, C.; Domingo-Calap, P. Phages for Biofilm Removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Manohar, P.; Madurantakam Royam, M.; Loh, B.; Bozdogan, B.; Nachimuthu, R.; Leptihn, S. Synergistic Effects of Phage–Antibiotic Combinations against Citrobacter Amalonaticus. ACS Infect. Dis. 2022, 8, 59–65. [Google Scholar] [CrossRef]
- Ammons, M.C.; Copié, V. Mini-Review: Lactoferrin: A Bioinspired, Anti-Biofilm Therapeutic. Biofouling 2013, 29, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Ramamourthy, G.; Vogel, H.J. Antibiofilm Activity of Lactoferrin-Derived Synthetic Peptides against Pseudomonas Aeruginosa PAO11. Biochem. Cell Biol. 2021, 99, 138–148. [Google Scholar] [CrossRef]
- Quintieri, L.; Caputo, L.; Monaci, L.; Cavalluzzi, M.M.; Denora, N. Lactoferrin-Derived Peptides as a Control Strategy against Skinborne Staphylococcal Biofilms. Biomedicines 2020, 8, 323. [Google Scholar] [CrossRef]
- Łubowska, N.; Grygorcewicz, B.; Kosznik-Kwaśnicka, K.; Zauszkiewicz-Pawlak, A.; Węgrzyn, A.; Dołęgowska, B.; Piechowicz, L. Characterization of the Three New Kayviruses and Their Lytic Activity against Multidrug-Resistant Staphylococcus aureus. Microorganisms 2019, 7, 471. [Google Scholar] [CrossRef]
- Kaźmierczak, N.; Grygorcewicz, B.; Piechowicz, L. Biofilm Formation and Prevalence of Biofilm-Related Genes among Clinical Strains of Multidrug-Resistant Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 956–964. [Google Scholar] [CrossRef]
- Wu, N.; Zhu, T. Potential of Therapeutic Bacteriophages in Nosocomial Infection Management. Front. Microbiol. 2021, 12, 638094. [Google Scholar] [CrossRef]
- Singh, A.; Padmesh, S.; Dwivedi, M.; Kostova, I. How Good Are Bacteriophages as an Alternative Therapy to Mitigate Biofilms of Nosocomial Infections. Infect. Drug Resist. 2022, 15, 503–532. [Google Scholar] [CrossRef]
- Jamal, M.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Ur Rahman, S.; Das, C.R. Isolation, Characterization and Efficacy of Phage MJ2 against Biofilm Forming Multi-Drug Resistant Enterobacter Cloacae. Folia Microbiol. 2019, 64, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Tkhilaishvili, T.; Wang, L.; Tavanti, A.; Trampuz, A.; Di Luca, M. Antibacterial Efficacy of Two Commercially Available Bacteriophage Formulations, Staphylococcal Bacteriophage and PYO Bacteriophage, against Methicillin-Resistant Staphylococcus aureus: Prevention and Eradication of Biofilm Formation and Control of a Systemic Infection of Galleria mellonella Larvae. Front. Microbiol. 2020, 11, 110. [Google Scholar] [PubMed]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage Treatment of Human Infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed]
- Górski, A.; Międzybrodzki, R.; Węgrzyn, G.; Jończyk-Matysiak, E.; Borysowski, J.; Weber-Dąbrowska, B. Phage Therapy: Current Status and Perspectives. Med. Res. Rev. 2020, 40, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K.; Abedon, S.T. Pharmacologically Aware Phage Therapy: Pharmacodynamic and Pharmacokinetic Obstacles to Phage Antibacterial Action in Animal and Human Bodies. Microbiol. Mol. Biol. Rev. MMBR 2019, 83, e00012-19. [Google Scholar] [CrossRef]
- Principi, N.; Silvestri, E.; Esposito, S. Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections. Front. Pharmacol. 2019, 10, 513. [Google Scholar] [CrossRef]
- Mangalea, M.R.; Duerkop, B.A. Fitness Trade-Offs Resulting from Bacteriophage Resistance Potentiate Synergistic Antibacterial Strategies. Infect. Immun. 2020, 88, e00926-19. [Google Scholar] [CrossRef]
- Kamal, F.; Dennis, J.J. Burkholderia Cepacia Complex Phage-Antibiotic Synergy (PAS): Antibiotics Stimulate Lytic Phage Activity. Appl. Environ. Microbiol. 2015, 81, 1132–1138. [Google Scholar] [CrossRef]
- Ghosh, A.; Ricke, S.C.; Almeida, G.; Gibson, K.E. Combined Application of Essential Oil Compounds and Bacteriophage to Inhibit Growth of Staphylococcus aureus In Vitro. Curr. Microbiol. 2016, 72, 426–435. [Google Scholar] [CrossRef]
- Jo, A.; Ding, T.; Ahn, J. Synergistic Antimicrobial Activity of Bacteriophages and Antibiotics against Staphylococcus aureus. Food Sci. Biotechnol. 2016, 25, 935–940. [Google Scholar] [CrossRef]
- Bruni, N.; Capucchio, M.T.; Biasibetti, E.; Pessione, E.; Cirrincione, S.; Giraudo, L.; Corona, A.; Dosio, F. Antimicrobial Activity of Lactoferrin-Related Peptides and Applications in Human and Veterinary Medicine. Molecules 2016, 21, 752. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.H.; Jenssen, H.; Gutteberg, T.J. Lactoferrin and Lactoferricin Inhibit Herpes Simplex 1 and 2 Infection and Exhibit Synergy When Combined with Acyclovir. Antivir. Res. 2003, 58, 209–215. [Google Scholar] [CrossRef]
- Mistry, N.; Drobni, P.; Näslund, J.; Sunkari, V.G.; Jenssen, H.; Evander, M. The Anti-Papillomavirus Activity of Human and Bovine Lactoferricin. Antivir. Res. 2007, 75, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.R.; Brewer, M.; Gauthier, J.J. Bactericidal Activity of Human Lactoferrin: Sensitivity of a Variety of Microorganisms. Infect. Immun. 1980, 28, 893–898. [Google Scholar] [CrossRef]
- Jahani, S.; Shakiba, A.; Jahani, L. The Antimicrobial Effect of Lactoferrin on Gram-Negative and Gram-Positive Bacteria. Int. J. Infect. 2015, 2, iji27594. [Google Scholar] [CrossRef]
- Ammons, M.C.B.; Ward, L.S.; Fisher, S.T.; Wolcott, R.D.; James, G.A. In Vitro Susceptibility of Established Biofilms Composed of a Clinical Wound Isolate of Pseudomonas Aeruginosa Treated with Lactoferrin and Xylitol. Int. J. Antimicrob. Agents 2009, 33, 230–236. [Google Scholar] [CrossRef]
- Petitclerc, D.; Lauzon, K.; Cochu, A.; Ster, C.; Diarra, M.S.; Lacasse, P. Efficacy of a Lactoferrin-Penicillin Combination to Treat β-Lactam-Resistant Staphylococcus aureus Mastitis. J. Dairy Sci. 2007, 90, 2778–2787. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Lauzon, K.; Diarra, M.S.; Petitclerc, D. Utilization of Lactoferrin to Fight Antibiotic-Resistant Mammary Gland Pathogens1,2. J. Anim. Sci. 2008, 86, 66–71. [Google Scholar] [CrossRef]
- Chen, P.-W.; Ho, S.-P.; Shyu, C.-L.; Mao, F.C. Effects of Bovine Lactoferrin Hydrolysate on the in Vitro Antimicrobial Susceptibility of Escherichia coli Strains Isolated from Baby Pigs. Am. J. Vet. Res. 2004, 65, 131–137. [Google Scholar] [CrossRef]
- Andrés, M.T.; Viejo-Diaz, M.; Pérez, F.; Fierro, J.F. Antibiotic Tolerance Induced by Lactoferrin in Clinical Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Antimicrob. Agents Chemother. 2005, 49, 1613–1616. [Google Scholar] [CrossRef]
- Ammons, M.C.B.; Ward, L.S.; Dowd, S.; James, G.A. Combined Treatment of Pseudomonas Aeruginosa Biofilm with Lactoferrin and Xylitol Inhibits the Ability of Bacteria to Respond to Damage Resulting from Lactoferrin Iron Chelation. Int. J. Antimicrob. Agents 2011, 37, 316–323. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Harris, M.R.; Coote, P.J. Combination of Caspofungin or Anidulafungin with Antimicrobial Peptides Results in Potent Synergistic Killing of Candida Albicans and Candida Glabrata in Vitro. Int. J. Antimicrob. Agents 2010, 35, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Leitch, E.C.; Willcox, M.D. Lactoferrin Increases the Susceptibility of S. epidermidis Biofilms to Lysozyme and Vancomycin. Curr. Eye Res. 1999, 19, 12–19. [Google Scholar] [CrossRef]
- Golshahi, L.; Lynch, K.H.; Dennis, J.J.; Finlay, W.H. In Vitro Lung Delivery of Bacteriophages KS4-M and ΦKZ Using Dry Powder Inhalers for Treatment of Burkholderia Cepacia Complex and Pseudomonas Aeruginosa Infections in Cystic Fibrosis. J. Appl. Microbiol. 2011, 110, 106–117. [Google Scholar] [CrossRef]
- Geagea, H.; Gomaa, A.; Remondetto, G.; Moineau, S.; Subirade, M. Molecular Structure of Lactoferrin Influences the Thermal Resistance of Lactococcal Phages. J. Agric. Food Chem. 2017, 65, 2214–2221. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Z.; Zhao, J.; Zhu, Z.; Yang, C.; Liu, Y. Prospects of Inhaled Phage Therapy for Combatting Pulmonary Infections. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Zimecki, M.; Artym, J.; Kocieba, M.; Weber-Dabrowska, B.; Lusiak-Szelachowska, M.; Górski, A. The Concerted Action of Lactoferrin and Bacteriophages in the Clearance of Bacteria in Sublethally Infected Mice. Postepy Hig. Med. Doswiadczalnej Online 2008, 62, 42–46. [Google Scholar]
- Kosznik-Kwaśnicka, K.; Ciemińska, K.; Grabski, M.; Grabowski, Ł.; Górniak, M.; Jurczak-Kurek, A.; Węgrzyn, G.; Węgrzyn, A. Characteristics of a Series of Three Bacteriophages Infecting Salmonella Enterica Strains. Int. J. Mol. Sci. 2020, 21, 6152. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosznik-Kwaśnicka, K.; Kaźmierczak, N.; Piechowicz, L. Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms. Antibiotics 2022, 11, 1256. https://doi.org/10.3390/antibiotics11091256
Kosznik-Kwaśnicka K, Kaźmierczak N, Piechowicz L. Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms. Antibiotics. 2022; 11(9):1256. https://doi.org/10.3390/antibiotics11091256
Chicago/Turabian StyleKosznik-Kwaśnicka, Katarzyna, Natalia Kaźmierczak, and Lidia Piechowicz. 2022. "Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms" Antibiotics 11, no. 9: 1256. https://doi.org/10.3390/antibiotics11091256
APA StyleKosznik-Kwaśnicka, K., Kaźmierczak, N., & Piechowicz, L. (2022). Activity of Phage–Lactoferrin Mixture against Multi Drug Resistant Staphylococcus aureus Biofilms. Antibiotics, 11(9), 1256. https://doi.org/10.3390/antibiotics11091256