
Identification of a Novel IncHI1B Plasmid in MDR Klebsiella pneumoniae 200 from Swine in China

Abstract
:
1. Introduction
2. Results and Discussion
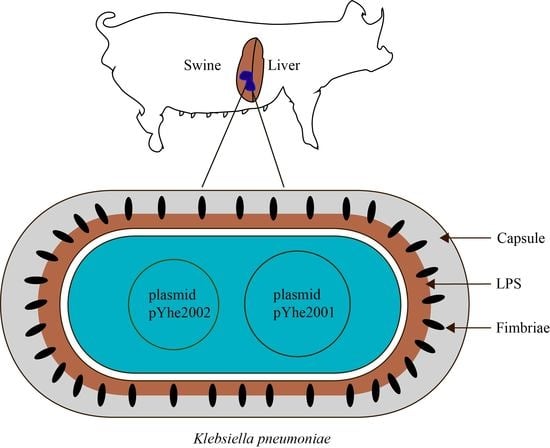
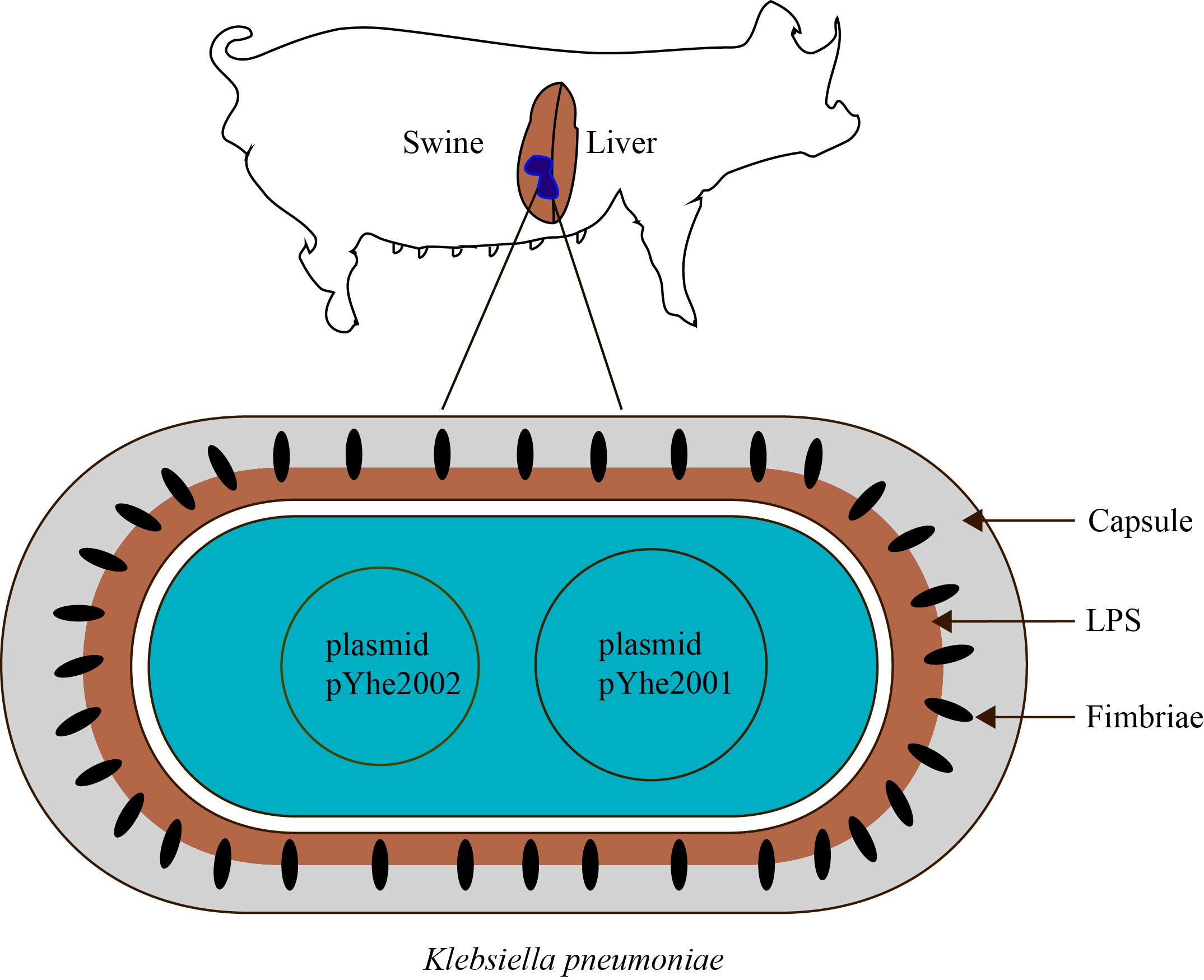
2.1. Characterization of K. pneumoniae 200
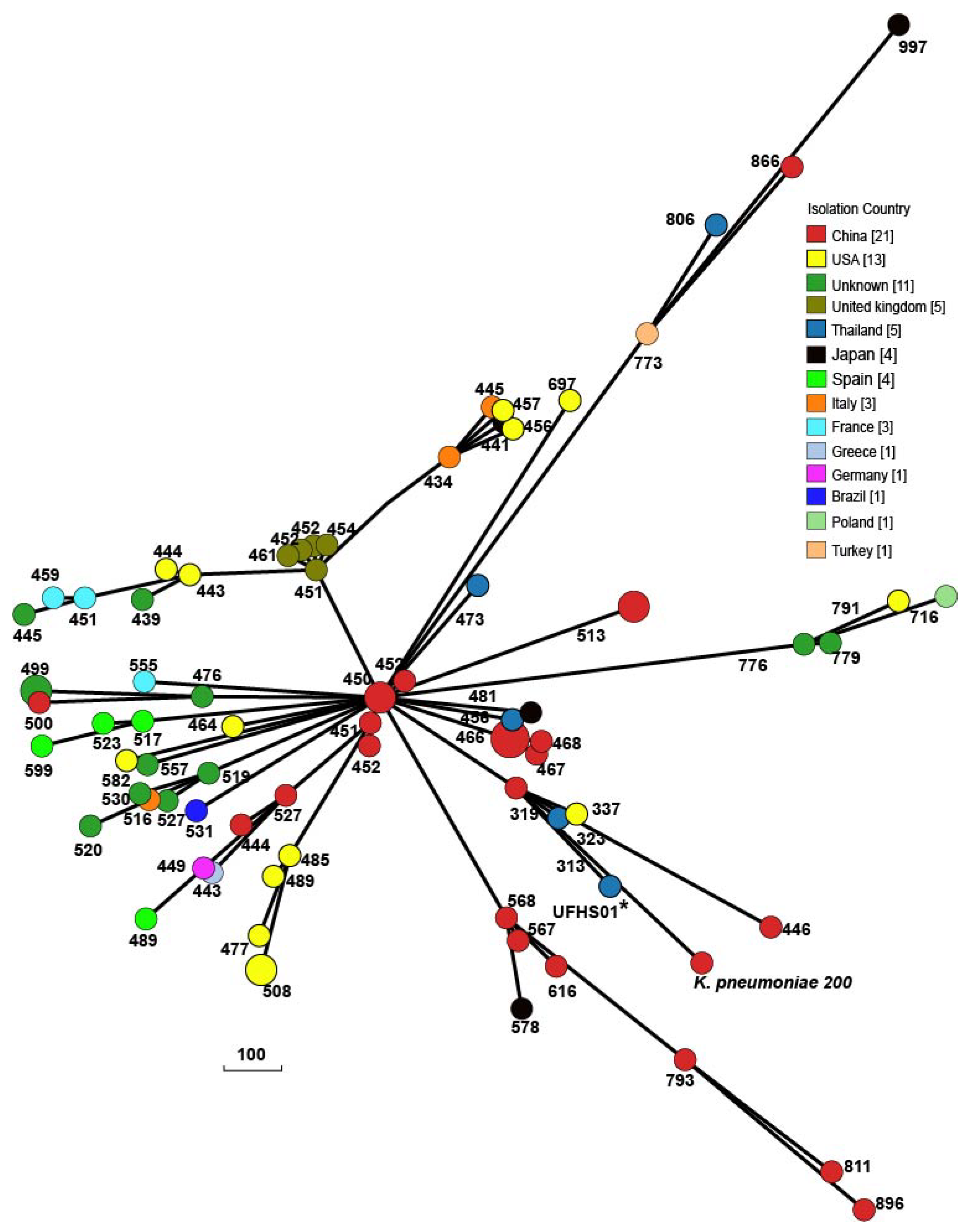
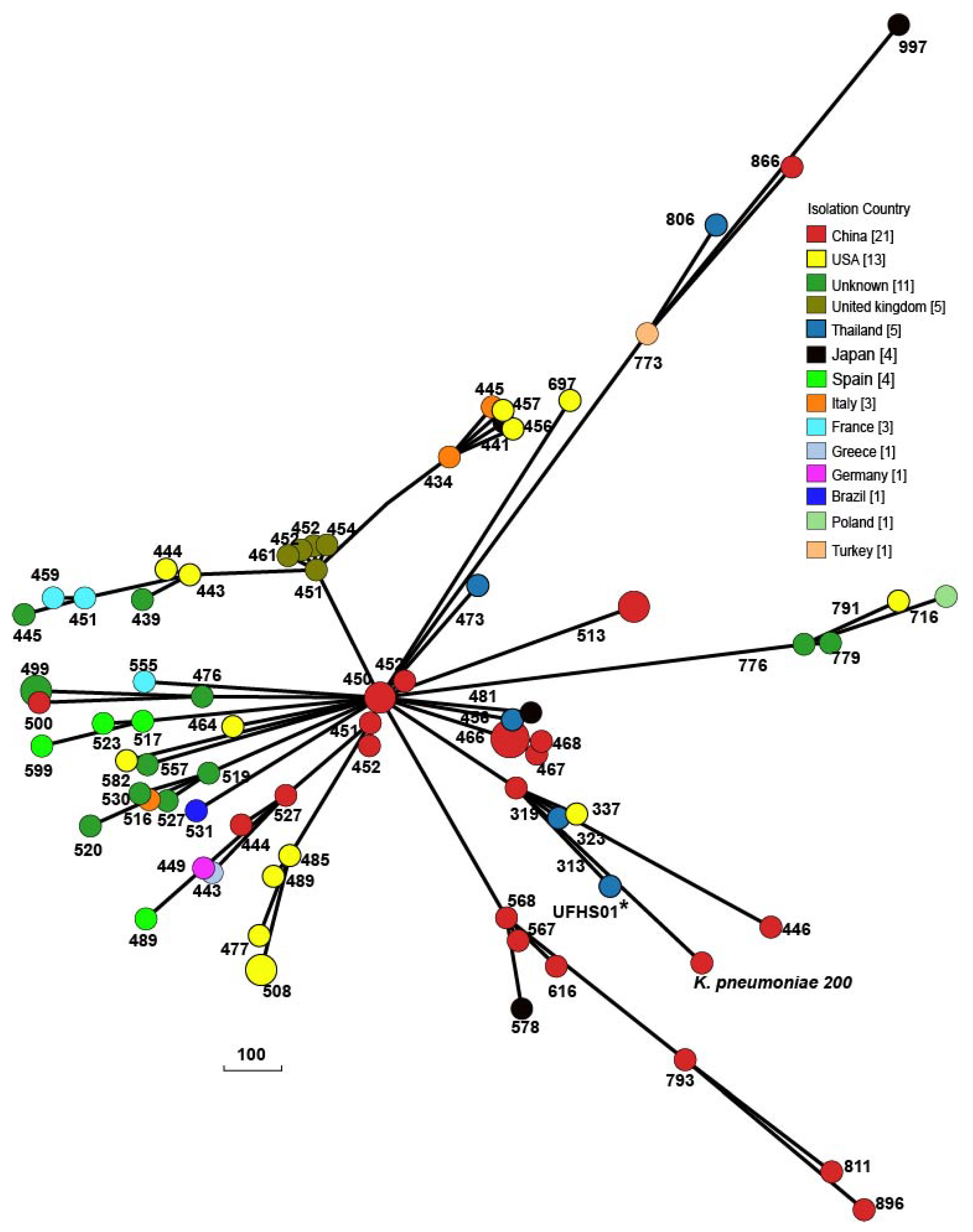
2.2. Characterization of Chromosome of K. pneumoniae 200
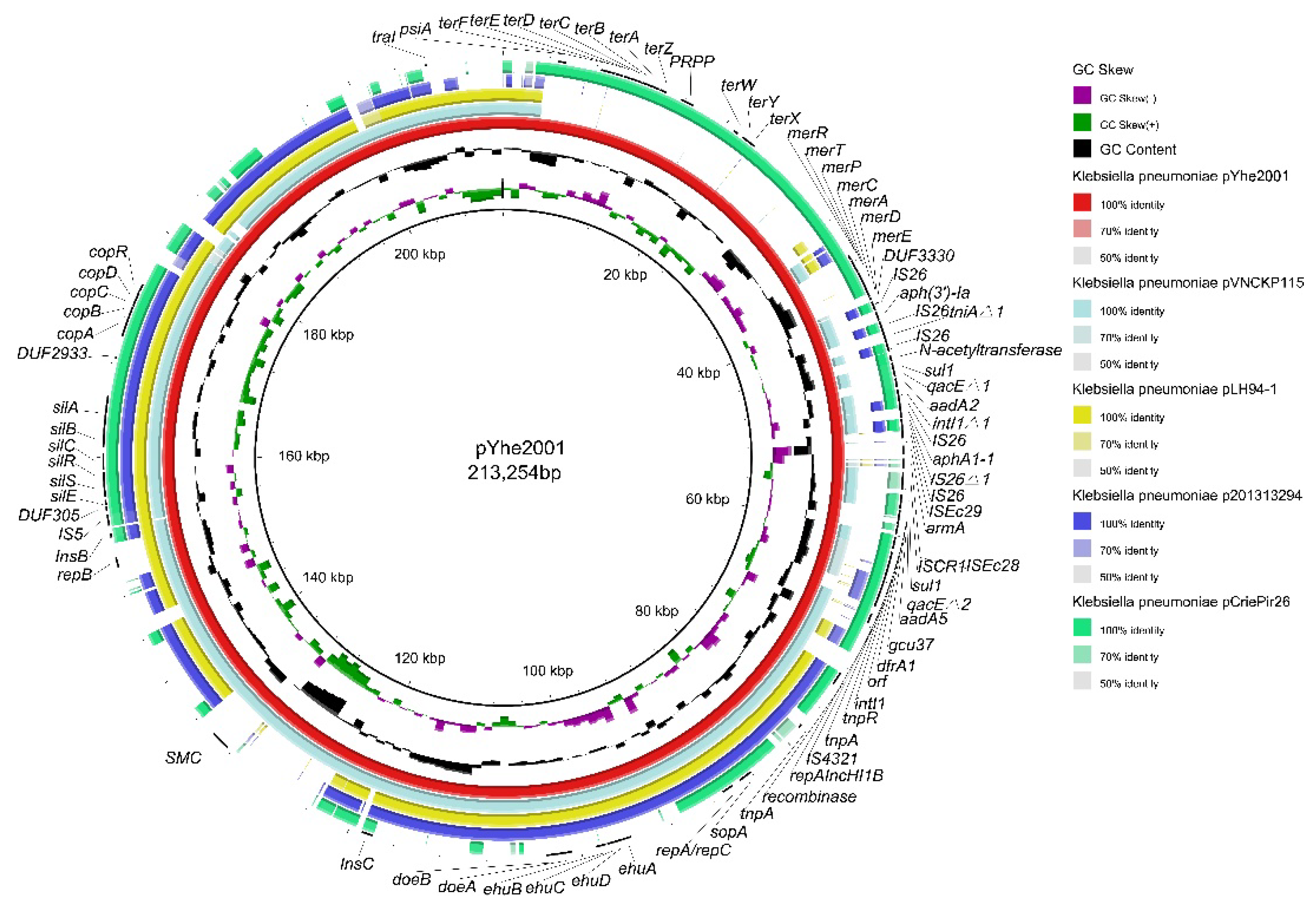
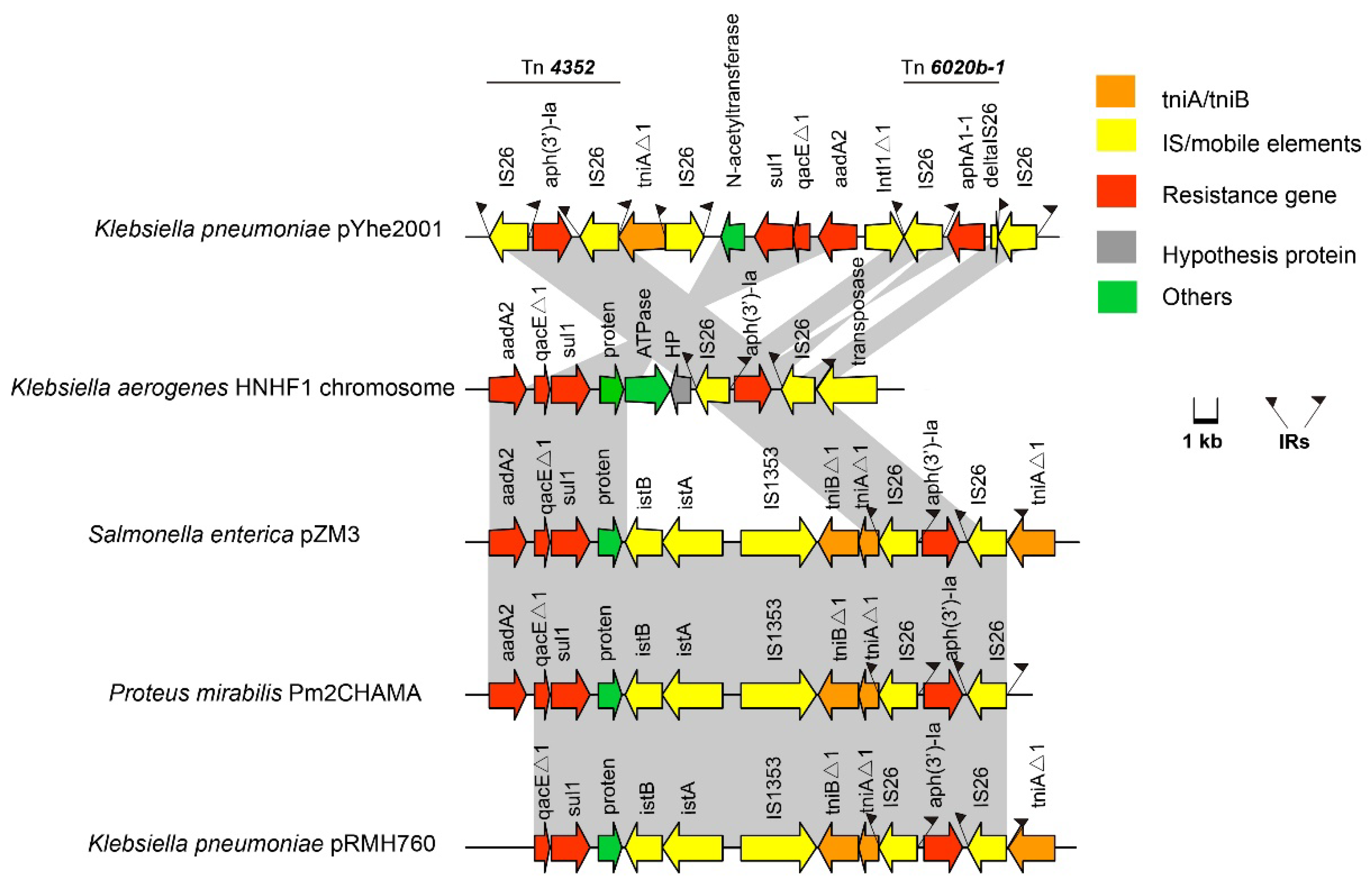
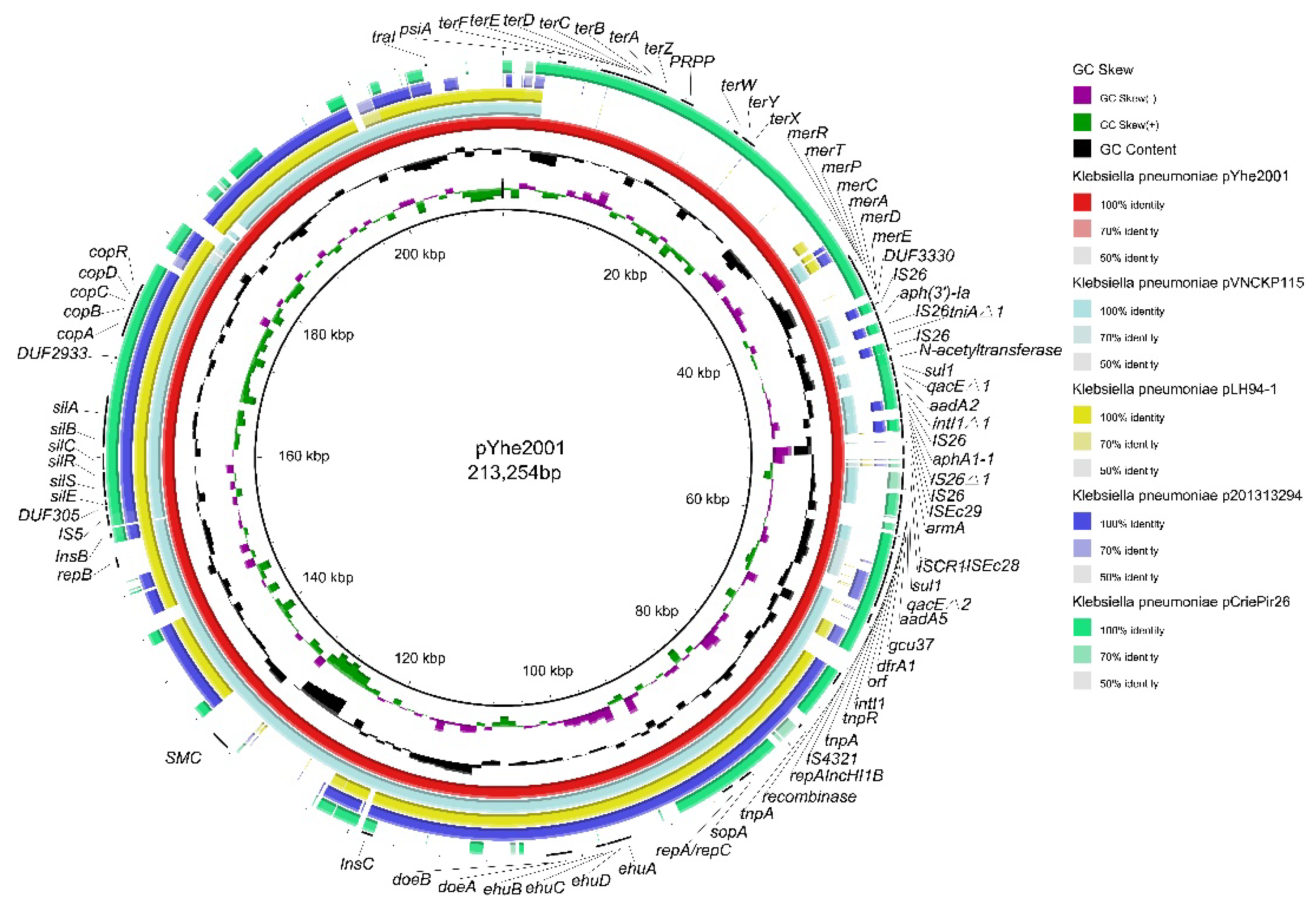
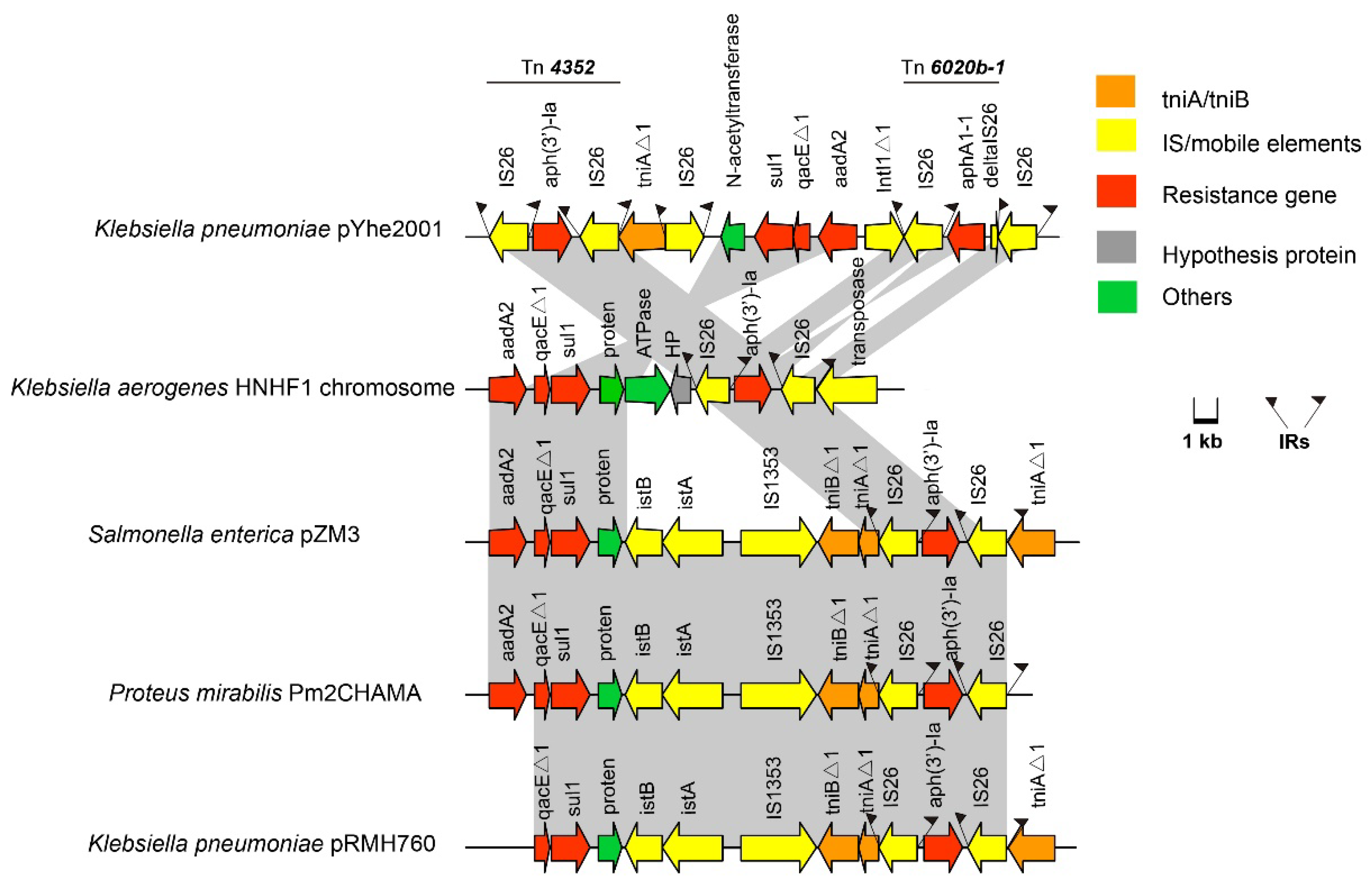
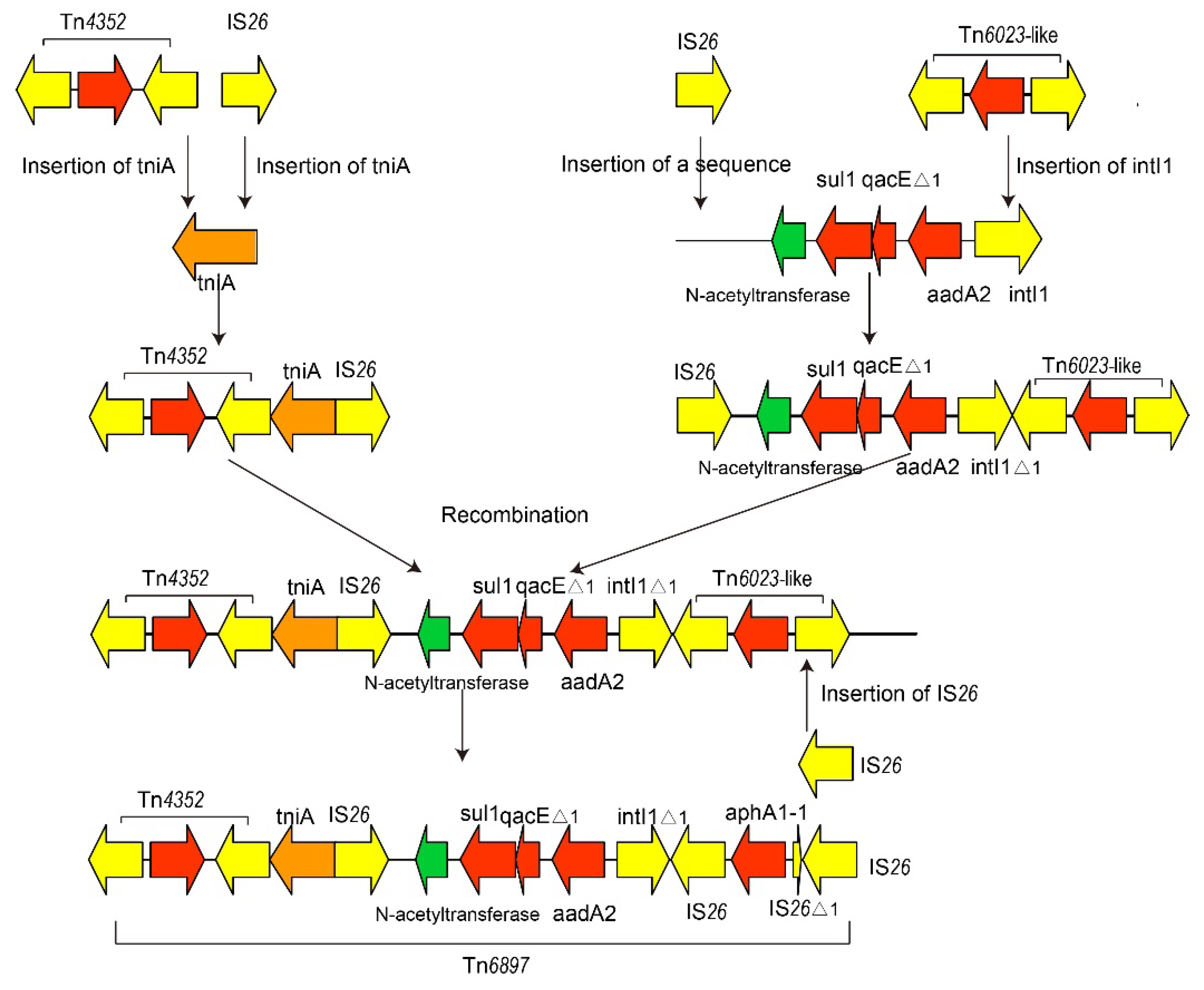
2.3. Characterization of Plasmids Carried by K. pneumoniae 200
2.4. Transformation Experiment
3. Materials and Methods
3.1. Bacterial Strain
3.2. Antimicrobial Susceptibility Testing
3.3. Genome Sequencing, Genome Assembly and Bioinformatics
3.4. Transformation Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, G.S.; Price, L.B. Recent research examining links among Klebsiella pneumoniae from food, food animals, and human extraintestinal infections. Curr. Environ. Health Rep. 2016, 3, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Grandemange, E.; Pommier, P.; Wessel-Robert, S.; Davot, J.L. Field evaluation of efficacy and tolerance of a 2% marbofloxacin injectable solution for the treatment of respiratory disease in fattening swines. Vet. Q. 2000, 22, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-producing Enterobacterales in swines in Portugal: Evidence of direct links with antibiotic selective pressure. Int. J. Antimicrob. Agents 2020, 55, 105802. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Liu, L.; Zhang, R.; Chen, K.; Xie, M.; Chan, E.; Chen, S. An IncR plasmid harbored by a hypervirulent carbapenem-resistant Klebsiella pneumoniae strain possesses five tandem repeats of the bla(KPC-2): NTEKPC-Id fragment. Antimicrob. Agents Chemother. 2019, 63, e01775-18. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Zhang, Y.; Liu, J.; Hu, X.; Nie, T.; Yang, X.; Wang, X.; Li, C.; You, X. Characterization of a hypervirulent multidrug-resistant ST23 Klebsiella pneumoniae carrying a blaCTX-M-24 IncFII plasmid and a pK2044-like plasmid. J. Glob. Antimicrob. Resist. 2020, 22, 674–679. [Google Scholar] [CrossRef]
- Naas, T.; Cuzon, G.; Villegas, M.V.; Lartigue, M.F.; Quinn, J.P.; Nordmann, P. Genetic structures at the origin of acquisition of the beta-lactamase blaKPC gene. Antimicrob. Agents Chemother. 2008, 52, 1257–1263. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.; Sebra, R.; Zhuge, J.; Yin, C.; Aguero-Rosenfeld, M.E.; Schuetz, A.N.; Dimitrova, N.; Fallon, J.T. Emergence and evolution of multidrug-resistant Klebsiella pneumoniae with both blaKPC and blaCTX-M integrated in the chromosome. Antimicrob. Agents Chemother. 2017, 61, e00076-17. [Google Scholar] [CrossRef]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Desmet, S.; Nepal, S.; van Dijl, J.M.; Van Ranst, M.; Chlebowicz, M.A.; Rossen, J.W.; Van Houdt, J.; Maes, P.; Lagrou, K.; Bathoorn, E. Antibiotic resistance plasmids cointegrated into a megaplasmid harboring the blaOXA-427 carbapenemase gene. Antimicrob. Agents Chemother. 2018, 62, e01448-17. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Spychala, C.N.; McElheny, C.L.; Doi, Y. Comparative analysis of an IncR plasmid carrying armA, blaDHA-1 and qnrB4 from Klebsiella pneumoniae ST37 isolates. J. Antimicrob. Chemother. 2016, 71, 882–886. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Dong, N.; Liu, C.; Zeng, Y.; Sun, Q.; Zhou, H.; Hu, Y.; Chen, S.; Shen, Z.; Zhang, R. Prevalence and molecular epidemiology of mcr-1-positive Klebsiella pneumoniae in healthy adults from China. J. Antimicrob. Chemother. 2020, 75, 2485–2494. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Elkhatib, W.F.; Ashour, H.M. Molecular typing and virulence analysis of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from Egyptian hospitals. Sci. Rep. 2016, 6, 38929. [Google Scholar] [CrossRef] [PubMed]
- Yuhan, Y.; Ziyun, Y.; Yongbo, Z.; Fuqiang, L.; Qinghua, Z. Overexpression of AdeABC and AcrAB-TolC efflux systems confers tigecycline resistance in clinical isolates of Acinetobacter baumannii and Klebsiella pneumoniae. Rev. Soc. Bras. Med. Trop. 2016, 49, 165–171. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, W.; Schwarz, S.; Zhu, Y.; Li, R.; Hua, X.; Liu, S. Genetic characterization of an MDR/virulence genomic element carrying two T6SS gene clusters in a clinical Klebsiella pneumoniae isolate of swine origin. J. Antimicrob. Chemother. 2019, 74, 1539–1544. [Google Scholar] [CrossRef]
- Kopotsa, K.; Osei Sekyere, J.; Mbelle, N.M. Plasmid evolution in carbapenemase-producing Enterobacteriaceae: A review. Ann. N. Y. Acad. Sci. 2019, 1457, 61–91. [Google Scholar] [CrossRef] [PubMed]
- Schmartz, G.P.; Hartung, A.; Hirsch, P.; Kern, F.; Fehlmann, T.; Müller, R.; Keller, A. PLSDB: Advancing a comprehensive database of bacterial plasmids. Nucleic Acids Res. 2022, 50, D273–D278. [Google Scholar] [CrossRef]
- Khezri, A.; Avershina, E.; Ahmad, R. Plasmid Identification and Plasmid-Mediated Antimicrobial Gene Detection in Norwegian Isolates. Microorganisms 2020, 9, 52. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Y.-H.; Wang, J.-Y.; Chang, M.-X.; Zhao, Q.-Y.; Jiang, H.-X. A Novel Structure Harboring blaCTX-M-27 on IncF Plasmids in Escherichia coli Isolated from Swine in China. Antibiotics 2021, 10, 387. [Google Scholar] [CrossRef]
- Kawamura, K.; Hayashi, K.; Matsuo, N.; Kitaoka, K.; Kimura, K.; Wachino, J.-I.; Kondo, T.; Iinuma, Y.; Murakami, N.; Fujimoto, S.; et al. Prevalence of CTX-M-Type Extended-Spectrum β-Lactamase-Producing Escherichia coli B2-O25-ST131 H30R Among Residents in Nonacute Care Facilities in Japan. Microb. Drug Resist. 2018, 24, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhong, Q.; Guo, Y.; Hang, Y.; Chen, Y.; Fang, X.; Xiao, Y.; Zhu, H.; Luo, H.; Yu, F.; et al. Emergence of the coexistence of mcr-1, blaNDM-5, and blaCTX-M-55 in Klebsiella pneumoniae ST485 clinical isolates in China. Infect. Drug Resist. 2021, 14, 3449–3458. [Google Scholar] [CrossRef]
- Stosic, M.S.; Leangapichart, T.; Lunha, K.; Jiwakanon, J.; Angkititrakul, S.; Järhult, J.D.; Magnusson, U.; Sunde, M. Novel mcr-3.40 variant co-located with mcr-2.3 and blaCTX-M-63 on an IncHI1B/IncFIB plasmid found in Klebsiella pneumoniae from a healthy carrier in Thailand. J. Antimicrob. Chemother. 2021, 76, 2218–2220. [Google Scholar] [CrossRef] [PubMed]
- Gancz, A.; Kondratyeva, K.; Cohen-Eli, D.; Navon-Venezia, S. Genomics and virulence of Klebsiella pneumoniae Kpnu95 ST1412 harboring a novel Incf plasmid encoding blactx-M-15 and qnrs1 causing community urinary tract infection. Microorganisms 2021, 9, 1022. [Google Scholar] [CrossRef]
- Mshana, E.S.; Hain, T.; Domann, E.; Lyamuya, E.F.; Chakraborty, T.; Imirzalioglu, C. Predominance of Klebsiella pneumoniaeST14 carrying CTX-M-15 causing neonatal sepsis in Tanzania. BMC Infect. Dis. 2013, 13, 466. [Google Scholar] [CrossRef]
- Yao, H.; Cheng, J.; Li, A.; Yu, R.; Zhao, W.; Qin, S.; Du, X. Molecular characterization of an IncFII(k) plasmid coharboring blaIMP-26 and tet(A) variant in a clinical Klebsiella pneumoniae isolate. Front. Microbiol. 2020, 11, 1610. [Google Scholar] [CrossRef] [PubMed]
- Hanczvikkel, A.; Fuzi, M.; Ungvari, E.; Toth, A. Transmissible silver resistance readily evolves in high-risk clone isolates of Klebsiella pneumoniae. Acta Microbiol. Immunol. Hung. 2018, 65, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Finley, P.J.; Norton, R.; Austin, C.; Mitchell, A.; Zank, S.; Durham, P. Unprecedented silver resistance in clinically isolated Enterobacteriaceae: Major implications for burn and wound management. Antimicrob. Agents Chemother. 2015, 59, 4734–4741. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; He, Z.; Kang, Y.; Yu, H.; Wang, J.; Du, P.; Zhang, Z.; Hu, S.; Gao, Z. Complete nucleotide sequence of pH11, an IncHI2 plasmid conferring multiantibiotic resistance and multiheavy metal resistance genes in a clinical Klebsiella pneumoniae isolate. Plasmid 2016, 86, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-T.; Chang, H.-Y.; Lai, Y.-C.; Pan, C.-C.; Tsai, S.-F.; Peng, H.-L. Sequencing and analysis of the large virulence plasmid pLVPK of Klebsiella pneumoniae CG43. Gene 2004, 337, 189–198. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Hickman, A.B.; Varani, A.M.; Siguier, P.; Chandler, M.; Dekker, J.P.; Dyda, F. Insertion Sequence IS 26 Reorganizes Plasmids in Clinically Isolated Multidrug-Resistant Bacteria by Replicative Transposition. mBio 2015, 6, e00762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubomura, A.; Sekizuka, T.; Onozuka, D.; Murakami, K.; Kimura, H.; Sakaguchi, M.; Oishi, K.; Hirai, S.; Kuroda, M.; Okabe, N. Truncated class 1 integron gene cassette arrays contribute to antimicrobial resistance of diarrheagenic Escherichia coli. BioMed Res. Int. 2020, 2020, 4908189. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Z.-L.; Huang, X.-Y.; Ma, Z.-B.; Guo, Z.-W.; Lv, L.-C.; Xia, Y.-B.; Zeng, L.; Song, Q.-H.; Liu, J.-H. Evolution and Comparative Genomics of F33:A−:B− Plasmids Carrying bla CTX-M-55 or bla CTX-M-65 in Escherichia coli and Klebsiella pneumoniae Isolated from Animals, Food Products, and Humans in China. mSphere 2018, 3, e00137-18. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, E.J.E.; Azevedo, R.P.; Lopes, A.T.S.; Rocha, J.M.; Albuquerque, G.R.; Wenceslau, A.A.; Miranda, F.R.; Rodrigues, D.D.P.; Maciel, B.M. Salmonella spp. in Wild Free-Living Birds from Atlantic Forest Fragments in Southern Bahia, Brazil. BioMed Res. Int. 2020, 2020, 7594136. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Yu, X.; Xu, H.; Guo, L.; Zhang, J.; Huang, C.; Shen, P.; Jiang, X.; Xiao, Y.; Li, L. Complete genome sequencing and genomic characterization of two Escherichia coli strains co-producing MCR-1 and NDM-1 from bloodstream infection. Sci. Rep. 2017, 7, 17885. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, Y.; Mei, Y.; Jiang, H.; Xie, Z.; Liu, H.; Chen, X.; Shen, P. Escherichia coli is naturally transformable in a novel transformation system. FEMS Microbiol. Lett. 2006, 265, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC (mg/L) | K. pneumonia 200 | Transformant | DH5α |
|---|---|---|---|
| Amikacin | >256 | 128 | 1 |
| Ampicillin | >256 | >256 | 2 |
| Ciprofloxacin | >256 | 32 | 0.008 |
| Chloramphenicol | >256 | 32 | 2 |
| Gentamicin | 256 | 32 | 0.125 |
| Kanamycin | >256 | 128 | 0.75 |
| Oxacillin | >256 | 64 | 0.25 |
| Streptomycin | 64 | 16 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Li, X.; Yan, H. Identification of a Novel IncHI1B Plasmid in MDR Klebsiella pneumoniae 200 from Swine in China. Antibiotics 2022, 11, 1225. https://doi.org/10.3390/antibiotics11091225
Liang H, Li X, Yan H. Identification of a Novel IncHI1B Plasmid in MDR Klebsiella pneumoniae 200 from Swine in China. Antibiotics. 2022; 11(9):1225. https://doi.org/10.3390/antibiotics11091225
Chicago/Turabian StyleLiang, Huixian, Xinhui Li, and He Yan. 2022. "Identification of a Novel IncHI1B Plasmid in MDR Klebsiella pneumoniae 200 from Swine in China" Antibiotics 11, no. 9: 1225. https://doi.org/10.3390/antibiotics11091225
APA StyleLiang, H., Li, X., & Yan, H. (2022). Identification of a Novel IncHI1B Plasmid in MDR Klebsiella pneumoniae 200 from Swine in China. Antibiotics, 11(9), 1225. https://doi.org/10.3390/antibiotics11091225