
Effect of Intramammary Dry Cow Antimicrobial Treatment on Fresh Cow’s Milk Microbiota in California Commercial Dairies

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
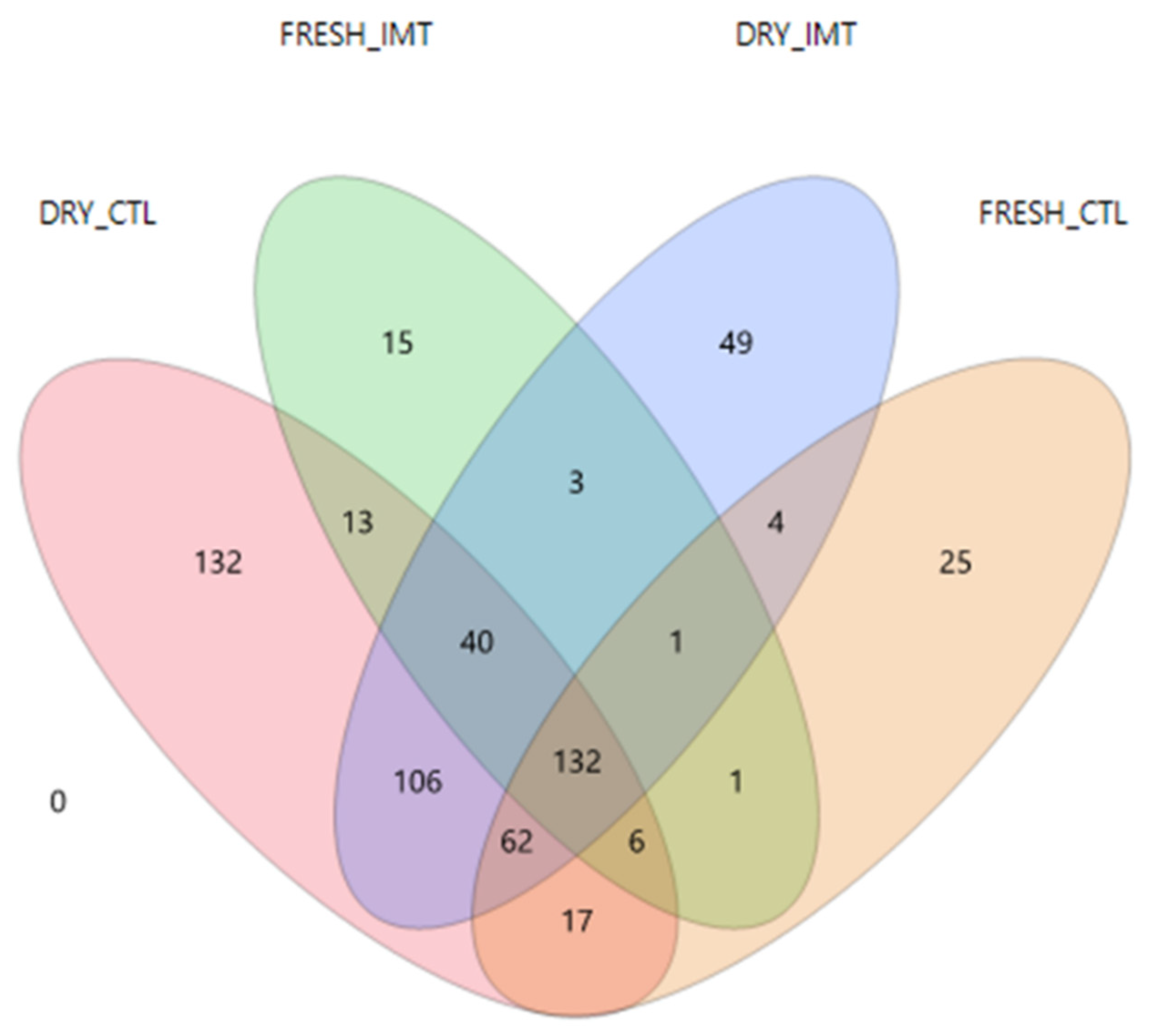
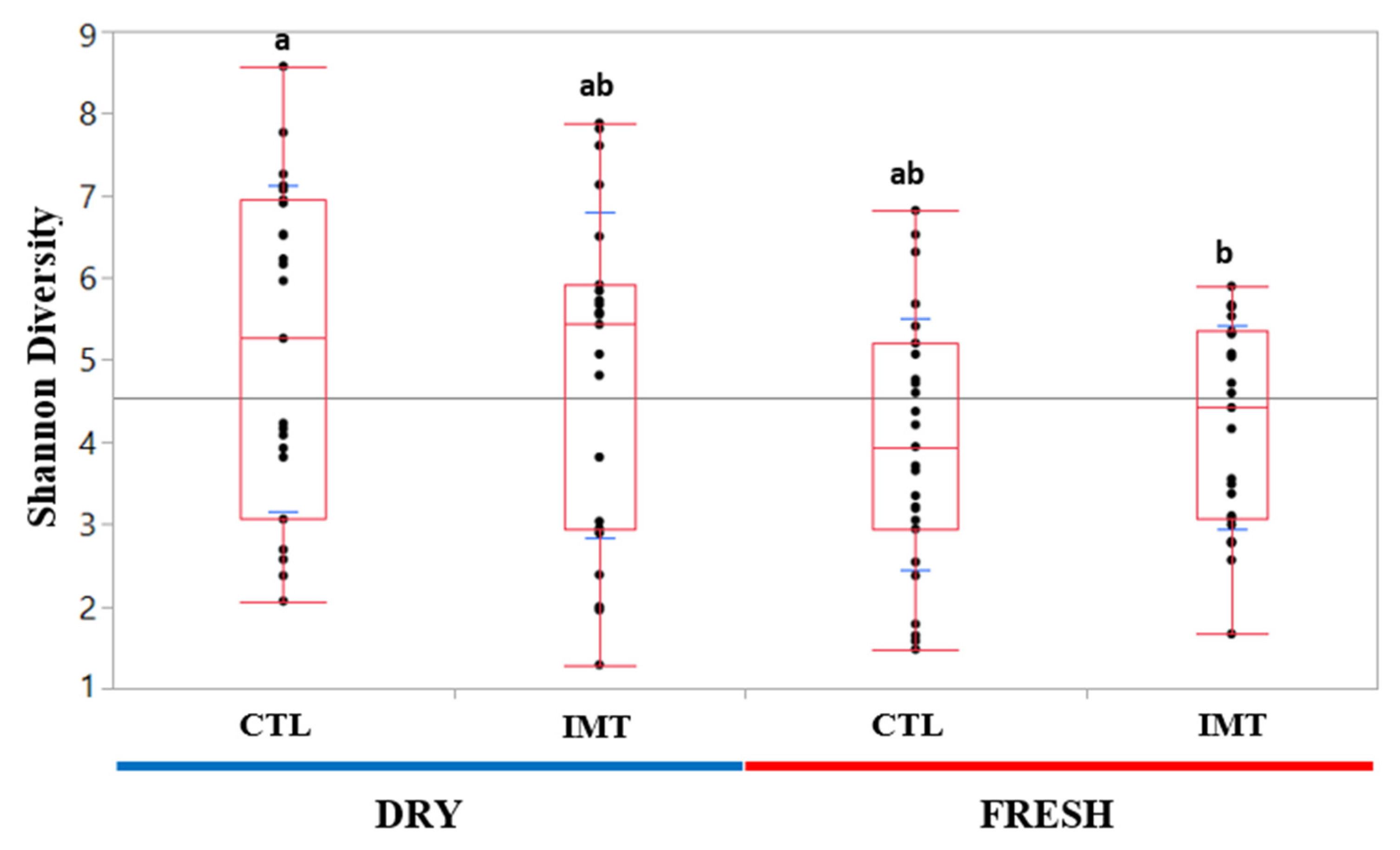
2.1. Descriptive Data and Microbial Diversity Data
2.2. Canonical Coefficients and Linear Regression
3. Discussion
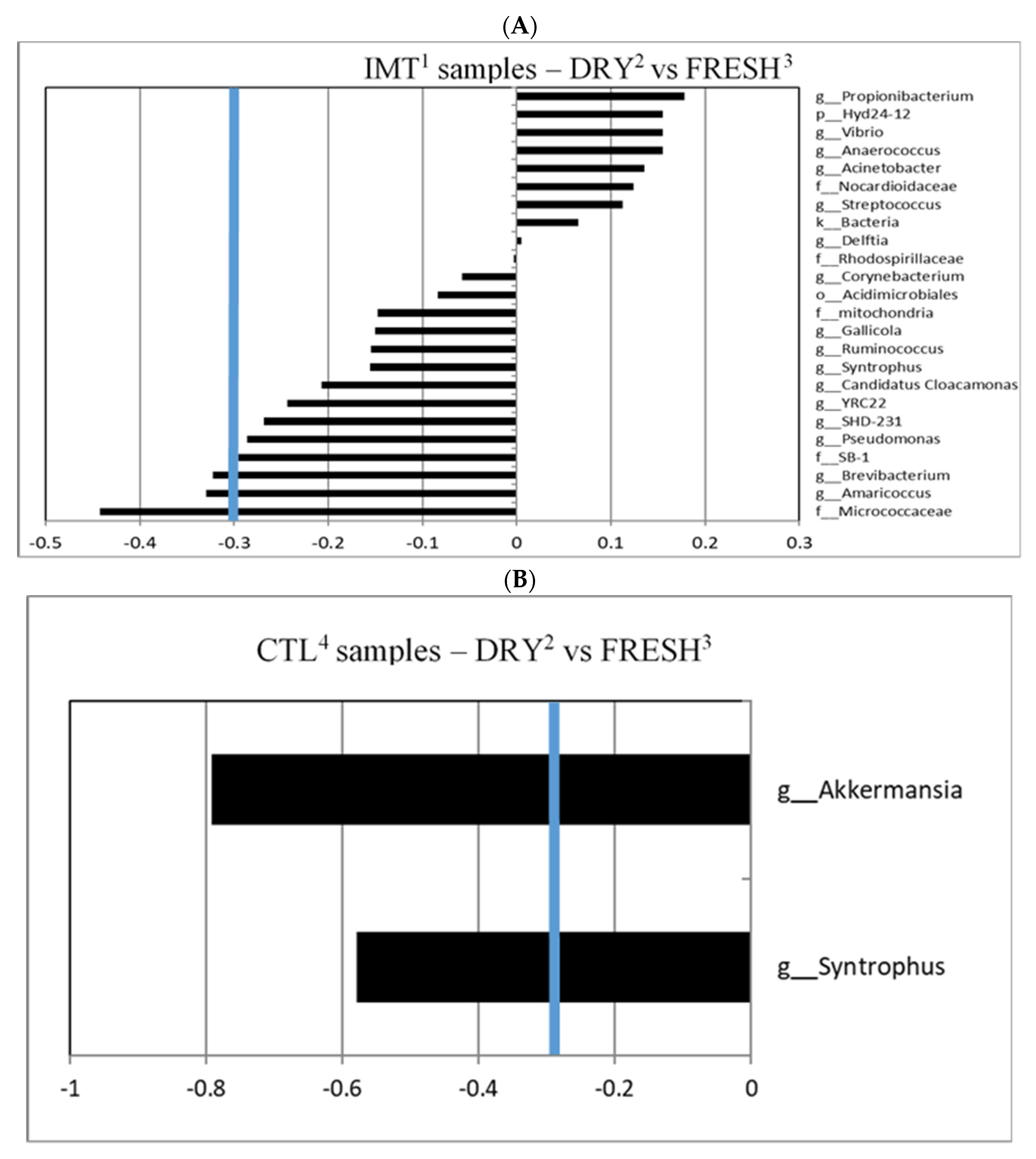
3.1. Lack of Microbiota Differentiation of FRESH Samples from CTL and IMT Cows
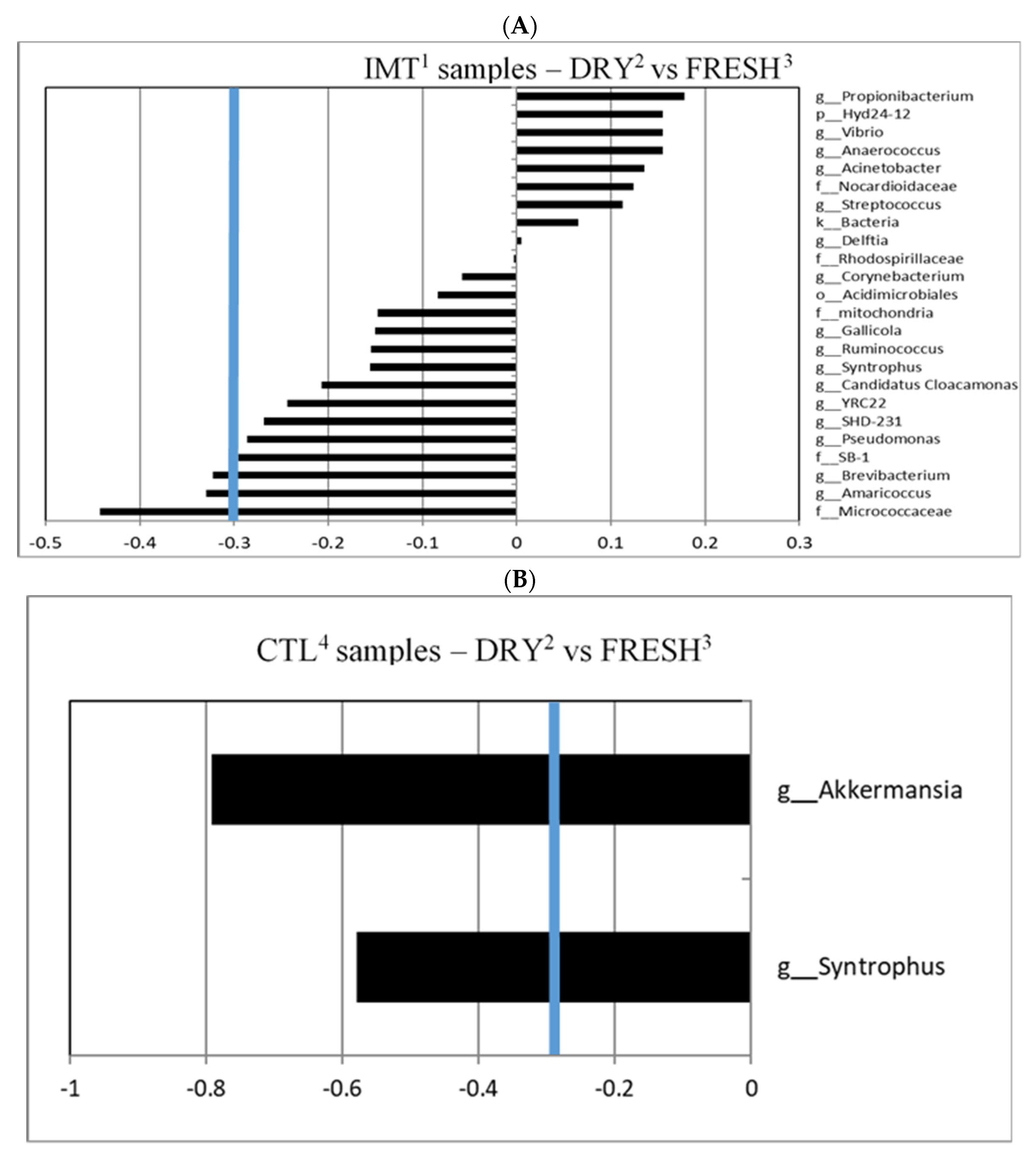
3.2. Microbiological Differentiation of DRY Samples from IMT and CTL Cows Compared to FRESH Samples
3.3. Differentiated Bacteria from DRY and FRESH Samples Collected from CTL Cows
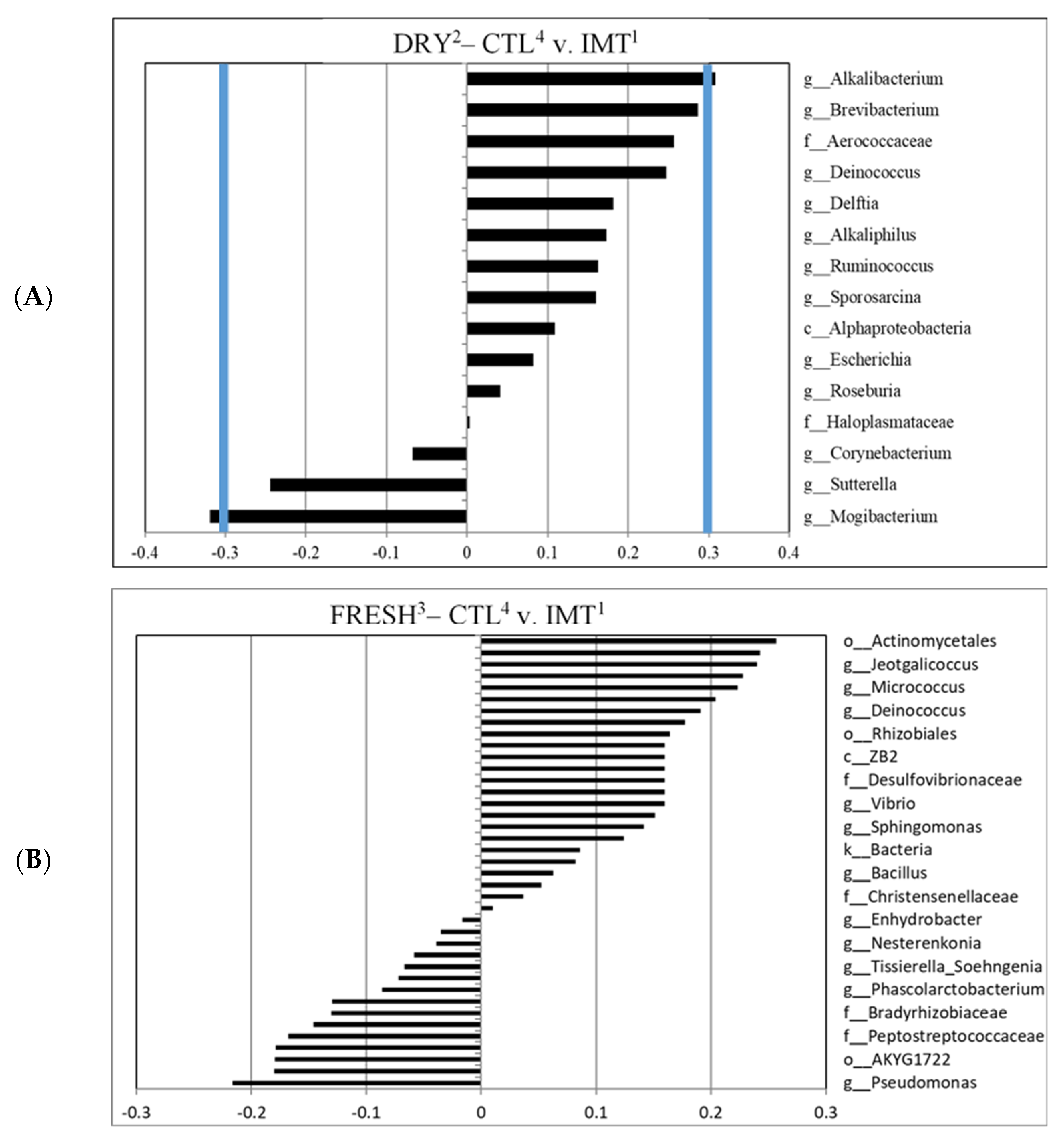
3.4. Differentiated Bacteria from DRY Samples Collected from IMT and CTL Cows
4. Material and Methods
4.1. Sampling
Milk Samples
4.2. DNA Extraction
4.3. PCR Amplification of 16S rRNA and DNA Sequencing
4.4. Bioinformatics
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cha, E.; Kristensen, A.R.; Hertl, J.; Schukken, Y.; Tauer, L.; Welcome, F.L.; Gröhn, Y. Optimal insemination and replacement decisions to minimize the cost of pathogen-specific clinical mastitis in dairy cows. J. Dairy Sci. 2014, 97, 2101–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, C.D.; Bradley, A.J.; Breen, J.E.; Green, M.J. Associations between udder health and reproductive performance in United Kingdom dairy cows. J. Dairy Sci. 2012, 95, 3683–3697. [Google Scholar] [CrossRef] [Green Version]
- Fuenzalida, M.J.; Fricke, P.M.; Ruegg, P.L. The association between occurrence and severity of subclinical and clinical mastitis on pregnancies per artificial insemination at first service of Holstein cows. J. Dairy Sci. 2015, 98, 3791–3805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA. Dairy 2014, Milk Quality, Milking Procedures, and Mastitisin the United States, 2014; USDA: Fort Collins, CO, USA, 2016.
- FDA. 2020 Summary Report on Antimicrobials Soldor Distributed for Usein Food-Producing Animals; Food and Drug Administration: Silver Spring, MD, USA, 2021.
- WHO. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151552-8. [Google Scholar]
- National Mastitis Council. Current Concepts of Bovine Mastitis, 5th ed.; National Mastitis Council: New Prague, MN, USA, 2017. [Google Scholar]
- Ward, G.E.; Schultz, L.H. Incidence and Control of Mastitis During the Dry Period. J. Dairy Sci. 1974, 57, 1341–1349. [Google Scholar] [CrossRef]
- Rindsig, R.B.; Rodewald, R.G.; Smith, A.; Spahr, S. Complete Versus Selective Dry Cow Therapy for Mastitis Control. J. Dairy Sci. 1978, 61, 1483–1497. [Google Scholar] [CrossRef]
- Woolford, M.W.; Williamson, J.H.; Day, A.M.; Copeman, P.J. The prophylactic effect of a teat sealer on bovine mastitis during the dry period and the following lactation. N. Z. Vet.-J. 1998, 46, 12–19. [Google Scholar] [CrossRef]
- Bonsaglia, E.C.; Gomes, M.S.; Canisso, I.F.; Zhou, Z.; Lima, S.F.; Rall, V.L.M.; Oikonomou, G.; Bicalho, R.C.; Lima, F.S.; Bonsaglia, E.C.; et al. Milk microbiome and bacterial load following dry cow therapy without antibiotics in dairy cows with healthy mammary gland. Sci. Rep. 2017, 7, 8067. [Google Scholar] [CrossRef] [Green Version]
- Huijps, K.; Hogeveen, H. Stochastic Modeling to Determine the Economic Effects of Blanket, Selective, and No Dry Cow Therapy. J. Dairy Sci. 2007, 90, 1225–1234. [Google Scholar] [CrossRef]
- Scherpenzeel, C.G.M.; Hogeveen, H.; Maas, L.; Lam, T. Economic optimization of selective dry cow treatment. J. Dairy Sci. 2018, 101, 1530–1539. [Google Scholar] [CrossRef]
- Vasquez, A.K.; Nydam, D.V.; Foditsch, C.; Wieland, M.; Lynch, R.; Eicker, S.; Virkler, P. Use of a culture-independent on-farm algorithm to guide the use of selective dry-cow antibiotic therapy. J. Dairy Sci. 2018, 101, 5345–5361. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, G.; Machado, V.S.; Santisteban, C.; Schukken, Y.; Bicalho, R.C. Microbial Diversity of Bovine Mastitic Milk as Described by Pyrosequencing of Metagenomic 16s rDNA. PLoS ONE 2012, 7, e47671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuehn, J.S.; Gorden, P.; Munro, D.; Rong, R.; Dong, Q.; Plummer, P.J.; Wang, C.; Phillips, G.J. Bacterial Community Profiling of Milk Samples as a Means to Understand Culture-Negative Bovine Clinical Mastitis. PLoS ONE 2013, 8, e61959. [Google Scholar] [CrossRef] [Green Version]
- Metzger, S.A.; Hernandez, L.L.; Skarlupka, J.; Walker, T.M.; Suen, G.; Ruegg, P.L. A Cohort Study of the Milk Microbiota of Healthy and Inflamed Bovine Mammary Glands from Dryoff Through 150 Days in Milk. Front. Vet.-Sci. 2018, 5, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganda, E.K.; Gaeta, N.C.; Sipka, A.; Pomeroy, B.; Oikonomou, G.; Schukken, Y.; Bicalho, R.C. Normal milk microbiome is reestablished following experimental infection with Escherichia coli independent of intramammary antibiotic treatment with a third-generation cephalosporin in bovines. Microbiome 2017, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Olsan, E.E.; Byndloss, M.; Faber, F.; Rivera-Chávez, F.; Tsolis, R.M.; Bäumler, A.J. Olonization resistance: The deconvolution of a complex trait. J. Biol. Chem. 2017, 292, 8577–8581. [Google Scholar] [CrossRef] [Green Version]
- Keeney, K.M.; Yurist-Doutsch, S.; Arrieta, M.-C.; Finlay, B.B. Effects of Antibiotics on Human Microbiota and Subsequent Disease. Annu. Rev. Microbiol. 2014, 68, 217–235. [Google Scholar] [CrossRef]
- Derakhshani, H.; Fehr, K.B.; Sepehri, S.; Francoz, D.; De Buck, J.; Barkema, H.; Plaizier, J.C.; Khafipour, E. Invited review: Microbiota of the bovine udder: Contributing factors and potential implications for udder health and mastitis susceptibility. J. Dairy Sci. 2018, 101, 10605–10625. [Google Scholar] [CrossRef] [Green Version]
- Aly, S.S.; Okello, E.; ElAshmawy, W.R.; Williams, D.R.; Anderson, R.J.; Rossitto, P.; Tonooka, K.; Glenn, K.; Karle, B.; Lehenbauer, T.W. Effectiveness of Intramammary Antibiotics, Internal Teat Sealants, or Both at Dry-Off in Dairy Cows: Clinical Mastitis and Culling Outcomes. Antibiotics 2022, 11, 954. [Google Scholar] [CrossRef]
- Lima, S.F.; Bicalho, M.L.S.; Bicalho, R.C. Evaluation of milk sample fractions for characterization of milk microbiota from healthy and clinical mastitis cows. PLoS ONE 2018, 13, e0193671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S rRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME2: Reproducible, interactive, scalable, and extensible microbiome data science. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Midway, S.; Robertson, M.; Flinn, S.; Kaller, M. Comparing multiple comparisons: Practical guidance for choosing the best multiple comparisons test. PeerJ 2020, 8, e10387. [Google Scholar] [CrossRef]
- Lambert, Z.V.; Durand, R.M. Some precautions in using canonical analysis. J. Mark. Res. 1975, 12, 468–475. [Google Scholar] [CrossRef]
- Jeon, S.J.; Neto, A.V.; Gobikrushanth, M.; Daetz, R.; Mingoti, R.D.; Parize, A.C.B.; de Freitas, S.L.; da Costa, A.N.L.; Bicalho, R.C.; Lima, S.; et al. Uterine Microbiota Progression from Calving until Establishment of Metritis in Dairy Cows. Appl. Environ. Microbiol. 2015, 81, 6324–6332. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Chiu, C.; Miao, M.; Wang, Y.; Settles, M.; Del Rio, N.S.; Castillo, A.; Souza, A.; Pereira, R.; Jeannotte, R. 16S rRNA analysis of diversity of manure microbial community in dairy farm environment. PLoS ONE 2018, 13, e0190126. [Google Scholar] [CrossRef]
- Derakhshani, H.; Plaizier, J.C.; De Buck, J.; Barkema, H.; Khafipour, E. Composition of the teat canal and intramammary microbiota of dairy cows subjected to antimicrobial dry cow therapy and internal teat sealant. J. Dairy Sci. 2018, 101, 10191–10205. [Google Scholar] [CrossRef] [Green Version]
- Ganda, E.K.; Bisinotto, R.S.; Lima, S.F.; Kronauer, K.; Decter, D.H.; Oikonomou, G.; Schukken, Y.; Bicalho, R.C. Longitudinal metagenomic profiling of bovine milk to assess the impact of intramammary treatment using a third-generation cephalosporin. Sci. Rep. 2016, 6, 37565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattray, F.P.; Fox, P.F. Aspects of Enzymology and Biochemical Properties of Brevibacterium linens Relevant to Cheese Ripening: A Review. J. Dairy Sci. 1999, 82, 891–909. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Christen, R.; Alexeeva, Y.V.; Zhukova, N.; Gorshkova, N.M.; Lysenko, A.M.; Mikhailov, V.V.; Nicolau, D.V. Brevibacterium celere sp. nov., isolated from degraded thallus of a brown alga. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 6, 2107–2111. [Google Scholar] [CrossRef]
- Bal, Z.S.; Sen, S.; Karapinar, D.Y.; Aydemir, S.; Vardar, F. The first reported catheter-related Brevibacterium casei bloodstream infection in a child with acute leukemia and review of the literature. Braz. J. Infect. Dis. 2015, 19, 213–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryssel, M.; Johansen, P.; Abu Al-Soud, W.; Sørensen, S.; Arneborg, N.; Jespersen, L. Microbial diversity and dynamics throughout manufacturing and ripening of surface ripened semi-hard Danish Danbo cheeses investigated by culture-independent techniques. Int. J. Food Microbiol. 2015, 215, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ekman, L.; Bagge, E.; Nyman, A.; Waller, K.P.; Pringle, M.; Segerman, B. A shotgun metagenomic investigation of the microbiota of udder cleft dermatitis in comparison to healthy skin in dairy cows. PLoS ONE 2020, 15, e0242880. [Google Scholar] [CrossRef] [PubMed]
- Maszenan, A.M.; Seviour, R.J.; Patel, B.; Rees, G.; McDougall, B.M. Amaricoccus gen. nov., a Gram-Negative Coccus Occurring in Regular Packages or Tetrads, Isolated from Activated Sludge Biomass, and Descriptions of Amaricoccus veronensis sp. nov., Amaricoccus tamworthensis sp. nov., Amaricoccus macauensis sp. nov., and Amaricoccus kaplicensis sp. nov. Int. J. Syst. Bacteriol. 1997, 47, 727–734. [Google Scholar] [CrossRef]
- Puyol, D.; Batstone, D.J.; Hülsen, T.; Astals, S.; Peces, M.; Krömer, J. Resource Recovery from Wastewater by Biological Technologies: Opportunities, Challenges, and Prospects. Front. Microbiol. 2017, 7, 2106. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.-X.; Gao, J.-F.; Dai, H.-H.; Zhang, W.-Z.; Zhang, D.; Wang, Z.-Q. DNA-based stable isotope probing identifies triclosan degraders in nitrification systems under different surfactants. Bioresour. Technol. 2020, 302, 122815. [Google Scholar] [CrossRef]
- Toscano, M.; De Grandi, R.; Peroni, D.G.; Grossi, E.; Facchin, V.; Comberiati, P.; Drago, L. Impact of delivery mode on the colostrum microbiota composition. BMC Microbiol. 2017, 17, 205. [Google Scholar] [CrossRef]
- Buffa, M.; Guamis, B.; Royo, C.; Trujillo, A.-J. Microbiological changes throughout ripening of goat cheese made from raw, pasteurized and high-pressure-treated milk. Food Microbiol. 2001, 18, 45–51. [Google Scholar] [CrossRef]
- Menéndez, S.; Godínez, R.; Centeno, J.; Rodríguez-Otero, J. Microbiological, chemical and biochemical characteristics of ‘Tetilla’ raw cows-milk cheese. Food Microbiol. 2001, 18, 151–158. [Google Scholar] [CrossRef]
- Cáceres, P.; Castillo, D.; Pizarro, M. Secondary flora of Casar de Cáceres cheese: Characterization of Micrococcaceae. Int. Dairy J. 1997, 7, 531–536. [Google Scholar] [CrossRef]
- Montel, M.-C.; Reitz, J.; Talon, R.; Berdagué, J.-L.; Rousset-Akrim, S. Biochemical activities of Micrococcaceae and their effects on the aromatic profiles and odours of a dry sausage model. Food Microbiol. 1996, 13, 489–499. [Google Scholar] [CrossRef]
- Garcia-Varona, M.; Santos, E.M.; Jaime, I.; Rovira, J. Characterisation of Micrococcaceae isolated from different varieties of chorizo. Int. J. Food Microbiol. 2000, 54, 189–195. [Google Scholar] [CrossRef]
- Chen, W.; Mi, J.; Lv, N.; Gao, J.; Cheng, J.; Wu, R.; Ma, J.; Lan, T.; Liao, X. Lactation Stage-Dependency of the Sow Milk Microbiota. Front. Microbiol. 2018, 9, 945. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-W.; Watanabe, K.; Hsu, C.-C.; Chao, S.-H.; Yang, Z.-H.; Lin, Y.-J.; Chen, C.-C.; Cao, Y.-M.; Huang, H.-C.; Chang, C.-H.; et al. Bacterial Composition and Diversity in Breast Milk Samples from Mothers Living in Taiwan and Mainland China. Front. Microbiol. 2017, 8, 965. [Google Scholar] [CrossRef]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef] [Green Version]
- Belzer, C.; De Vos, W.M. Microbes inside—From diversity to function: The case of Akkermansia. ISME J. 2012, 6, 1449–1458. [Google Scholar] [CrossRef]
- Cani, P.D.; De Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, S.; Ma, C.; Huo, D.; Peng, Q.; Shao, Y.; Zhang, J. Evaluation of the nutrition and function of cow and goat milk based on intestinal microbiota by metagenomic analysis. Food Funct. 2018, 9, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Lackey, K.A.; Williams, J.E.; Meehan, C.L.; Zachek, J.A.; Benda, E.D.; Price, W.J.; Foster, J.A.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; et al. What’s Normal? Microbiomes in Human Milk and Infant Feces Are Related to Each Other but Vary Geographically: The INSPIRE Study. Front. Nutr. 2019, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Ji, S.; Yan, H.; Hao, Y.; Zhang, J.; Wang, Y.; Cao, Z.; Li, S. The day-to-day stability of the ruminal and fecal microbiota in lactating dairy cows. Microbiol. Open 2020, 9, e990. [Google Scholar] [CrossRef] [PubMed]
- De Godoi, L.A.G.; Fuess, L.T.; Delforno, T.P.; Foresti, E.; Damianovic, M. Characterizing phenol-removing consortia under methanogenic and sulfate-reducing conditions: Potential metabolic pathways. Environ. Technol. 2018, 40, 3216–3226. [Google Scholar] [CrossRef]
- McInerney, M.J.; Rohlin, L.; Mouttaki, H.; Kim, U.; Krupp, R.S.; Rios-Hernandez, L.; Sieber, J.; Struchtemeyer, C.G.; Bhattacharyya, A.; Campbell, J.W.; et al. The genome of Syntrophus aciditrophicus: Life at the thermodynamic limit of microbial growth. Proc. Natl. Acad. Sci. USA 2007, 104, 7600–7605. [Google Scholar] [CrossRef] [Green Version]
- Spirito, C.M.; Daly, S.E.; Werner, J.J.; Angenent, L.T. Redundancy in Anaerobic Digestion Microbiomes during Disturbances by the Antibiotic Monensin. Appl. Environ. Microbiol. 2018, 84, e02692-17. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Tanasupawat, S.; Nakajima, K.; Kanamori, H.; Ishizaki, S.; Kodama, K.; Okamoto-Kainuma, A.; Koizumi, Y.; Yamamoto, Y.; Yamasato, K. Alkalibacterium thalassium sp. nov., Alkalibacterium pelagium sp. nov., Alkalibacterium putridalgicola sp. nov. and Alkalibacterium kapii sp. nov., slightly halophilic and alkaliphilic marine lactic acid bacteria isolated from marine organisms and salted foods collected in Japan and Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59 Pt 5, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Yunita, D.; Dodd, C.E. Microbial community dynamics of a blue-veined raw milk cheese from the United Kingdom. J. Dairy Sci. 2018, 101, 4923–4935. [Google Scholar] [CrossRef]
- Lucena-Padrós, H.; Jiménez, E.; Maldonado-Barragán, A.; Rodríguez, J.M.; Ruiz-Barba, J.L. PCR-DGGE assessment of the bacterial diversity in Spanish-style green table-olive fermentations. Int. J. Food Microbiol. 2015, 205, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, T.; Aino, K.; Narihiro, T.; Matsuyama, H.; Yumoto, I. Analysis of microbiota involved in the aged natural fermentation of indigo. World J. Microbiol. Biotechnol. 2017, 33, 70. [Google Scholar] [CrossRef]
- Ramanathan, T.; Ting, Y.-P. Alkaline bioleaching of municipal solid waste incineration fly ash by autochthonous extremophiles. Chemosphere 2016, 160, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Schwenninger, S.M.; Eugster-Meier, E.; Lacroix, C. Facultative anaerobic halophilic and alkaliphilic bacteria isolated from a natural smear ecosystem inhibit Listeria growth in early ripening stages. Int. J. Food Microbiol. 2011, 147, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Huo, W.; Zhu, W.; Mao, S. Characterization of bacterial community of raw milk from dairy cows during subacute ruminal acidosis challenge by high-throughput sequencing. J. Sci. Food Agric. 2014, 95, 1072–1079. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAXON 3 | Treatment/Time, % (SE) | |||
|---|---|---|---|---|
| CTL 1 | IMT 2 | |||
| DRY 4 | FRESH 5 | DRY 4 | FRESH | |
| g__Staphylococcus | 21.3 (4.7) | 19.2 (5.6) | 13.7 (3.7) | 15.7 (3.9) |
| g__Delftia | 11.4 (3.8) | 19.7 (5.2) | 19.9 (6.3) | 20 (5) |
| f__Peptostreptococcaceae | 7.6 (1.7) | 5.1 (1.2) | 6.2 (0.9) | 3.0 (1.0) |
| f__Ruminococcaceae | 4.8 (1.2) | 5.3 (1.2) | 5.7 (1.8) | 4.7 (1.3) |
| g__Corynebacterium | 3.9 (1.3) | 2.2 (0.9) | 3.1 (1.2) | 2.5 (0.5) |
| g__Turicibacter | 3.3 (0.8) | 1.9 (0.6) | 2.8 (0.6) | 3.3 (1.2) |
| o__Clostridiales | 2.7 (0.6) | 1.2 (0.3) | 3.1 (0.9) | 1.8 (0.6) |
| f__Lachnospiraceae | 2.6 (0.6) | 1.4 (0.4) | 1.7 (0.6) | 2.1 (0.7) |
| g__Serratia | 0.7 (0.7) | 2.5 (2.5) | 1.6 (1.6) | 1.5 (1.5) |
| o__Bacteroidales | 1.1 (0.4) | 1.2 (0.5) | 1.2 (0.5) | 2.3 (1.4) |
| g__Epulopiscium | 0.6 (3.4) | 4 (3.6) | 0.1 (0) | 0.1 (0.1) |
| g__Streptococcus | 1 (0.4) | 1.2 (0.9) | 0.7 (0.3) | 2.2 (2.1) |
| g__Acinetobacter | 1.3 (1.0) | 0.8 (0.5) | 1.1 (0.4) | 1.7 (0.6) |
| f__Clostridiaceae | 1.5 (0.4) | 0.9 (0.3) | 1 (0.3) | 1.3 (0.6) |
| g__5-7N15 | 1.3 (0.5) | 1.1 (0.4) | 1.2 (0.5) | 1.0 (3.7) |
| g__Salinicoccus | 1.1 (0.6) | 0.4 (0.2) | 1.4 (0.6) | 1.7 (1.2) |
| g__Herbaspirillum | 0.7 (0.6) | 0.4 (0.2) | 0 (0) | 3.5 (2.4) |
| f__Neisseriaceae | 0.9 (0.5) | 1.0 (0.7) | 1.1 (7.6) | 0.8 (0.7) |
| g__Bacillus | 0.8 (0.3) | 0.7 (0.3) | 1.2 (0.4) | 0.8 (0.5) |
| f__Aerococcaceae | 0.5 (0.2) | 0.5 (0.2) | 1.2 (0.4) | 1.2 (0.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basbas, C.; Aly, S.; Okello, E.; Karle, B.M.; Lehenbauer, T.; Williams, D.; Ganda, E.; Wiedmann, M.; Pereira, R.V. Effect of Intramammary Dry Cow Antimicrobial Treatment on Fresh Cow’s Milk Microbiota in California Commercial Dairies. Antibiotics 2022, 11, 963. https://doi.org/10.3390/antibiotics11070963
Basbas C, Aly S, Okello E, Karle BM, Lehenbauer T, Williams D, Ganda E, Wiedmann M, Pereira RV. Effect of Intramammary Dry Cow Antimicrobial Treatment on Fresh Cow’s Milk Microbiota in California Commercial Dairies. Antibiotics. 2022; 11(7):963. https://doi.org/10.3390/antibiotics11070963
Chicago/Turabian StyleBasbas, Carl, Sharif Aly, Emmanuel Okello, Betsy M. Karle, Terry Lehenbauer, Deniece Williams, Erika Ganda, Martin Wiedmann, and Richard V. Pereira. 2022. "Effect of Intramammary Dry Cow Antimicrobial Treatment on Fresh Cow’s Milk Microbiota in California Commercial Dairies" Antibiotics 11, no. 7: 963. https://doi.org/10.3390/antibiotics11070963
APA StyleBasbas, C., Aly, S., Okello, E., Karle, B. M., Lehenbauer, T., Williams, D., Ganda, E., Wiedmann, M., & Pereira, R. V. (2022). Effect of Intramammary Dry Cow Antimicrobial Treatment on Fresh Cow’s Milk Microbiota in California Commercial Dairies. Antibiotics, 11(7), 963. https://doi.org/10.3390/antibiotics11070963