
Resveratrol Reverts Tolerance and Restores Susceptibility to Chlorhexidine and Benzalkonium in Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antimicrobial Activity of CHX and BZK against a Panel of Reference Strains and Clinical Isolates of Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts
2.2. Effect of Carbonyl Cyanide M-Chlorophenylhydrazine Protonophore (CCCP) EP Inhibitor on CHX and BZK MIC and MBC against Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts
2.3. Effect of RV on CHX and BZK MIC and MBC (or MFC) against Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts
2.4. Effect of RV on CHX and BZK Combination against Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts
3. Materials and Methods
3.1. Bacterial Strain, Growth Condition and Reagents
3.2. Determination of Minimum Inhibitory Concentration and Minimum Bactericidal Concentration
3.3. In Vitro Combination Studies
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durante-Mangoni, E.; Utili, R.; Zarrilli, R. Combination therapy in severe Acinetobacter baumannii infections: An update on the evidence to date. Future Microbiol. 2014, 9, 773–789. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, 00031-19. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Treatment of Staphylococcus aureus infections. Rev. Microbiol. Immunol. 2017, 409, 325–383. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, 00181-19. [Google Scholar] [CrossRef] [PubMed]
- Zemanick, E.T.; Wagner, B.D.; Robertson, C.E.; Ahrens, R.C.; Chmiel, J.F.; Clancy, J.P.; Gibson, R.L.; Harris, W.T.; Kurland, G.; Laguna, T.A.; et al. Airway microbiota across age and disease spectrum in cystic fibrosis. Eur. Respir. J. 2017, 50, 1700832. [Google Scholar] [CrossRef] [PubMed]
- Martin-Loeches, I.; Antonelli, M.; Cuenca-Estrella, M.; Dimopoulos, G.; Einav, S.; De Waele, J.J.; Garnacho-Montero, J.; Kanj, S.S.; Machado, F.R.; Montravers, P.; et al. ESICM/ESCMID task force on practical management of invasive candidiasis in critically ill patients. Intensive Care Med. 2019, 45, 789–805. [Google Scholar] [CrossRef]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- Milstone, A.M.; Passaretti, C.L.; Perl, T.M. Chlorhexidine: Expanding the armamentarium for infection control and prevention. Clin. Infect. Dis. 2008, 46, 274–281. [Google Scholar] [CrossRef]
- Merchel, P.P.B.; Tagkopoulos, I. Benzalkonium chlorides: Uses, regulatory status, and microbial resistance. Appl. Environ. Microbiol. 2019, 85, 00377-19. [Google Scholar] [CrossRef] [Green Version]
- Kampf, G. Acquired resistance to chlorhexidine—Is it time to establish an ‘antiseptic stewardship’ initiative? J. Hosp. Infect. 2016, 94, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.J.; Rutala, W.A.; Sickbert-Bennett, E.E. Use of germicides in health care settings-is there a relationship between germicide use and antimicrobial resistance: A concise review. Am. J. Infect. Control 2019, 47, A106–A109. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Zhang, Z.; Shen, R.; Liu, X.; Li, X.; Chen, B.; Wu, X.; Li, H.; Xie, X.; Huang, S. Disinfection Strategies for Carbapenem-Resistant Klebsiella pneumoniae in a Healthcare Facility. Antibiotics 2022, 11, 736. [Google Scholar] [CrossRef] [PubMed]
- Abuzaid, A.; Hamouda, A.; Amyes, S.G. Klebsiella pneumoniae susceptibility to biocides and its association with cepA, qacDE and qacE efflux pump genes and antibiotic resistance. J. Hosp. Infect. 2012, 81, 87–91. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, X.; Zhou, B.; Liu, S.; Chen, W.; Chen, L.; Zhang, Y.; Liao, W.; Zeng, W.; Wu, Q.; et al. Clinical characteristics, tolerance mechanisms, and molecular epidemiology of reduced susceptibility to chlorhexidine among Pseudomonas aeruginosa isolated from a teaching hospital in China. Int. J. Antimicrob. Agents 2022, 60, 106605. [Google Scholar] [CrossRef]
- Rajamohan, G.; Srinivasan, V.B.; Gebreyes, W.A. Novel role of Acinetobacter baumannii RND efflux transporters in mediating decreased susceptibility to biocides. J. Antimicrob. Chemother. 2010, 65, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Du, D.; Wang-Kan, X.; Neuberger, A.; Van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Tucker, A.T.; Nowicki, E.M.; Boll, J.M.; Knauf, G.A.; Burdis, N.C.; Trent, M.S.; Davies, B.W. Defining gene-phenotype relationships in Acinetobacter baumannii through one-step chromosomal gene inactivation. mBio 2014, 5, 01313–01314. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, A.; Esposito, E.P.; Bagattini, M.; Berisio, R.; Triassi, M.; De Gregorio, E.; Zarrilli, R. Inhibition of AdeB, AceI, and AmvA Efflux Pumps Restores Chlorhexidine and Benzalkonium Susceptibility in Acinetobacter baumannii ATCC 19606. Front. Microbiol. 2022, 12, 790263. [Google Scholar] [CrossRef]
- Hassan, K.A.; Jackson, S.M.; Penesyan, A.; Patching, S.G.; Tetu, S.G.; Eijkelkamp, B.A.; Brown, M.H.; Henderson, P.J.F.; Paulsen, I.T. Transcriptomic and biochemical analyses identify a family of chlorhexidine efflux proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20254–20259. [Google Scholar] [CrossRef] [Green Version]
- Mattio, L.M.; Catinella, G.; Dallavalle, S.; Pinto, A. Stilbenoids: A natural arsenal against bacterial pathogens. Antibiotics 2020, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.S.L.; Tan, L.T.; Chan, K.G.; Yap, W.H.; Pusparajah, P.; Chuah, L.H.; Chuah, L.H.; Ming, L.C.; Khan, T.M.; Lee, L.H.; et al. Resveratrol—Potential Antibacterial Agent against Foodborne Pathogens. Front. Pharmacol. 2018, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Singkham-In, U.; Higgins, P.G.; Wannigama, D.L.; Hongsing, P.; Chatsuwan, T. Rescued chlorhexidine activity by resveratrol against carbapenem-resistant Acinetobacter baumannii via down-regulation of AdeB efflux pump. PLoS ONE 2020, 15, e0243082. [Google Scholar] [CrossRef]
- Lacour, J.P.; Castanet, J.; Boutté, P.; Ortonne, J.P. Antiseptic treatment of the umbilical cord in newborns: Survey and recommendations. Arch. Pediatr. 1999, 6, 631–634. [Google Scholar] [CrossRef]
- Mimoz, O.; Villeminey, S.; Ragot, S.; Dahyot-Fizelier, C.; Laksiri, L.; Petitpas, F.; Debaene, B. Chlorhexidine-based antiseptic solution vs alcohol-based povidone-iodine for central venous catheter care. Arch. Intern. Med. 2007, 167, 2066–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornschuh, M.; Zwicker, P.; Kramer, A.; Schaufler, K.; Heiden, S.; Bohnert, J.; Becker, K.; Hübner, N.-O. Extensively-drug-resistant Klebsiella pneumoniae ST307 outbreak strain from north-eastern Germany does not show increased tolerance to quaternary ammonium compounds and chlorhexidine. J. Hosp. Infect. 2021, 113, 52–58. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (Eucast) of the European Society of Clinical Microbiology and Infectious Diseases (Escmid). Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2020, 6, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clin Lab Stand Institute: Wayne, PA, USA, 2019. [Google Scholar]
- EUCAST Definitive Document EDef 7.1. Method for the determination of broth dilution MICs of antifungal agents for fermentative yeasts. Clin. Microbiol. Infect. 2008, 14, 398–405. [CrossRef]
- Hall, M.; Middleton, R.; Westmacott, D. The fractional inhibitory concentration (FIC) index as a measure of synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
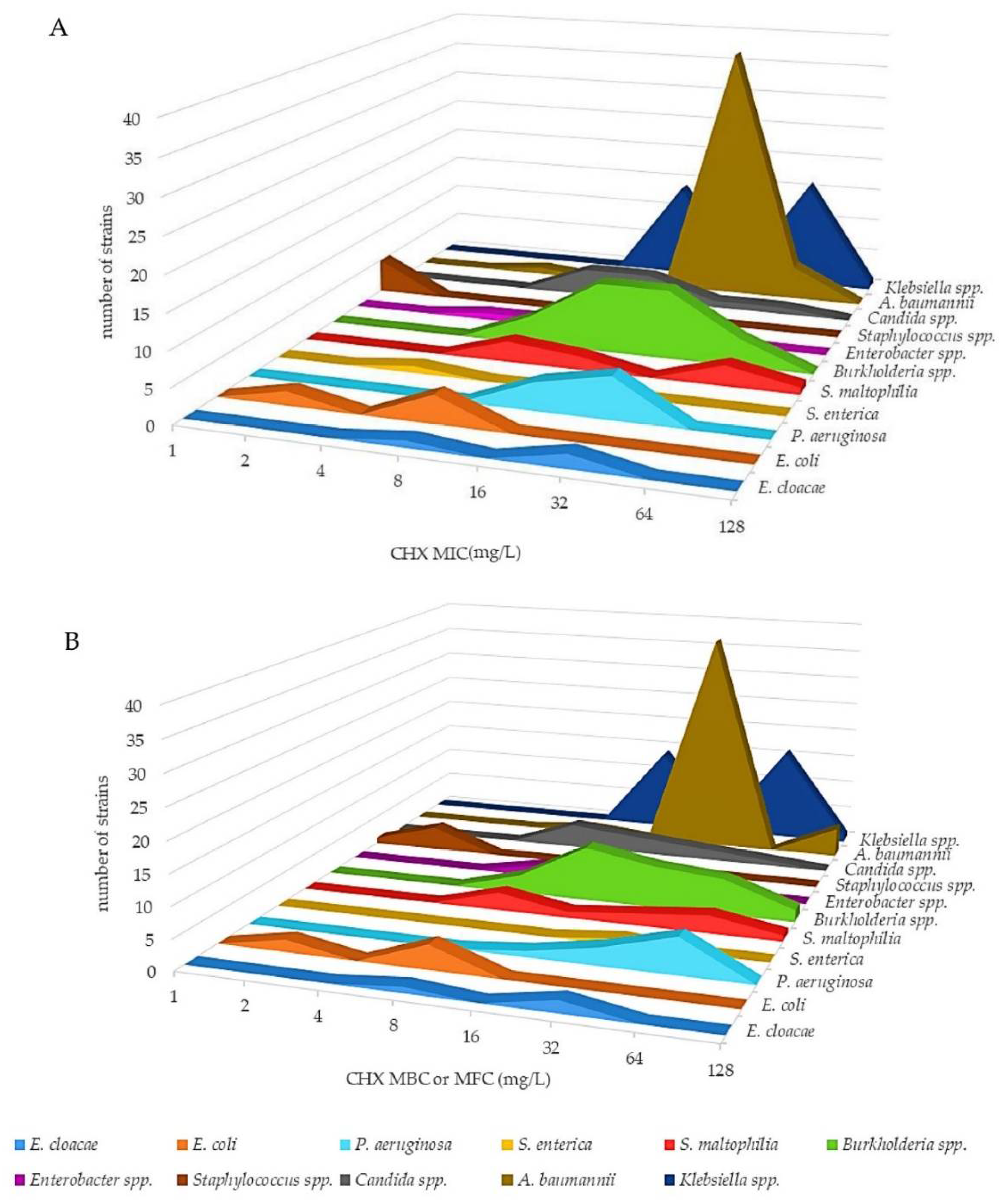


{kind=link}
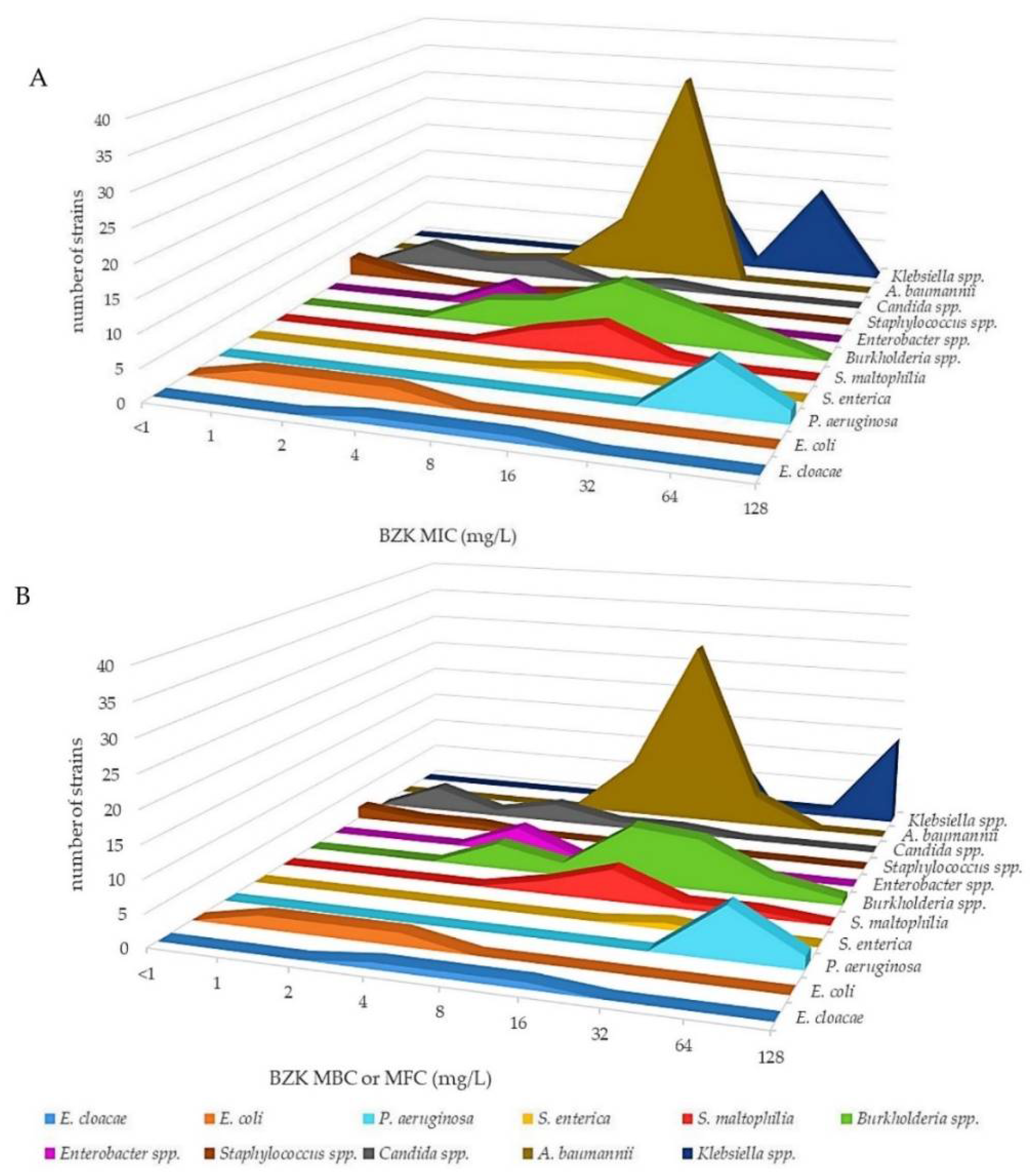{kind=link}
| Strain | CHX MIC (MBC or MFC) | BZK MIC (MBC or MFC) | ||||||
|---|---|---|---|---|---|---|---|---|
| CCCP | CCCP | |||||||
| 0 | 1 | 2 | 4 | 0 | 1 | 2 | 4 | |
| A. baumannii ATCC 19606 | 32 (32) | 8 (8) | 4 (4) | 1 (1) | 16 (32) | 8 (16) | 8 (16) | 8 (16) |
| A. baumannii ∆adeB ATCC 19606 | 4 (4) | 2 (2) | 2 (2) | 2 (2) | 8 (8) | 4 (4) | 4 (4) | 2 (2) |
| A. baumannii ACICU | 64 (128) | 32 (64) | 8 (16) | 8 (16) | 16 (16) | 16 (16) | 16 (16) | 16 (16) |
| A. baumannii AYE | 32 (32) | 8 (8) | 4 (4) | 1 (1) | 16 (32) | 8 (16) | 8 (16) | 8 (16) |
| B. cenocepacia LMG 16654 | 64 (64) | 64 (64) | 32 (32) | 16 (16) | 64 (128) | 64 (128) | 64 (128) | 64 (128) |
| B.cepacia LMG 2161 | 32 (64) | 32 (32) | 8 (8) | 4 (4) | 32 (32) | 32 (32) | 16 (32) | 16 (32) |
| B. vietnamiensis LMG 22486 | 32 (32) | 32 (32) | 8 (8) | 4 (4) | 32 (32) | 32 (32) | 16 (32) | 16 (32) |
| B. gladioli LMG 2121 | 16 (16) | 16 (32) | 8 (16) | 4 (4) | 4 (4) | 4 (4) | 1 (1) | 1 (1) |
| B. dolosa LMG 21443 | 16 (32) | 16 (32) | 8 (16) | 4 (4) | 8 (16) | 8 (16) | 4 (8) | 4 (8) |
| B. multivorans LMG 16665 | 64 (64) | 32 (32) | 16 (16) | 8 (8) | 32 (64) | 32 (32) | 16 (16) | 8 (8) |
| E. cloacae ATCC13047 | 8 (8) | 4 (8) | 2 (4) | 1 (2) | 32 (64) | 32 (64) | 32 (64) | 32 (64) |
| E. cloacae EnC-Na-1 | 32 (32) | 16 (16) | 4 (8) | 2 (4) | 4 (4) | 4 (8) | 4 (8) | 4 (8) |
| K. aerogenes ATCC 13048 | 32 (32) | 8 (16) | 4 (4) | 2 (2) | 32 (32) | 16 (16) | 16 (16) | 16 (16) |
| K. pneumoniae ATCC 700603 | 128 (128) | 32 (32) | 8 (8) | 4 (4) | 32 (64) | 32 (64) | 32 (64) | 32 (64) |
| K. pneumoniae KP-Mo-7 | 64 (64) | 16 (16) | 4 (4) | 1 (1) | 32 (32) | 32 (32) | 32 (32) | 32 (32) |
| K. pneumoniae KP-Mo-6 | 64 (64) | 16 (16) | 4 (4) | 1 (1) | 32 (32) | 32 (32) | 32 (32) | 32 (32) |
| P. aeruginosa RP73 | 32 (32) | 32 (32) | 16 (16) | 16 (16) | 64 (128) | 64 (64) | 64 (64) | 64 (64) |
| P. aeruginosa PAO1 | 16 (16) | 16 (16) | 8 (8) | 8 (8) | 64 (128) | 64 (128) | 64 (128) | 64 (64) |
| P. aeruginosa PA14 | 16 (32) | 16 (16) | 8 (8) | 8 (8) | 64 (64) | 64 (64) | 64 (64) | 64 (64) |
| P. aeruginosa PA-Na-1 | 32 (64) | 32 (32) | 16 (16) | 16 (16) | 64 (128) | 64 (64) | 64 (64) | 64 (64) |
| S. enterica ATCC 13076 | 4 (4) | 1 (1) | 0.5 (1) | 0.5 (1) | 16 (16) | 16 (16) | 16 (16) | 4 (4) |
| S. maltophilia K279 | 32 (32) | 8 (8) | 2 (2) | 2 (2) | 16 (16) | 16 (16) | 16 (16) | 16 (16) |
| S. maltophilia LMG 10853 | 8 (8) | 4 (4) | 2 (2) | 0.5 (1) | 8 (8) | 4 (8) | 4 (8) | 4 (8) |
| S. maltophilia OBGTC20 | 64 (64) | 32 (32) | 8 (8) | 2 (2) | 32 (32) | 16 (16) | 16 (16) | 16 (16) |
| E. faecalis ATCC 29212 | 64 (64) | 16 (16) | 16 (16) | 8 (8) | 4 (8) | 4 (8) | 4 (8) | 4 (8) |
| E. faecium ATCC 6057 | 8 (8) | 4 (4) | 2 (2) | 0.5 (1) | 4 (8) | 4 (8) | 4 (8) | 2 (4) |
| S. aureus ATCC 43300 | 1 (2) | <1 (<1) | <1 (<1) | <1 (<1) | 8 (8) | <1 (<1) | <1 (<1) | <1 (<1) |
| C. albicans ATCC 10231 | 16 (16) | 16 (16) | 16 (16) | 8 (8) | 2 (2) | <1 (<1) | <1 (<1) | <1 (<1) |
| C. albicans 62033 | 16 (16) | 16 (16) | 16 (16) | 8 (8) | 4 (4) | 4 (4) | 1 (1) | 1 (1) |
| C. parapsilosis 4609 | 32 (32) | 16 (16) | 16 (16) | 4 (4) | 2 (2) | <1 (<1) | <1 (<1) | <1 (<1) |
| C. krusei 81667 | 8 (8) | 8 (8) | 8 (8) | 8 (8) | 1 (1) | <1 (<1) | <1 (<1) | <1 (<1) |
| C. glabrata 61112 | 16 (16) | 16 (16) | 16 (16) | 8 (8) | 2 (2) | <1 (<1) | <1 (<1) | <1 (<1) |
| C. tropicalis 61220 | 8 (16) | 8 (8) | 8 (8) | 8 (8) | 1 (1) | <1 (<1) | <1 (<1) | <1 (<1) |
| Strain | CHX MIC (MBC or MFC) | ||||
|---|---|---|---|---|---|
| RV | |||||
| 0 | 32 | 64 | 128 | 256 | |
| A. baumannii ATCC 19606 | 32 (32) | 8 (16) | 4 (8) | 0.5 (2) | 0.125 (0.125) |
| A. baumannii ∆adeB ATCC 19606 | 4 (4) | 4 (4) | 4 (4) | 0.5 (0.5) | 0.125 (0.125) |
| A. baumannii ACICU | 64 (128) | 16 (32) | 4 (8) | 0.5 (0.5) | 0.5 (0.5) |
| A. baumannii AYE | 32 (32) | 4 (8) | 2 (4) | 0.5 (2) | 0.125 (0.125) |
| B. cenocepacia LMG 16654 | 64 (64) | 32 (32) | 32 (32) | 4 (4) | 2 (2) |
| B. cepacia LMG 2161 | 32 (32) | 16 (16) | 16 (16) | 2 (4) | 2 (2) |
| B. vietnamiensis LMG 22486 | 32 (32) | 32 (32) | 16 (16) | 2 (2) | 0.5 (0.5) |
| B. gladioli LMG 2121 | 16 (16) | 4 (4) | 4 (4) | 4 (4) | 1 (1) |
| B. dolosa LMG 21443 | 16 (16) | 8 (8) | 8 (4) | 4 (4) | 2 (2) |
| B. multivorans LMG 16665 | 64 (64) | 64 (64) | 32 (32) | 4 (8) | 2 (2) |
| E. cloacae ATCC 13047 | 8 (8) | 4 (8) | 2 (2) | 0.5 (1) | 0.5 (0.5) |
| E. cloacae EnC-Na-1 | 32 (32) | 8 (8) | 4 (8) | 4 (4) | 2 (2) |
| K. aerogenes ATCC 13048 | 32 (32) | 8 (16) | 8 (16) | 2 (2) | 1 (1) |
| K. pneumoniae ATCC 700603 | 128 (128) | 64 (64) | 32 (32) | 8 (8) | 4 (4) |
| K. pneumoniae KP-Mo-7 | 64 (64) | 32 (32) | 32 (32) | 4 (8) | 2 (4) |
| K. pneumoniae KP-Mo-6 | 64 (64) | 16 (32) | 16 (16) | 8 (16) | 2 (2) |
| P. aeruginosa RP73 | 32 (32) | 8 (16) | 8 (16) | 8 (16) | 4 (8) |
| P. aeruginosa PAO1 | 16 (16) | 4 (8) | 4 (8) | 4 (4) | 4 (4) |
| P. aeruginosa PA14 | 16 (32) | 16 (32) | 8 (32) | 4 (8) | 4 (8) |
| P. aeruginosa PA-Na-1 | 32 (64) | 8 (16) | 4 (16) | 4 (8) | 4 (8) |
| S. enterica ATCC 13076 | 4 (4) | 1 (1) | 0.5 (1) | 0.5 (1) | 0.5 (1) |
| S. maltophilia K279 | 64 (128) | 32 (32) | 16 (16) | 4 (8) | 0.5 (1) |
| S. maltophilia LMG 10853 | 8 (8) | 2 (2) | 0.5 (1) | 0.5 (1) | 0.5 (0.5) |
| S. maltophilia OBGTC20 | 64 (64) | 16 (16) | 16 (16) | 4 (4) | 1 (1) |
| E. faecalis ATCC 29212 | 32 (32) | 16 (16) | 4 (8) | 2 (2) | 0.5 (0.5) |
| E. faecium ATCC 6057 | 8 (8) | 8 (8) | 4 (4) | 2 (2) | 0.5 (0.5) |
| S. aureus ATCC 43300 | 1 (2) | 0.25 (0.25) | 0.25 (0.25) | 0.125 (0.25) | 0.125 (0.25) |
| C. albicans ATCC 10231 | 16 (32) | 16 (32) | 16 (16) | 8 (8) | 8 (8) |
| C. albicans 62033 | 16 (16) | 16 (16) | 16 (16) | 16 (16) | 8 (8) |
| C. parapsilosis 4609 | 64 (64) | 32 (64) | 16 (32) | 16 (32) | 8 (8) |
| C. krusei 81667 | 8 (8) | 4 (8) | 4 (4) | 2 (4) | 2 (4) |
| C. glabrata 61112 | 16 (16) | 8 (16) | 8 (16) | 8 (8) | 8 (8) |
| C. tropicalis 61220 | 8 (16) | 8 (8) | 8 (8) | 4 (4) | 4 (4) |
| Strain | BZK MIC (MBC or MFC) | |||
|---|---|---|---|---|
| RV | ||||
| 0 | 32 | 64 | 128 | |
| A. baumannii ATCC 19606 | 16 (32) | 8 (16) | 4 (16) | 0.5 (1) |
| A. baumannii ∆adeB ATCC 19606 | 8 (8) | 2 (4) | 1 (1) | 0.25 (0.5) |
| A. baumannii ACICU | 16 (32) | 2 (4) | 1 (2) | 0.5 (1) |
| A. baumannii AYE | 8 (8) | 1 (2) | 1 (1) | 0.25 (1) |
| B. cenocepacia LMG 16654 | 64 (64) | 32 (32) | 32 (32) | 4 (4) |
| B.cepacia LMG 2161 | 32 (32) | 16 (16) | 16 (16) | 2 (4) |
| B. vietnamiensis LMG 22486 | 32 (32) | 32 (32) | 16 (16) | 2 (2) |
| B. gladioli LMG2121 | 4 (4) | 2 (2) | 0.5 (1) | 0.5 (1) |
| B. dolosa LMG21443 | 32 (32) | 16 (16) | 16 (16) | 8 (8) |
| B. multivorans LMG 16665 | 64 (64) | 64 (64) | 32 (32) | 4 (4) |
| E. cloacae ATCC 13047 | 32 (32) | 16 (32) | 16 (16) | 16 (16) |
| E. cloacae EnC-Na-1 | 4 (4) | 4 (4) | 2 (2) | 2 (2) |
| K. aerogenes ATCC 13048 | 8 (8) | 8 (8) | 4 (4) | 2 (2) |
| K. pneumoniae ATCC 700603 | 32 (32) | 32 (32) | 32 (32) | 16 (16) |
| K. pneumoniae KP-Mo-7 | 16 (16) | 8 (16) | 4 (8) | 2 (2) |
| K. pneumoniae KP-Mo-6 | 16 (16) | 16 (16) | 4 (8) | 2 (2) |
| P. aeruginosa RP73 | 32 (32) | 32 (32) | 32 (32) | 16 (16) |
| P. aeruginosa PAO1 | 64 (64) | 64 (64) | 64 (64) | 32 (32) |
| P. aeruginosa PA14 | 64 (64) | 32 (64) | 32 (64) | 32 (64) |
| P. aeruginosa PA-Na-1 | 64 (128) | 64 (128) | 64 (64) | 64 (64) |
| S. enterica ATCC 13076 | 32 (32) | 32 (32) | 16 (16) | 8 (8) |
| S. maltophilia K279 | 16 (16) | 16 (16) | 8 (16) | 4 (8) |
| S. maltophilia LMG 10853 | 8 (8) | 1 (1) | 0.5 (1) | 0.5 (1) |
| S. maltophilia OBGTC20 | 32 (32) | 8 (16) | 4 (8) | 2 (2) |
| E. faecalis ATCC 29212 | 4 (8) | 4 (8) | 4 (8) | 2 (2) |
| E. faecium ATCC 6057 | 4 (4) | 4 (4) | 1 (1) | 0.5 (1) |
| S. aureus ATCC 43300 | 8 (16) | 0.5 (0.5) | 0.25 (0.25) | 0.125 (0.25) |
| C. albicans ATCC 10231 | 4 (4) | 4 (4) | 2 (2) | 2 (2) |
| C. albicans 62033 | 4 (4) | 4 (4) | 2 (2) | 2 (2) |
| C. parapsilosis 4609 | 4 (8) | 4 (4) | 2 (2) | 2 (2) |
| C. krusei 81667 | 1 (1) | 0.5 (1) | 0.5 (1) | 0.5 (1) |
| C. glabrata 61112 | 2 (2) | 0.5 (1) | 0.5 (1) | 0.5 (1) |
| C. tropicalis 61220 | 1 (1) | 0.5 (1) | 0.125 (0.125) | 0.125 (0.125) |
| Strain | CHX MIC (MBC or MFC) | BZK MIC (MBC or MFC) | 0 RV | FIC * Index (a) | 32 RV | FIC * Index (b) | 64 RV | FIC * Index (c) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CHX + BZK MIC (MBC or MFC) | CHX + BZK MIC (MBC or MFC) | CHX + BZK MIC (MBC or MFC) | |||||||||
| A. baumannii ATCC 19606 | 32 (32) | 16 (16) | 8 (8) | 4 (4) | 0.5 | 2 (2) | 2 (2) | 0.187 | 2 (2) | 0.5 (0.5) | 0.093 |
| A. baumannii ∆adeB ATCC 19606 | 4 (4) | 8 (8) | 1 (1) | 2 (2) | 0.5 | 2 (2) | 0.06 (0.06) | 0.50 | 0.5 (0.5) | 0.5 (0.5) | 0.18 |
| A. baumannii ACICU | 64 (128) | 16 (32) | 8 (8) | 2 (2) | 0.25 | 2 (2) | 0.5 (0.5) | 0.062 | 2 (2) | 0.5 (0.5) | 0.062 |
| B. cenocepacia LMG 16654 | 64 (64) | 64 (64) | 8 (8) | 2 (2) | 0.15 | 8 (8) | 2 (2) | 0.15 | 8 (8) | 0.5 (0.5) | 0.13 |
| B. dolosa LMG 21443 | 16 (16) | 8 (8) | 4 (8) | 2 (2) | 0.31 | 4 (4) | 1 (1) | 0.28 | 4 (4) | 0.5 (0.5) | 0.26 |
| B. multivorans LMG16665 | 64 (64) | 32 (64) | 8 (8) | 8 (8) | 0.25 | 8 (8) | 8 (8) | 0.25 | 8 (8) | 0.5 (0.5) | 0.13 |
| E. cloacae ATCC 13047 | 8 (8) | 8 (8) | 2 (2) | 4 (4) | 0.5 | 2 (2) | 4 (4) | 0.5 | 2 (2) | 2 (2) | 0.375 |
| K. pneumoniae ATCC 700603 | 128 (128) | 16 (16) | 16 (32) | 4 (4) | 0.25 | 4 (4) | 2 (2) | 0.093 | 4 (4) | 2 (2) | 0.093 |
| K. pneumoniae kp-Mo-7 | 64 (64) | 16 (16) | 2 (2) | 2 (2) | 0.156 | 2 (2) | 1 (1) | 0.092 | 1 (1) | 0.5 (0.5) | 0.062 |
| P. aeruginosa RP73 | 32 (32) | 64 (64) | 8 (16) | 4 (4) | 0.31 | 4 (8) | 8 (8) | 0.25 | 4 (4) | 1 (1) | 0.14 |
| P. aeruginosa PAO1 | 16 (16) | 64 (128) | 16 (32) | 2 (2) | 1.03 | 4 (4) | 2 (2) | 0.28 | 4 (4) | 0.5 (0.5) | 0.25 |
| P. aeruginosa PA14 | 16 (32) | 64 (64) | 4 (8) | 4 (4) | 0.31 | 4 (4) | 4 (4) | 0.31 | 4 (4) | 1 (1) | 0.26 |
| P. aeruginosa PA-Na-1 | 32 (64) | 64 (128) | 8 (8) | 2 (2) | 0.28 | 4 (4) | 2 (2) | 0.156 | 4 (4) | 0.5 (0.5) | 0.132 |
| S. enterica ATCC 13076 | 4 (4) | 16 (32) | 1 (1) | 1 (1) | 0.312 | 1 (1) | 1 (1) | 0.312 | 0.5 (0.5) | 0.5 (0.5) | 0.15 |
| S. maltophilia K279 | 128 (128) | 16 (16) | 16 (32) | 8 (8) | 0.62 | 4 (4) | 4 (8) | 0.281 | 4 (4) | 2 (2) | 0.156 |
| E. faecalis ATCC 29212 | 32 (32) | 4 (8) | 4 (4) | 2 (2) | 0.625 | 0.5 (0.5) | 0.5 (0.5) | 0.14 | 0.5 (0.5) | 0.5 (0.5) | 0.14 |
| E. faecium ATCC 6057 | 8 (8) | 4 (4) | 0.125 (0.125) | 0.5 (0.5) | 0.14 | 0.5 (0.5) | 0.125 (0.125) | 0.093 | 0.25 (0.25) | 0.06 (0.06) | 0.046 |
| S. aureus ATCC43300 | 1 (2) | 8 (16) | 0.125 (0.125) | 0.5 (0.5) | 0.187 | 0.25 (0.25) | 0.06 (0.06) | 0.257 | 0.125 (0.125) | 0.25 (0.25) | 0.156 |
| C. albicans ATCC 10231 | 16 (32) | 4 (4) | 8 (8) | 1 (1) | 0.75 | 2 (4) | 0.125 (0.125) | 0.156 | 2 (2) | 0.125 (0.0125) | 0.156 |
| C. albicans 62033 | 16 (16) | 2 (2) | 8 (16) | 2 (2) | 1 | 4 (4) | 2 (2) | 0.75 | 4 (4) | 2 (2) | 0.75 |
| C. parapsilosis 4609 | 64 (64) | 2 (2) | 8 (16) | 2 (2) | 0.51 | 1 (1) | 2 (2) | 0.26 | 1 (1) | 2 (2) | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliaccio, A.; Stabile, M.; Bagattini, M.; Triassi, M.; Berisio, R.; De Gregorio, E.; Zarrilli, R. Resveratrol Reverts Tolerance and Restores Susceptibility to Chlorhexidine and Benzalkonium in Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts. Antibiotics 2022, 11, 961. https://doi.org/10.3390/antibiotics11070961
Migliaccio A, Stabile M, Bagattini M, Triassi M, Berisio R, De Gregorio E, Zarrilli R. Resveratrol Reverts Tolerance and Restores Susceptibility to Chlorhexidine and Benzalkonium in Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts. Antibiotics. 2022; 11(7):961. https://doi.org/10.3390/antibiotics11070961
Chicago/Turabian StyleMigliaccio, Antonella, Maria Stabile, Maria Bagattini, Maria Triassi, Rita Berisio, Eliana De Gregorio, and Raffaele Zarrilli. 2022. "Resveratrol Reverts Tolerance and Restores Susceptibility to Chlorhexidine and Benzalkonium in Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts" Antibiotics 11, no. 7: 961. https://doi.org/10.3390/antibiotics11070961
APA StyleMigliaccio, A., Stabile, M., Bagattini, M., Triassi, M., Berisio, R., De Gregorio, E., & Zarrilli, R. (2022). Resveratrol Reverts Tolerance and Restores Susceptibility to Chlorhexidine and Benzalkonium in Gram-Negative Bacteria, Gram-Positive Bacteria and Yeasts. Antibiotics, 11(7), 961. https://doi.org/10.3390/antibiotics11070961