
The RNA Polymerase Inhibitor Corallopyronin A Has a Lower Frequency of Resistance Than Rifampicin in Staphylococcus aureus

 , , , ,
, , , ,  ,
, 
Abstract
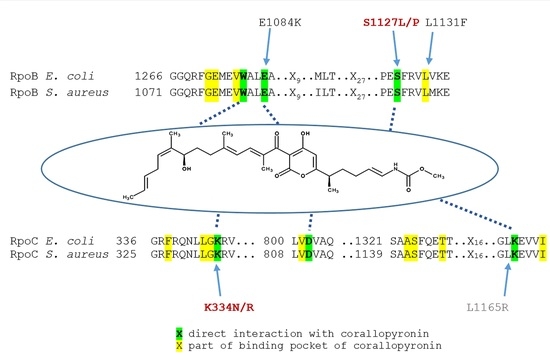
:
1. Introduction
2. Results
2.1. Stability of CorA in Agar Plates
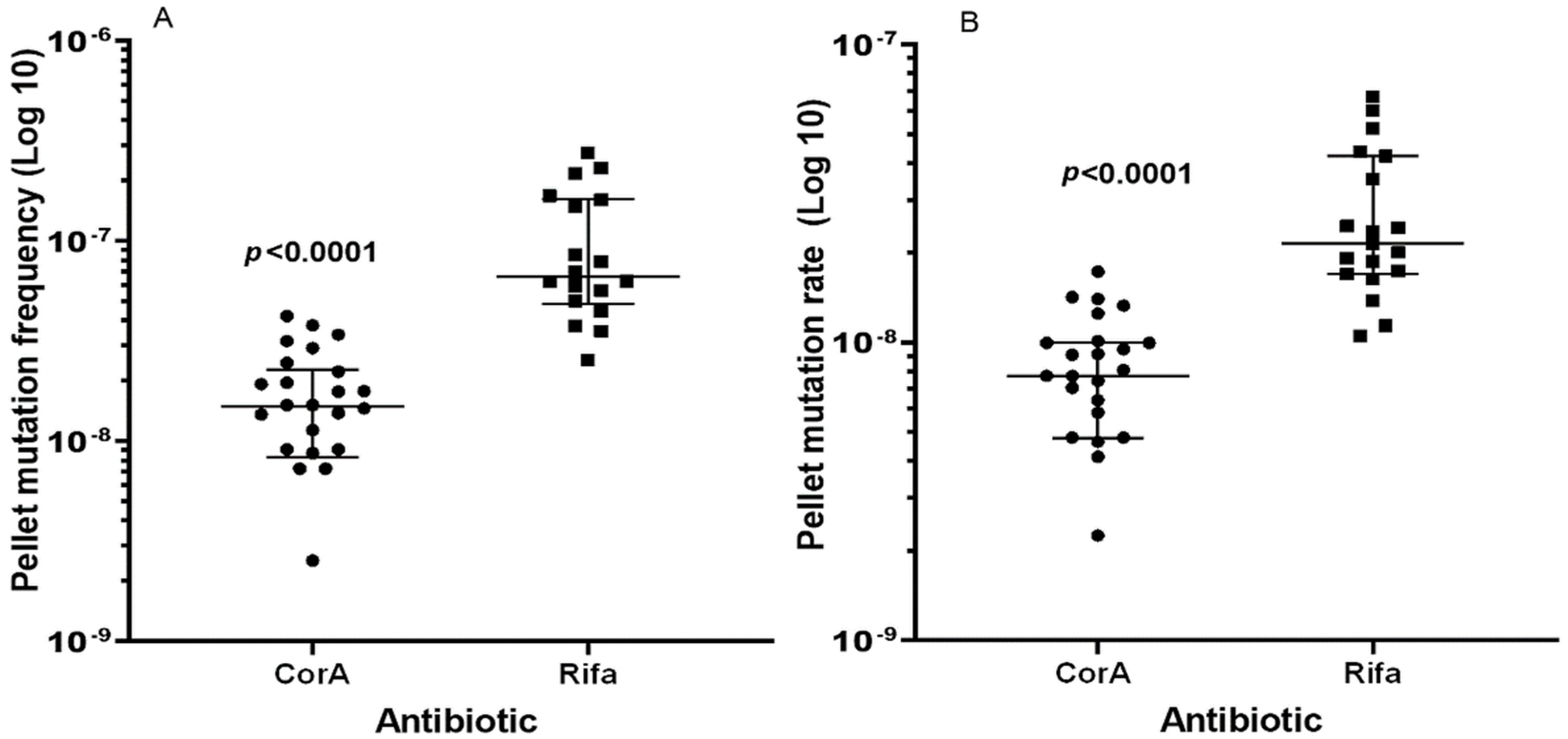
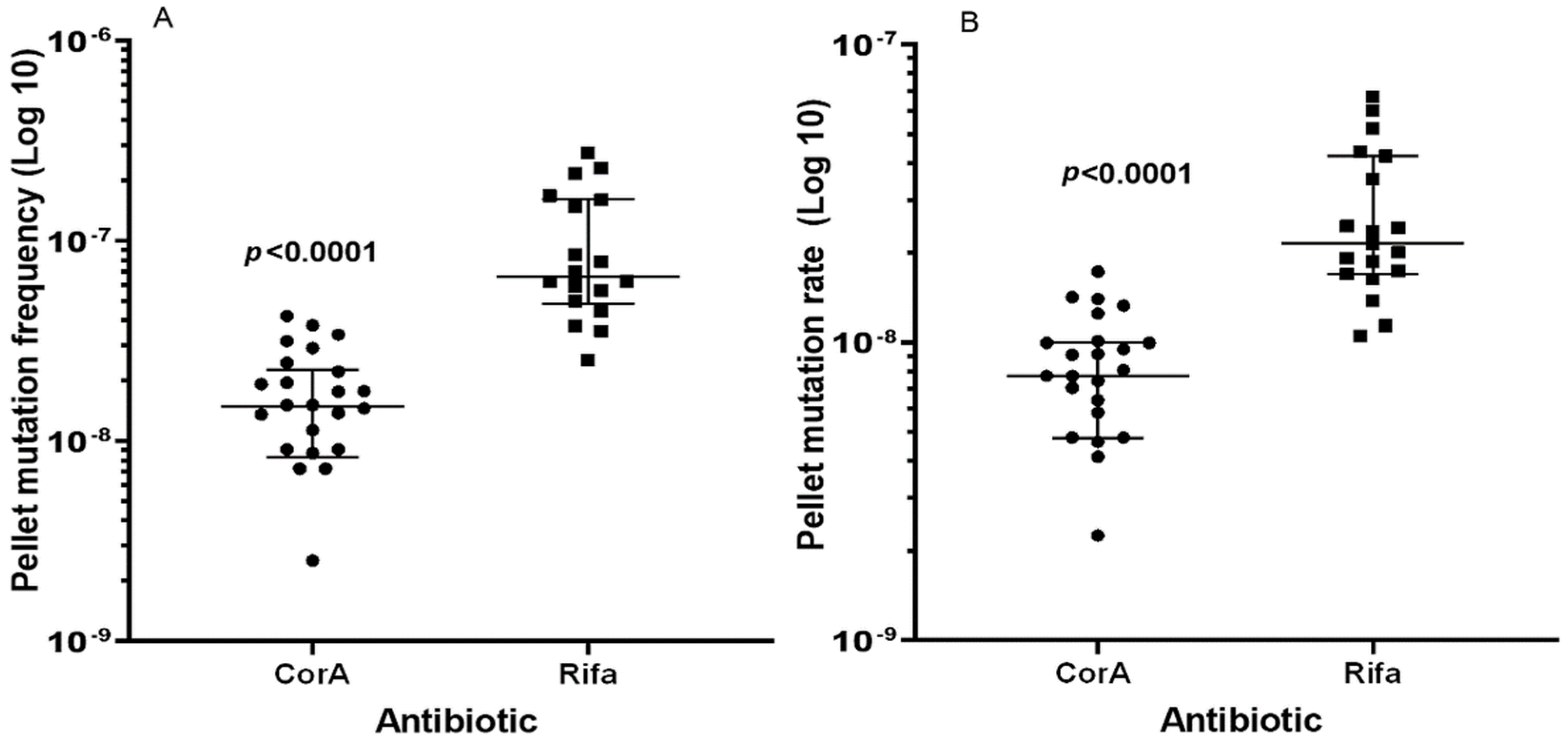
2.2. Determination of Mutation Frequency
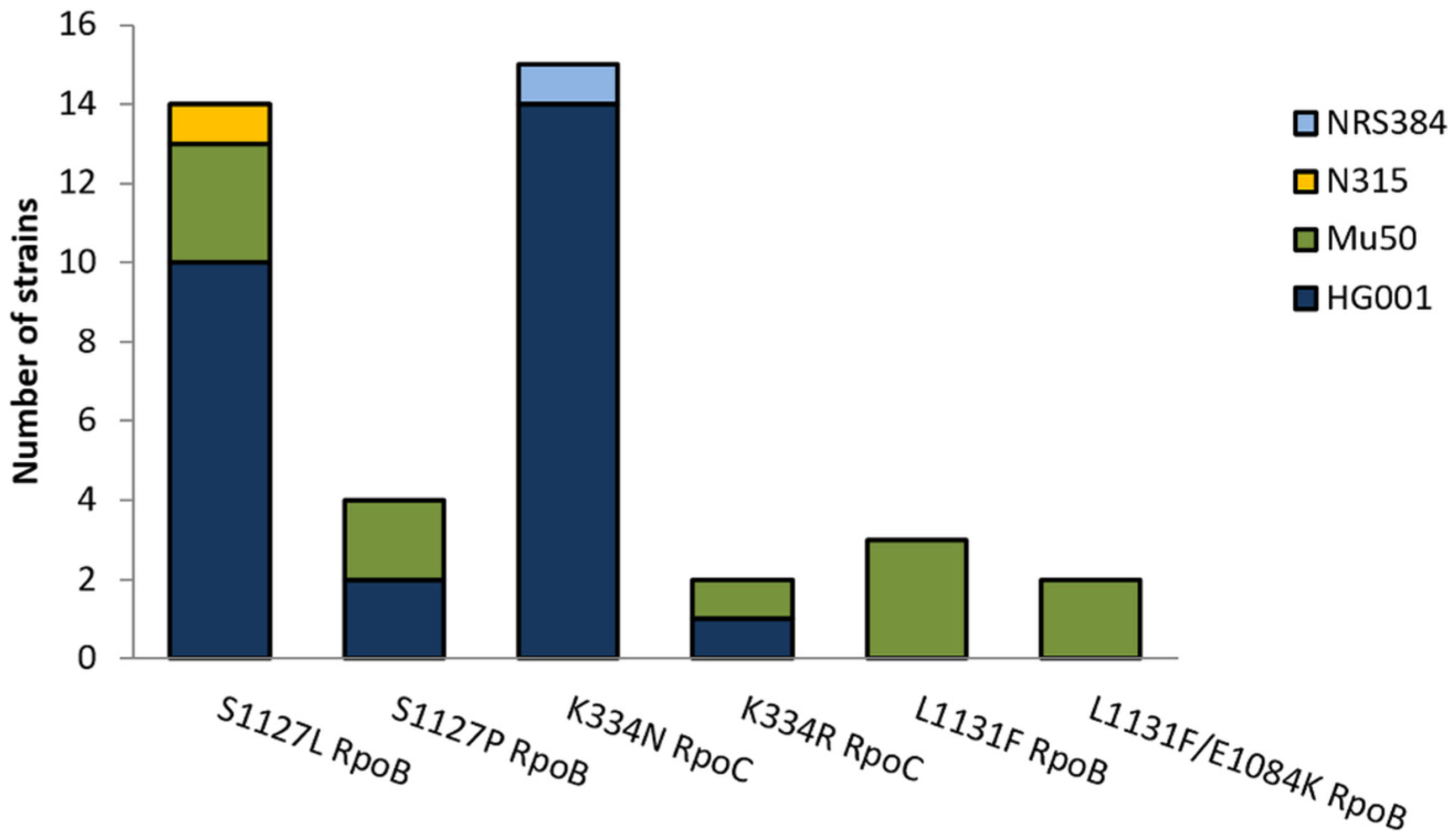
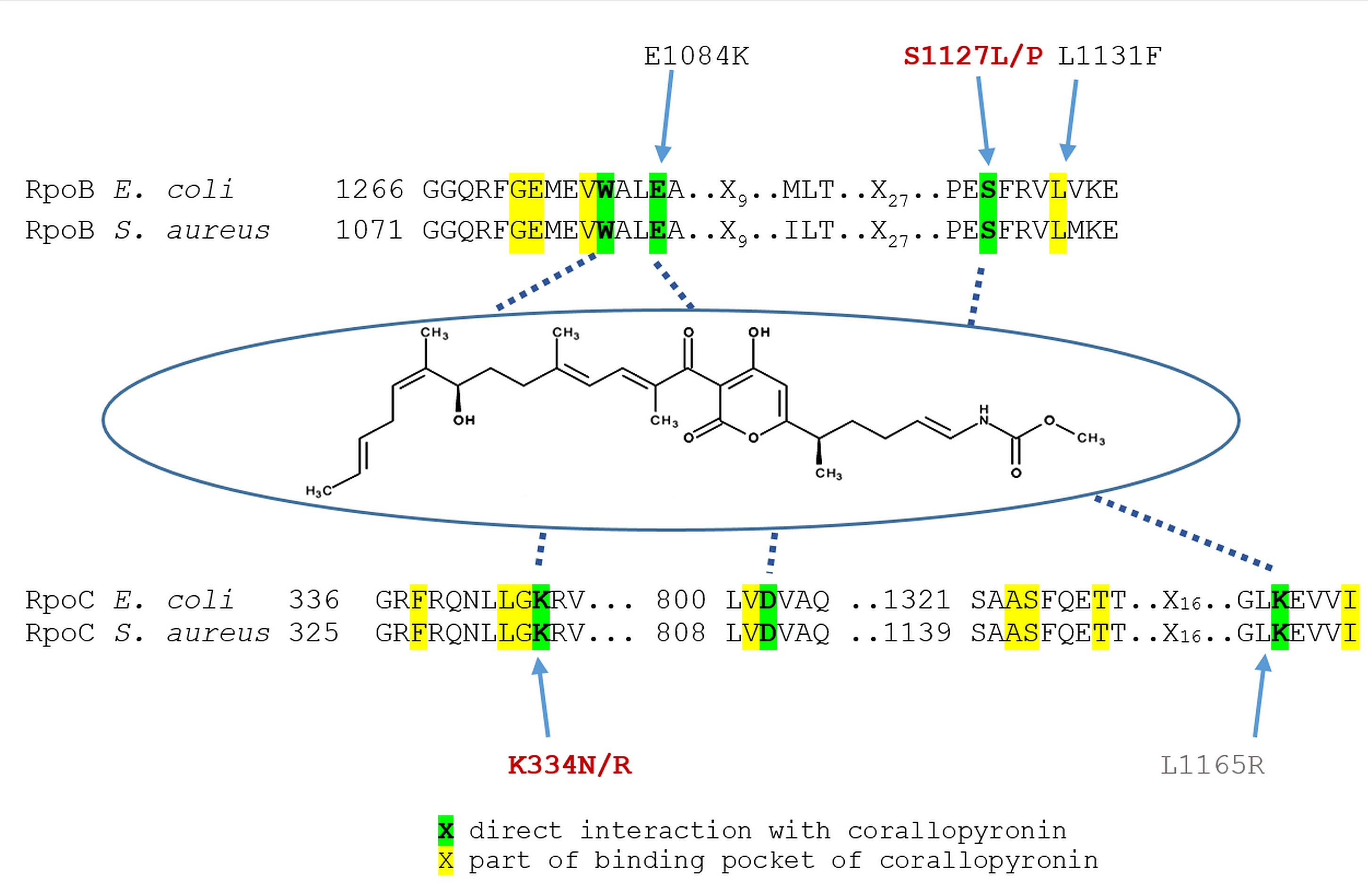
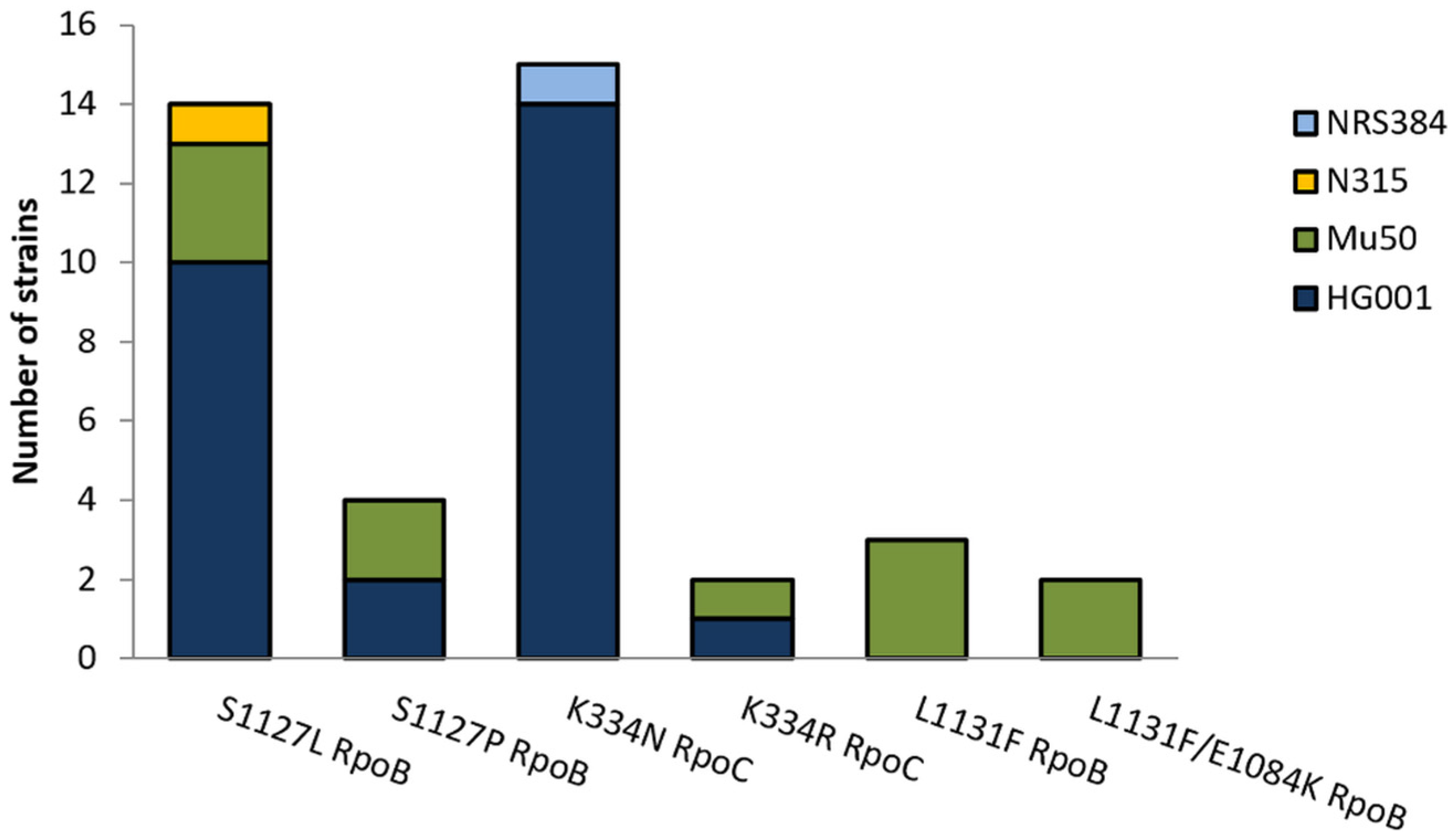
2.3. Mutations in CorA Resistant Strains
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Corallopyronin
4.3. Minimal Inhibitory Concentration
4.4. Determination of Mutation Frequency and Mutation Rates
4.5. Identification of Mutations in rpoB and rpoC
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irschik, H.; Jansen, R.; Höfle, G.; Gerth, K.; Reichenbach, H. The corallopyronins, new inhibitors of bacterial RNA synthesis from Myxobacteria. J. Antibiot. 1985, 38, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeper, N.; Graspeuntner, S.; Ledig, S.; Kaufhold, I.; Hoellen, F.; Schiefer, A.; Henrichfreise, B.; Pfarr, K.; Hoerauf, A.; Shima, K.; et al. Elaborations on Corallopyronin A as a novel treatment strategy against genital chlamydial infections. Front. Microbiol. 2019, 10, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiefer, A.; Schmitz, A.; Schäberle, T.F.; Specht, S.; Lämmer, C.; Johnston, K.L.; Vassylyev, D.G.; König, G.M.; Hoerauf, A.; Pfarr, K. Corallopyronin A specifically targets and depletes essential obligate Wolbachia endobacteria from filarial nematodes in vivo. J. Infect. Dis. 2012, 206, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kock, F.; Hauptmann, M.; Osterloh, A.; Schäberle, T.F.; Poppert, S.; Frickmann, H.; Menzel, K.-D.; Peschel, G.; Pfarr, K.; Schiefer, A.; et al. Orientia tsutsugamushi is highly susceptible to the RNA polymerase switch region inhibitor Corallopyronin A in vitro and in vivo. Antimicrob. Agents Chemother. 2018, 62, e01732-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiefer, A.; Hübner, M.P.; Krome, A.; Lämmer, C.; Ehrens, A.; Aden, T.; Koschel, M.; Neufeld, H.; Chaverra-Muñoz, L.; Jansen, R.; et al. Corallopyronin A for short-course anti-wolbachial, macrofilaricidal treatment of filarial infections. PLoS Negl. Trop. Dis. 2020, 14, e0008930. [Google Scholar] [CrossRef] [PubMed]
- Belogurov, G.A.; Vassylyeva, M.N.; Sevostyanova, A.; Appleman, J.R.; Xiang, A.X.; Lira, R.; Webber, S.E.; Klyuyev, S.; Nudler, E.; Artsimovitch, I.; et al. Transcription inactivation through local refolding of the RNA polymerase structure. Nature 2009, 457, 332–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, J.; Das, K.; Ismail, S.; Koppstein, D.; Jang, M.; Hudson, B.; Sarafianos, S.; Tuske, S.; Patel, J.; Jansen, R.; et al. The RNA polymerase “switch region” is a target for inhibitors. Cell 2008, 135, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariner, K.; McPhillie, M.; Trowbridge, R.; Smith, C.; O’Neill, A.J.; Fishwick, C.W.G.; Chopra, I. Activity of and development of resistance to corallopyronin A, an inhibitor of RNA polymerase. Antimicrob. Agents Chemother. 2011, 55, 2413–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäberle, T.F.; Schmitz, A.; Zocher, G.; Schiefer, A.; Kehraus, S.; Neu, E.; Roth, M.; Vassylyev, D.G.; Stehle, T.; Bierbaum, G.; et al. Insights into structure-activity relationships of bacterial RNA polymerase inhibiting Corallopyronin derivatives. J. Nat. Prod. 2015, 78, 2505–2509. [Google Scholar] [CrossRef] [PubMed]
- Pogorevc, D.; Panter, F.; Schillinger, C.; Jansen, R.; Wenzel, S.C.; Müller, R. Production optimization and biosynthesis revision of corallopyronin A, a potent anti-filarial antibiotic. Metab. Eng. 2019, 55, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Krome, A.K.; Becker, T.; Kehraus, S.; Schiefer, A.; Steinebach, C.; Aden, T.; Frohberger, S.J.; López Mármol, Á.; Kapote, D.; Jansen, R.; et al. Solubility and stability enhanced oral formulations for the anti-infective Corallopyronin A. Pharmaceutics 2020, 12, 1105. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.; Ziebandt, A.-K.; Ohlsen, K.; Schäfer, T.; Hecker, M.; Albrecht, D.; Novick, R.; Götz, F. Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect. Immun. 2010, 78, 2877–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, A.; Degen, D.; Ebright, Y.W.; Ebright, R.H. Frequency, spectrum, and nonzero fitness costs of resistance to myxopyronin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 6250–6255. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.M.; Ma, C.-X.; Liang, P.; Singh, K.K. Fluctuation analysis CalculatOR: A web tool for the determination of mutation rate using Luria-Delbruck fluctuation analysis. Bioinformatics 2009, 25, 1564–1565. [Google Scholar] [CrossRef] [Green Version]
- Doundoulakis, T.; Xiang, A.X.; Lira, R.; Agrios, K.A.; Webber, S.E.; Sisson, W.; Aust, R.M.; Shah, A.M.; Showalter, R.E.; Appleman, J.R.; et al. Myxopyronin B analogs as inhibitors of RNA polymerase, synthesis and biological evaluation. Bioorg. Med. Chem. Lett. 2004, 14, 5667–5672. [Google Scholar] [CrossRef]
- Lira, R.; Xiang, A.X.; Doundoulakis, T.; Biller, W.T.; Agrios, K.A.; Simonsen, K.B.; Webber, S.E.; Sisson, W.; Aust, R.M.; Shah, A.M.; et al. Syntheses of novel myxopyronin B analogs as potential inhibitors of bacterial RNA polymerase. Bioorg. Med. Chem. Lett. 2007, 17, 6797–6800. [Google Scholar] [CrossRef]
- O’Neill, A.J.; Huovinen, T.; Fishwick, C.W.G.; Chopra, I. Molecular genetic and structural modeling studies of Staphylococcus aureus RNA polymerase and the fitness of rifampin resistance genotypes in relation to clinical prevalence. Antimicrob. Agents Chemother. 2006, 50, 298–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Katayama, Y.; Sekine, M.; Hishinuma, T.; Aiba, Y.; Hiramatsu, K. Complete reconstitution of the vancomycin-intermediate Staphylococcus aureus phenotype of strain Mu50 in vancomycin-susceptible S. aureus. Antimicrob. Agents Chemother. 2016, 60, 3730–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Intrepretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 23 June 2022).
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Strain | Exchange in Resistant Mutant | CorA MIC |
|---|---|---|
| S. aureus HG001 | Wildtype | 0.25 mg/L |
| S. aureus HG001CSz1 | K334N RpoC | 64 mg/L |
| S. aureus Mu50 | Wildtype | 0.25 mg/L |
| S. aureus Mu50CSz1 | L1131F RpoB | 4 mg/L |
| S. aureus Mu50R1.1 | S1127P RpoB | 32 mg/L |
| S. aureus Mu50R3.1 | L1131F/E1084K RpoB | 128 mg/L |
| S. aureus Mu50R4.1 | S1127L RpoC | 64 mg/L |
| S. aureus Mu50R5.1 | K334R RpoC | 64 mg/L |
| S. aureus N315 | Wildtype | 0.25 mg/L |
| S. aureus N315CSz1 | S1127L RpoB | 64 mg/L |
| S. aureus USA300 | Wildtype | 0.25 mg/L |
| S. aureus USA300CSz1 | K334N RpoC | 128 mg/L |
| 1 mL Culture on CorA Agar (+/− 95% CI 1) | 1 mL Culture on Rifa Agar (+/− 95% CI) | Ratio Rifa/CorA | |
|---|---|---|---|
| Median of FoM | 1.48 × 10−8 (4.7 × 10−9, 5.7 × 10−9) | 6.67 × 10−8 (8.2 × 10−8, 1.0 × 10−8) | 4.5 |
| Mutation rate (LC) 2 | 7.72 × 10−9 (2.3 × 10−9, 2.9 × 10−9) | 2.15 × 10−8 (1.4 × 10−8, 4.1 × 10−9) | 2.8 |
| Mutation rate (MSS-MLE) 3 | 5.93 × 10−9 (2.0 × 10−9, 1.79 × 10−9) | 2.49 × 10−8 (7.3 × 10−9, 6.6 × 10−9) | 4.2 |
| 0.1 mL culture on CorA agar (+/− 95% CI) | 0.1 mL culture on rifampicin agar (+/− 95% CI) | Ratio Rifa/CorA | |
| Median of FoM | 2.27 × 10−8 (1.5 × 10−7, 2.3 × 10−7) | 1.27 × 10−7 (1.0 × 10−7, 4.0 × 10−8) | 5.6 |
| Mutation rate (LC) 2 | 8.77 × 10−9 (5.8 × 10−9, 8.8 × 10−9) | 3.49 × 10−8 (2.0 × 10−8, 9.0 × 10−9) | 4.0 |
| Mutation rate (MSS-MLE) 3 | 9.5 × 10−9 (3.1 × 10−9, 2.8 × 10−9) | 3.51 × 10−8 (8.6 × 10−9, 7.8 × 10−9) | 3.7 |
| Primer | Gene | Sequence | Ref. |
|---|---|---|---|
| rpoAI | rpoA | 5′-TAACTGCGATCAGAGACGTTACTCC-3′ | [17] |
| rpoAII | rpoA | 5′-GCTGCATTACGACGAGAAGCTAAAT-3′ | [17] |
| rpoBF | rpoB | 5′-ATTAGTGTTGCCGTTTTCTTTT-3′ | [17] |
| rpoBR | rpoB | 5′-AGTATCTTTTTGCCTGTTTTG-3′ | [17] |
| rpoCI | rpoC | 5′-GACGATGATGTTGTAGAACGCAAAG-3′ | [17] |
| rpoCII | rpoC | 5′-TGTTGTTTGTTAAAGCGTGCAACTT-3′ | [17] |
| rpoCfor2 | rpoC | 5′-GGACGATTTGCAACAAGTGA-3′ | This study |
| rpoCrev2 | rpoC | 5′-CCAACTGCTTCACCAACTTC-3′ | This study |
| F3 | rpoB | 5′-AGTCTATCACACCTCAACAA-3′ | This study |
| F4 | rpoB | 5′-TAATAGCCGCACCAGAATCA-3′ | This study |
| 2955F | rpoB | 5′-GATCGATATCATGTTAAATCCTCTTGGTGTACCATC-3′ | This study |
| 3824R | rpoB | 5′-CTATTTTCATTAATGGAAATTATTT-ACATCAATCA-3′ | This study |
| 955F | rpoB | 5′-ACTGAAACTGGTGAAATTGTAGTTG-3′ | This study |
| 2706R | rpoB | 5′-GTGAGGTACACGTAATGAAGTATCT-3′ | This study |
| 653F | rpoC | 5′-TTTTGATGTACTTCCAATCATCCCACCAGAAATTC-3′ | This study |
| 1552R | rpoC | 5′-CATATGCTTTTAATACTTCATTTGTATTATTAAAGA-3′ | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balansky, J.; Pfarr, K.; Szekat, C.; Kehraus, S.; Aden, T.; Grosse, M.; Jansen, R.; Hesterkamp, T.; Schiefer, A.; König, G.M.; et al. The RNA Polymerase Inhibitor Corallopyronin A Has a Lower Frequency of Resistance Than Rifampicin in Staphylococcus aureus. Antibiotics 2022, 11, 920. https://doi.org/10.3390/antibiotics11070920
Balansky J, Pfarr K, Szekat C, Kehraus S, Aden T, Grosse M, Jansen R, Hesterkamp T, Schiefer A, König GM, et al. The RNA Polymerase Inhibitor Corallopyronin A Has a Lower Frequency of Resistance Than Rifampicin in Staphylococcus aureus. Antibiotics. 2022; 11(7):920. https://doi.org/10.3390/antibiotics11070920
Chicago/Turabian StyleBalansky, Jan, Kenneth Pfarr, Christiane Szekat, Stefan Kehraus, Tilman Aden, Miriam Grosse, Rolf Jansen, Thomas Hesterkamp, Andrea Schiefer, Gabriele M. König, and et al. 2022. "The RNA Polymerase Inhibitor Corallopyronin A Has a Lower Frequency of Resistance Than Rifampicin in Staphylococcus aureus" Antibiotics 11, no. 7: 920. https://doi.org/10.3390/antibiotics11070920
APA StyleBalansky, J., Pfarr, K., Szekat, C., Kehraus, S., Aden, T., Grosse, M., Jansen, R., Hesterkamp, T., Schiefer, A., König, G. M., Stadler, M., Hoerauf, A., & Bierbaum, G. (2022). The RNA Polymerase Inhibitor Corallopyronin A Has a Lower Frequency of Resistance Than Rifampicin in Staphylococcus aureus. Antibiotics, 11(7), 920. https://doi.org/10.3390/antibiotics11070920