
Laboratory Diagnosis and In Vitro Antifungal Susceptibility of Trichophyton quinckeanum from Human Zoonoses and Cats




Abstract
:1. Introduction
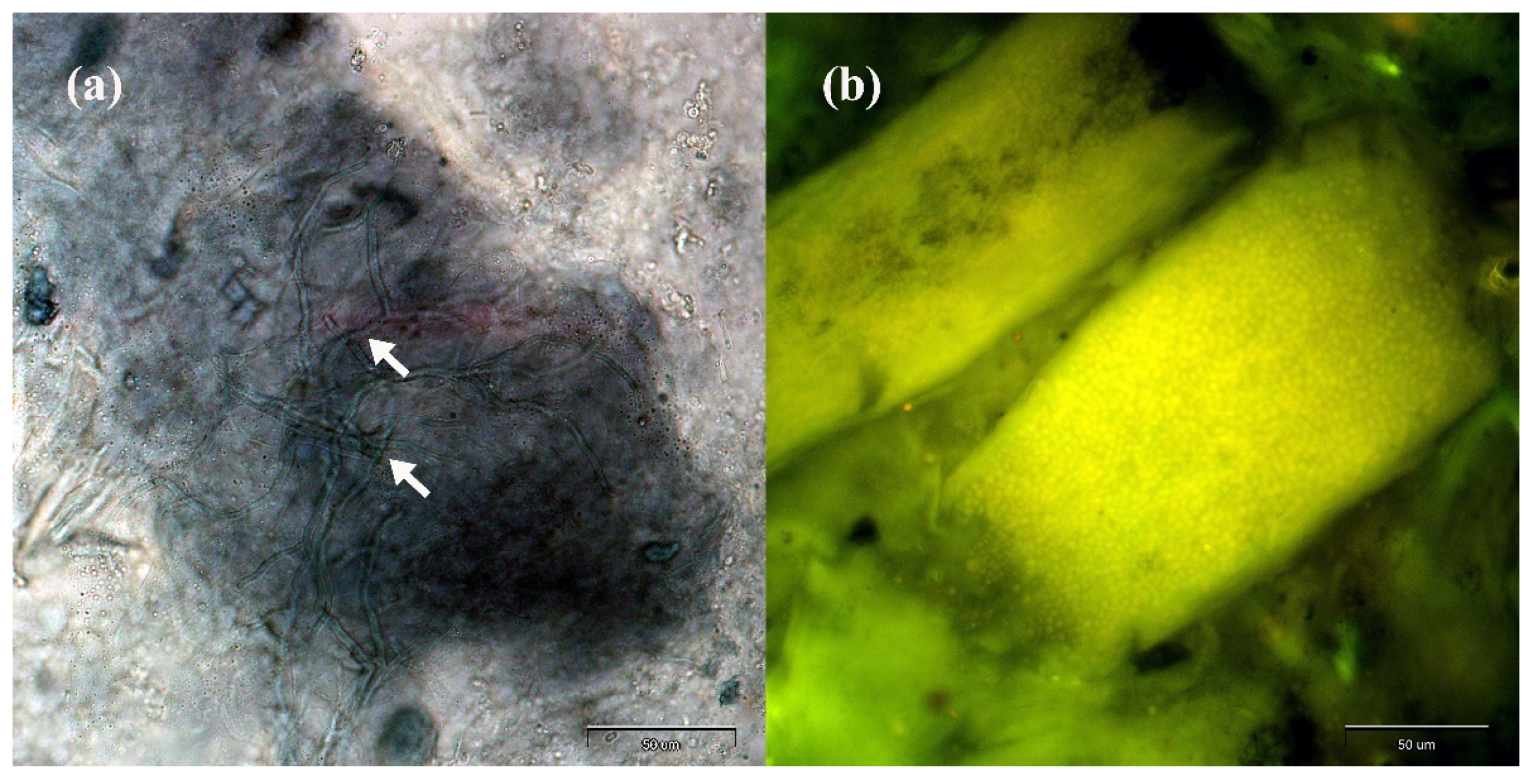
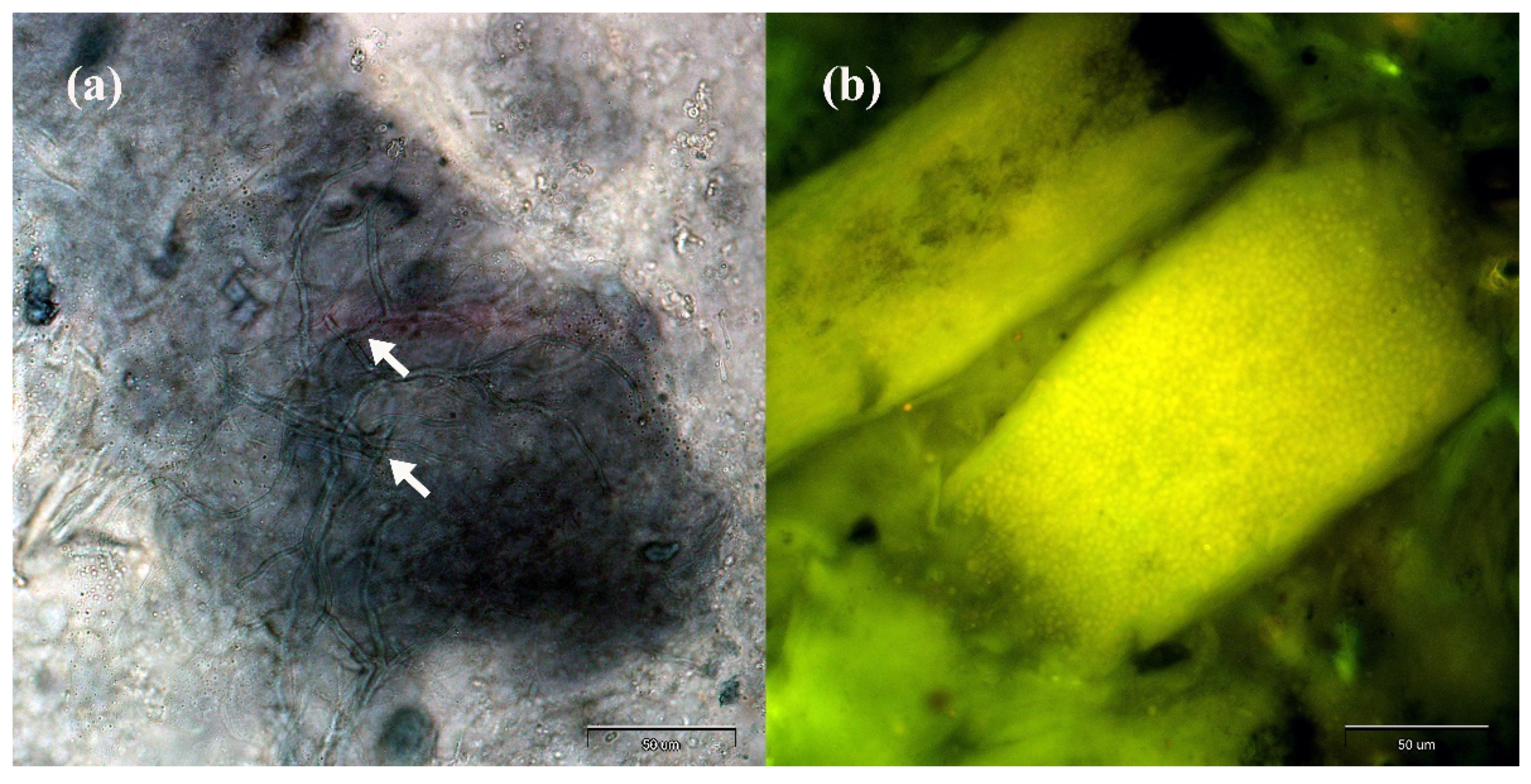
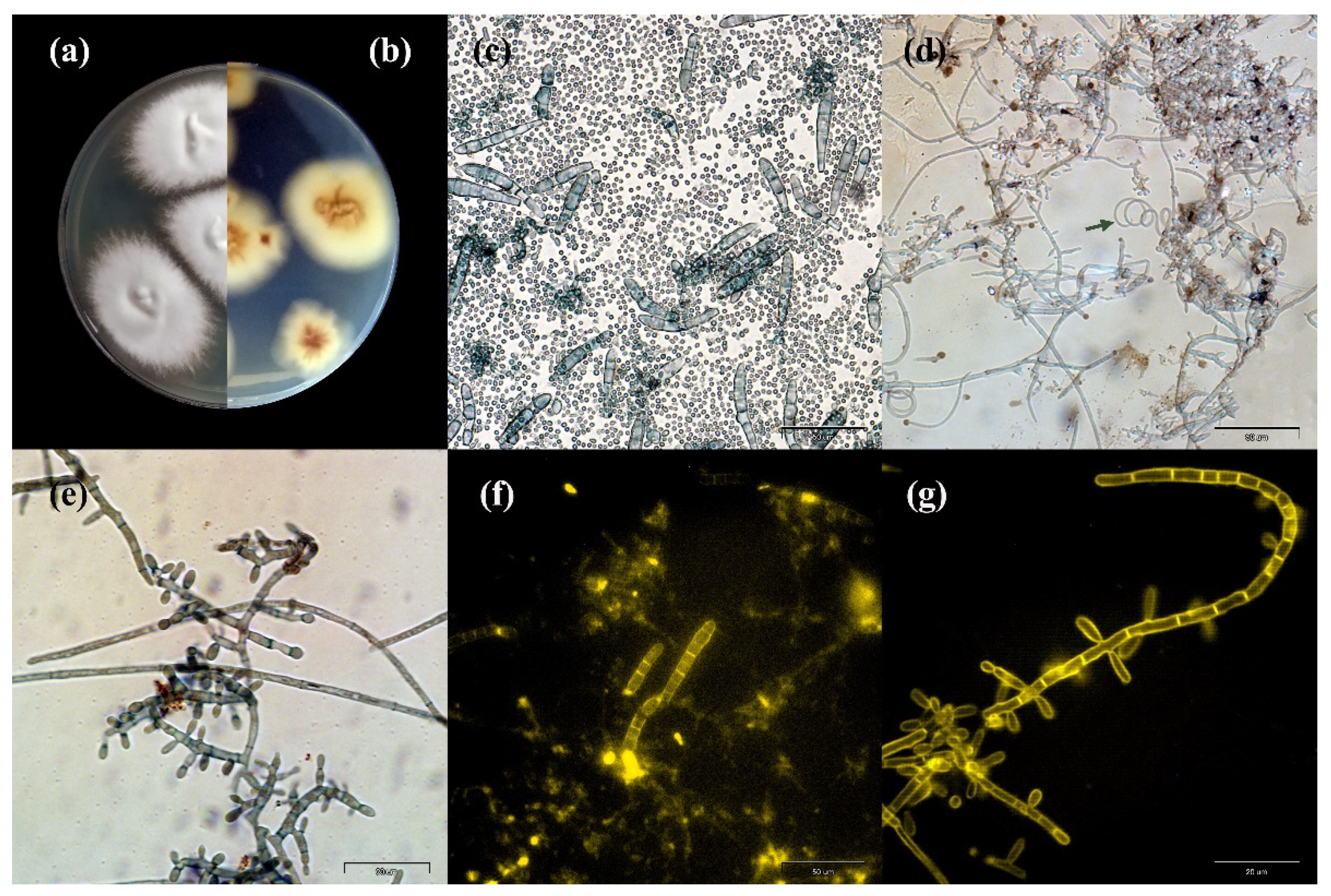
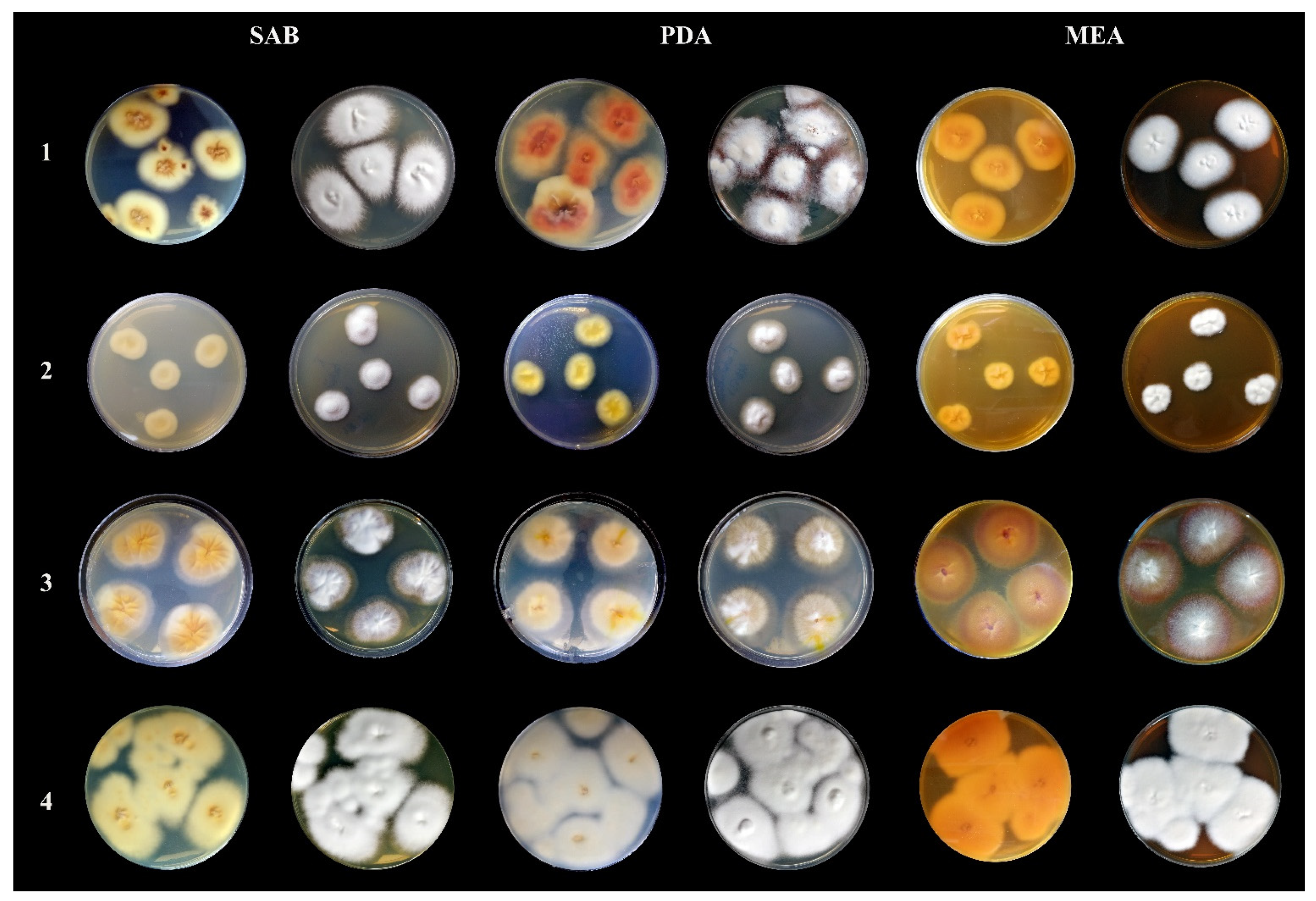
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Dermatophyte Clinical Isolates
4.3. Diagnosis Procedure
4.3.1. Direct Microscopical Examination
4.3.2. Primary Isolation on Culture
4.3.3. Molecular Identification
4.4. Antifungal Susceptibility Testing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murmu, S.; Debnath, C.; Pramanik, A.K.; Mitra, T.; Jana, S.; Dey, S.; Banerjee, S.; Batabyal, K. Detection and Characterization of Zoonotic Dermatophytes from Dogs and Cats in and around Kolkata. Vet. World 2015, 8, 1078–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, M.J.; Breitschwerdt, E.; Cleaveland, S.; Karkare, U.; Khanna, C.; Kirpensteijn, J.; Kuiken, T.; Lappin, M.R.; McQuiston, J.; Mumford, E.; et al. Surveillance of Zoonotic Infectious Disease Transmitted by Small Companion Animals. Emerg. Infect. Dis. 2012, 18, e1. [Google Scholar] [CrossRef]
- Moretti, A.; Agnetti, F.; Mancianti, F.; Nardoni, S.; Righi, C.; Moretta, I.; Morganti, G.; Papini, M. Dermatophytosis in Animals: Epidemiological, Clinical and Zoonotic Aspects. G. Ital. Dermatol. Venereol. 2013, 148, 563–572. [Google Scholar] [PubMed]
- Martinez-Rossi, N.M.; Peres, N.T.A.; Rossi, A. Antifungal Resistance Mechanisms in Dermatophytes. Mycopathologia 2008, 166, 369–383. [Google Scholar] [CrossRef]
- Falahati, M.; Akhlaghi, L.; Lari, A.R.; Alaghehbandan, R. Epidemiology of Dermatophytoses in an Area South of Tehran, Iran. Mycopathologia 2003, 156, 279–287. [Google Scholar] [CrossRef]
- Rajagopalan, M.; Inamadar, A.; Mittal, A.; Miskeen, A.K.; Srinivas, C.R.; Sardana, K.; Godse, K.; Patel, K.; Rengasamy, M.; Rudramurthy, S.; et al. Expert Consensus on The Management of Dermatophytosis in India (ECTODERM India). BMC Dermatol. 2018, 18, 6. [Google Scholar] [CrossRef]
- Dworecka-Kaszak, B.; Dąbrowska, I. Dermatophytes: New Taxonomy and Differentiation Methods. Review of Current State of Knowledge about Mechanisms of Pathogenesis and Pathogen-Host Interaction. Med. Weter. 2017, 73, 613–617. [Google Scholar] [CrossRef] [Green Version]
- Baumbach, C.M.; Müller, S.; Reuschel, M.; Uhrlaß, S.; Nenoff, P.; Baums, C.G.; Schrödl, W. Identification of Zoophilic Dermatophytes Using MALDI-TOF Mass Spectrometry. Front. Cell. Infect. Microbiol. 2021, 11, 365. [Google Scholar] [CrossRef]
- Petrucelli, M.F.; de Abreu, M.H.; Cantelli, B.A.M.; Segura, G.G.; Nishimura, F.G.; Bitencourt, T.A.; Marins, M.; Fachin, A.L. Epidemiology and Diagnostic Perspectives of Dermatophytoses. J. Fungi 2020, 6, 310. [Google Scholar] [CrossRef]
- Gallo, M.G.; Lanfranchi, P.; Poglayen, G.; Calderola, S.; Menzano, A.; Ferroglio, E.; Peano, A.; Lanfranchi, P.; Poglayen, G.; Calderola, S.; et al. Seasonal 4-Year Investigation into the Role of the Alpine Marmot (Marmota marmota) as a Carrier of Zoophilic Dermatophytes. Med. Mycol. 2005, 43, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Tizzani, P.; Gallo, M.G.; Peano, A.; Molinar Min, A.; Martínez-Carrasco Pleite, C.; Meneguz, P.G. Dermatophytosis Caused by Microsporum canis in Eastern Cottontail (Sylvilagus floridanus). Eur. J. Wildl. Res. 2007, 53, 238–240. [Google Scholar] [CrossRef]
- Gnat, S.; Łagowski, D.; Nowakiewicz, A.; Dyląg, M. Unusual Dermatomycoses Caused by Nannizzia nana: The Geophilic Origin of Human Infections. Infection 2020, 48, 429–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhrlaß, S.; Schroedl, W.; Mehlhorn, C.; Krüger, C.; Hubka, V.; Maier, T.; Gräser, Y.; Paasch, U.; Nenoff, P. Molecular Epidemiology of Trichophyton quinckeanum—A Zoophilic Dermatophyte on the Rise. JDDG—J. Ger. Soc. Dermatol. 2018, 16, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sdnchez, M.S.; Pereiro, M.; Pereiro, M.M.; Toribio, J. Favus Due to Trichophyton mentagrophytes var. quinckeanum. Dermatology 1997, 194, 177–179. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a Novel Multilocus Phylogenetic Taxonomy for the Dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef] [Green Version]
- Czaika, V.A.; Lam, P.A. Trichophyton mentagrophytes Cause Underestimated Contagious Zoophilic Fungal Infection. Mycoses 2013, 56, 33–37. [Google Scholar] [CrossRef]
- Szathmary, S. Extensive human mouse favus endemic in the Hungarian plateau. Mykosen 1966, 4, 50–63. [Google Scholar]
- García-Romero, M.T.; Arenas, R. New Insights into Genes, Immunity, and the Occurrence of Dermatophytosis. J. Investig. Dermatol. 2015, 135, 655–657. [Google Scholar] [CrossRef] [Green Version]
- Lysková, P.; Dobiáš, R.; Čmoková, A.; Kolařík, M.; Hamal, P.; Šmatláková, K.; Hušek, J.; Mencl, K.; Mallátová, N.; Poláčková, Z.; et al. An Outbreak of Trichophyton quinckeanum Zoonotic Infections in the Czech Republic Transmitted from Cats and Dogs. J. Fungi 2021, 7, 684. [Google Scholar] [CrossRef]
- Beguin, H.; Pyck, N.; Hendrickx, M.; Planard, C.; Stubbe, D.; Detandt, M. The Taxonomic Status of Trichophyton quinckeanum and T. interdigitale Revisited: A Multigene Phylogenetic Approach. Med. Mycol. 2012, 87, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Makimura, K.; Tamura, Y.; Mochizuki, T.; Hasegawa, A.; Tajiri, Y.; Hanazawa, R.; Uchida, K.; Saito, H.; Yamaguchi, H. Phylogenetic Classification and Species Identification of Dermatophyte Strains Based on DNA Sequences of Nuclear Ribosomal Internal Transcribed Spacer 1 Regions. J. Clin. Microbiol. 1999, 37, 920–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, B.; Mirhendi, H.; Shidfar, M.R.; Nouripour-Sisakht, S.; Jalalizand, N.; Geramishoar, M.; Shokoohi, G.R. A Comparative Study on Morphological versus Molecular Identification of Dermatophyte Isolates. J. Mycol. Med. 2015, 25, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Hoog, G.; de Guarro, J.; Gené, J.; Figueras, M. Atlas of Clinical Fungi, 2nd ed.; Centraalbureau voor Schimmelcultures (CBS): Utrecht, The Netherlands, 2000. [Google Scholar]
- Packeu, A.; Hendrickx, M.; Beguin, H.; Martiny, D.; Vandenberg, O.; Detandt, M. Identification of the Trichophyton mentagrophytes Complex Species Using MALDI-TOF Mass Spectrometry. Med. Mycol. 2013, 51, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J.; Otčenášek, M. Zoophilic Dermatophytes Commonly Attacking Man. In Mycological Diagnosis of Animal Dermatophytoses; Dvořák, J., Otčenášek, M., Eds.; Springer: Dordrecht, The Netherlands, 1969; pp. 69–84. ISBN 978-94-010-3426-5. [Google Scholar]
- La Touche, C.J. Mouse Favus Due to Trichophyton quinckeanum (Zopf) MacLeod & Muende: A Reappraisal in the Light of Recent 447 Investigations. Mycopatol. Mycol. Appl. 1959, 11, 257–276. [Google Scholar]
- Frymus, T.; Gruffydd-Jones, T.; Pennisi, M.G.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Hartmann, K.; Hosie, M.J.; Lloret, A.; et al. Dermatophytosis in Cats. J. Feline Med. Surg. 2013, 15, 598–604. [Google Scholar] [CrossRef]
- Moriello, K.A.; Coyner, K.; Paterson, S.; Mignon, B. Diagnosis and Treatment of Dermatophytosis in Dogs and Cats. Vet. Dermatol. 2017, 28, 266-e68. [Google Scholar] [CrossRef]
- Łagowski, D.; Gnat, S.; Nowakiewicz, A.; Osińska, M.; Trościańczyk, A.; Zięba, P. In Search of the Source of Dermatophytosis: Epidemiological Analysis of Trichophyton verrucosum Infection in Llamas and the Breeder (Case Report). Zoonoses Public Health 2019, 66, 982–989. [Google Scholar] [CrossRef]
- Tekin, H.G.; Sigsgaard, V.; Zachariae, C.; Hare, R.K.; Arendrup, M.C.; Saunte, D.M.L. Would You like to Purchase a Rodent with Dermatophytes? Mycoses 2019, 62, 584–587. [Google Scholar] [CrossRef]
- Śpiewak, R.; Szostak, W. Zoophilic and Geophilic Dermatophytoses among Farmers and Non-Farmers in Eastern Poland. Ann. Agric. Environ. Med. 2000, 7, 125–129. [Google Scholar]
- Bilek, J.; Baranova, Z.; Kozak, M.; Fialkovicova, M.; Weissova, T.; Sesztakova, E. Trichophyton mentagrophytes var. quinckeanum as a Cause of Zoophilic Dermatomycosis in a Human Family. Bratisl. Lek. Listy 2005, 106, 383–385. [Google Scholar]
- Besbes, M.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Bouassida, S.; Ayadi, A. Favus Due to Trichophyton mentagrophytes var. quinckeanum. Mycoses 2003, 46, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Skořepová, M.; Štork, J.; Hrabáková, J. Case Reports. Tinea Gladiatorum Due to Trichophyton mentagrophytes. Mycoses 2002, 45, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Niewerth, M.; Splanemann, V.; Korting, H.C.; Ring, J.; Abeck, D. Antimicrobial Susceptibility Testing of Dermatophytes—Comparison of the Agar Macrodilution and Broth Microdilution Tests. Chemotherapy 1998, 44, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woznicki, P.; Wilczynska, J. Koty i Psy w Polsce Oraz Charakterystyka Właścicieli i Ich Postawa Względem Zwierząt. Weter. Prakt. 2017, 14, 10–12. [Google Scholar]
- Gnat, S.; Nowakiewicz, A.; Ziółkowska, G.; Trościańczyk, A.; Majer-Dziedzic, B.; Zięba, P. Evaluation of Growth Conditions and DNA Extraction Techniques Used in the Molecular Analysis of Dermatophytes. J. Appl. Microbiol. 2017, 122, 1368–1379. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 0-12-372180-6. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; ISBN 1562388304. [Google Scholar]
- Maurya, V.K.; Kachhwaha, D.; Bora, A.; Khatri, P.K.; Rathore, L. Determination of antifungal minimum inhibitory concentration and its clinical correlation among treatment failure cases of dermatophytosis. J. Fam. Med. Prim. Care 2019, 8, 2577–2581. [Google Scholar] [CrossRef]
- Shaw, D.; Singh, S.; Dogra, S.; Jayaraman, J.; Bhat, R.; Panda, S.; Chakrabarti, A.; Anjum, N.; Chowdappa, A.; Nagamoti, M.; et al. MIC and Upper Limit of Wild-Type Distribution for 13 Antifungal Agents against a Trichophyton mentagrophytes-Trichophyton interdigitale Complex of Indian Origin. Antimicrob. Agents Chemother. 2020, 64, e01964-19. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| No. of Patients | Sex | Age | Type of Infection | Month of Isolation | Contact with Cat | Race | Leaving Home | Confirmed Infection | Treatment | |
|---|---|---|---|---|---|---|---|---|---|---|
| Humans | Animals | |||||||||
| 1 | F | 23 | tinea capitis | April | + | European Shorthair Cat | freely | asymptomatic | Terbinafine | Miconazole, chlorhexidine |
| 2 | M | 62 | tinea corporis | March | + | Dachshund | freely | asymptomatic | Ciclopirox, terbinafine | Enilconazole |
| 3 | F | 32 | tinea capitis | April | + | Siberian Cat | freely | asymptomatic | Terbinafine | Terbinafine, chlorexidine |
| 4 | F | 80 | tinea corporis | May | + | Ragdoll | with the owner | asymptomatic | Clotrimazole, terbinafine | Terbinafine |
| 5 | M | 68 | tinea capitis | October | + | European Cat | freely | asymptomatic | Fusidic acid, ciclopirox, terbinafine | Miconazole, chlorhexidine |
| 6 | F | 79 | tinea corporis | May | + | European Cat | freely | asymptomatic | Terbinafine, ciclopirox | Miconazole, chlorhexidine |
| 7 | F | 45 | tinea corporis | July | + | Dachshund | freely | asymptomatic | Fusidic acid, ciclopirox, terbinafine | Sulphur lyme |
| 8 | M | 69 | tinea capitis | April | + | Scottish Fold | with the owner | none | Terbinafine | NA |
| 9 | F | 31 | tinea corporis | June | + | Persian | with the owner | none | Clotrimazole, terbinafine | NA |
| 10 | F | 70 | tinea corporis | May | + | British shorthair | with the owner | asymptomatic | Terbinafine, ciclopirox, clotrimazole | Enilconazole |
| 11 | F | 81 | tinea capitis | March | + | European Cat | freely | asymptomatic | Fusidic acid, ciclopirox, terbinafine | Miconazole, chlorhexidine |
| 12 | F | 52 | tinea capitis | July | + | Archangel Blue | with the owner | asymptomatic | Clotrimazole, terbinafine | Enilconazole |
| 13 | F | 74 | tinea corporis | June | + | Persian | no possibility | none | Clotrimazole, ciclopirox | NA |
| 14 | F | 69 | tinea corporis | May | + | European Cat | freely | asymptomatic | Clotrimazole, ciclopirox | Miconazole, chlorhexidine |
| 15 | M | 75 | tinea faciei | March | + | Siamese | no possibility | none | Ciclopirox, terbinafine | NA |
| 16 | M | 28 | tinea corporis | April | + | Dachshund | freely | asymptomatic | Terbinafine, ciclopirox, clotrimazole | Miconazole, chlorhexidine |
| 17 | F | 76 | tinea corporis | July | + | Sphynx | no possibility | none | Clotrimazole, terbinafine | NA |
| 18 | M | 60 | tinea capitis | November | + | Bengal cat | no possibility | none | Clotrimazole, terbinafine | NA |
| 19 | F | 32 | tinea corporis | April | + | British shorthair | with the owner | asymptomatic | Terbinafine, ciclopirox | Sulfur lyme |
| 20 | M | 40 | tinea corporis | July | + | Abyssinian cat | with the owner | asymptomatic | Clotrimazole, terbinafine | Terbinafine, chlorohexidine |
| 21 | M | 37 | tinea capitis | May | + | British Longhair | with the owner | asymptomatic | Terbinafine | Enilconazole |
| Antifungal Agents | Host | MIC (µg/mL) | MIC Range | MIC50 | MIC90 | MICGM | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.004 | 0.008 | 0.016 | 0.03 | 0.06 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | |||||||
| Allylamine | NFT | humans | 3 | 6 | 12 | 0.004–0.016 | 0.016 | 0.016 | 0.012 | |||||||||||
| animals | 13 | 2 | 0.008–0.016 | 0.008 | 0.008 | 0.009 | ||||||||||||||
| TRB | humans | 4 | 10 | 7 | 0.004–0.016 | 0.008 | 0.016 | 0.01 | ||||||||||||
| animals | 11 | 4 | 0.008–0.016 | 0.008 | 0.016 | 0.01 | ||||||||||||||
| Imidazoles | KTC | humans | 4 | 5 | 6 | 5 | 0.125–1 | 0.5 | 1 | 0.46 | ||||||||||
| animals | 5 | 7 | 1 | 2 | 0.125–1 | 0.25 | 1 | 0.33 | ||||||||||||
| MCZ | humans | 1 | 5 | 10 | 5 | 0.008–0.06 | 0.03 | 0.06 | 0.03 | |||||||||||
| animals | 4 | 11 | 0.016–0.03 | 0.03 | 0.03 | 0.03 | ||||||||||||||
| ENC | humans | 6 | 12 | 2 | 1 | 0.03–0.25 | 0.06 | 0.125 | 0.07 | |||||||||||
| animals | 2 | 8 | 5 | 0.03–0.125 | 0.06 | 0.125 | 0.08 | |||||||||||||
| CLT | humans | 3 | 6 | 8 | 4 | 0.06–0.5 | 0.25 | 0.5 | 0.19 | |||||||||||
| animals | 1 | 14 | 0.125–0.25 | 0.25 | 0.25 | 0.24 | ||||||||||||||
| Triazoles | ITC | humans | 7 | 9 | 4 | 1 | 0.03–0.25 | 0.06 | 0.125 | 0.07 | ||||||||||
| animals | 14 | 1 | 0.06–0.125 | 0.06 | 0.125 | 0.06 | ||||||||||||||
| FLC | humans | 2 | 6 | 7 | 3 | 3 | 2–32 | 8 | 32 | 10.85 | ||||||||||
| animals | 9 | 4 | 2 | 8–32 | 8 | 32 | 13.33 | |||||||||||||
| VRC | humans | 3 | 11 | 5 | 2 | 0.03–0.25 | 0.06 | 0.25 | 0.09 | |||||||||||
| animals | 1 | 10 | 4 | 0.03–0.125 | 0.06 | 0.125 | 0.08 | |||||||||||||
| Pyridinone derivatives | CPO | humans | 2 | 3 | 8 | 8 | 0.06–0.5 | 0.25 | 0.5 | 0.31 | ||||||||||
| animals | 4 | 9 | 2 | 0.125–0.5 | 0.25 | 0.5 | 0.25 | |||||||||||||
| Phenyl morpholine derivatives | AMR | humans | 6 | 7 | 7 | 1 | 0.008–0.06 | 0.016 | 0.03 | 0.02 | ||||||||||
| animals | 1 | 9 | 5 | 0.008–0.03 | 0.016 | 0.03 | 0.02 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łagowski, D.; Gnat, S.; Dyląg, M.; Nowakiewicz, A. Laboratory Diagnosis and In Vitro Antifungal Susceptibility of Trichophyton quinckeanum from Human Zoonoses and Cats. Antibiotics 2022, 11, 739. https://doi.org/10.3390/antibiotics11060739
Łagowski D, Gnat S, Dyląg M, Nowakiewicz A. Laboratory Diagnosis and In Vitro Antifungal Susceptibility of Trichophyton quinckeanum from Human Zoonoses and Cats. Antibiotics. 2022; 11(6):739. https://doi.org/10.3390/antibiotics11060739
Chicago/Turabian StyleŁagowski, Dominik, Sebastian Gnat, Mariusz Dyląg, and Aneta Nowakiewicz. 2022. "Laboratory Diagnosis and In Vitro Antifungal Susceptibility of Trichophyton quinckeanum from Human Zoonoses and Cats" Antibiotics 11, no. 6: 739. https://doi.org/10.3390/antibiotics11060739
APA StyleŁagowski, D., Gnat, S., Dyląg, M., & Nowakiewicz, A. (2022). Laboratory Diagnosis and In Vitro Antifungal Susceptibility of Trichophyton quinckeanum from Human Zoonoses and Cats. Antibiotics, 11(6), 739. https://doi.org/10.3390/antibiotics11060739