
The Bacterial Urban Resistome: Recent Advances


Abstract
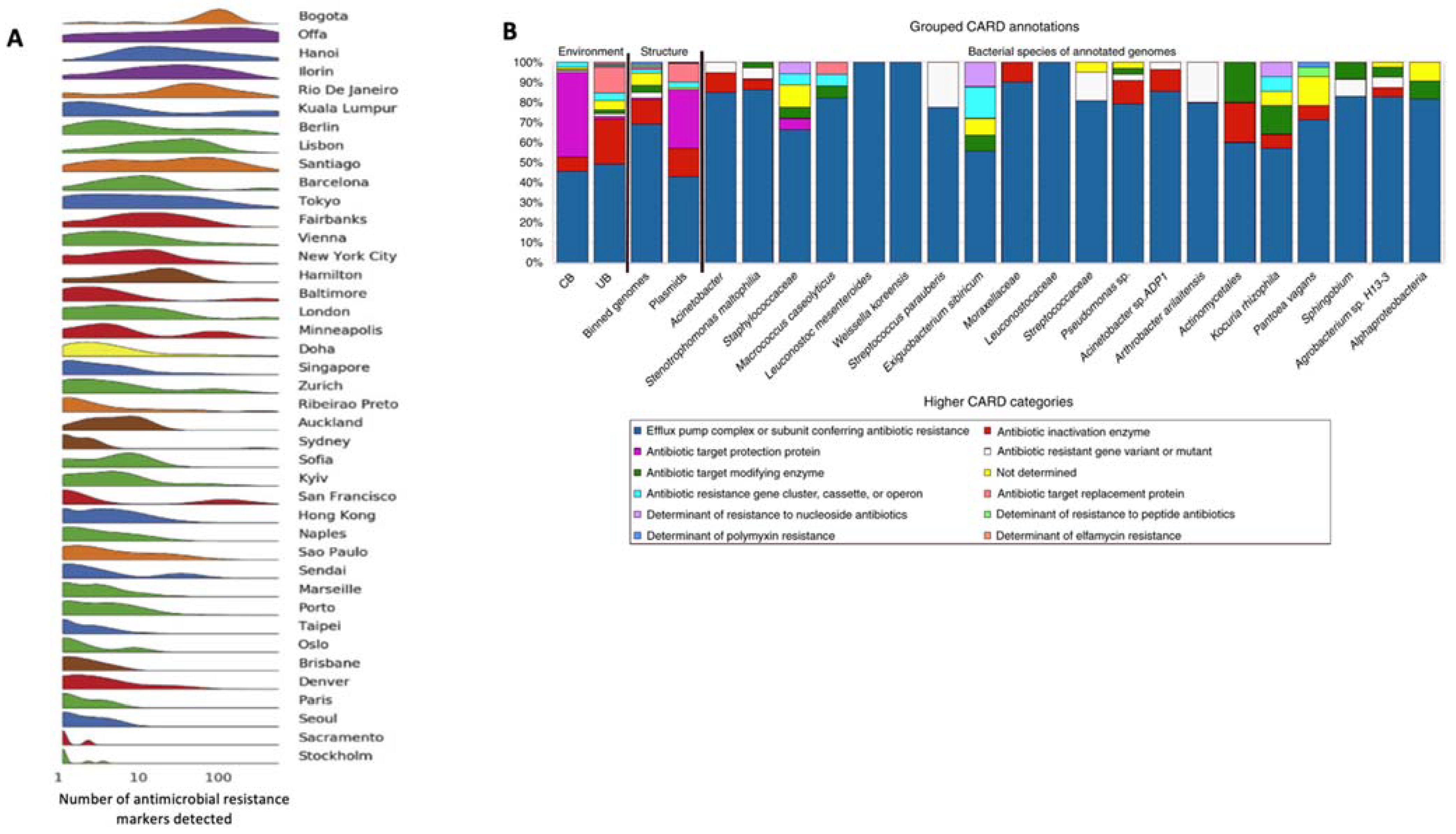
1. Introduction
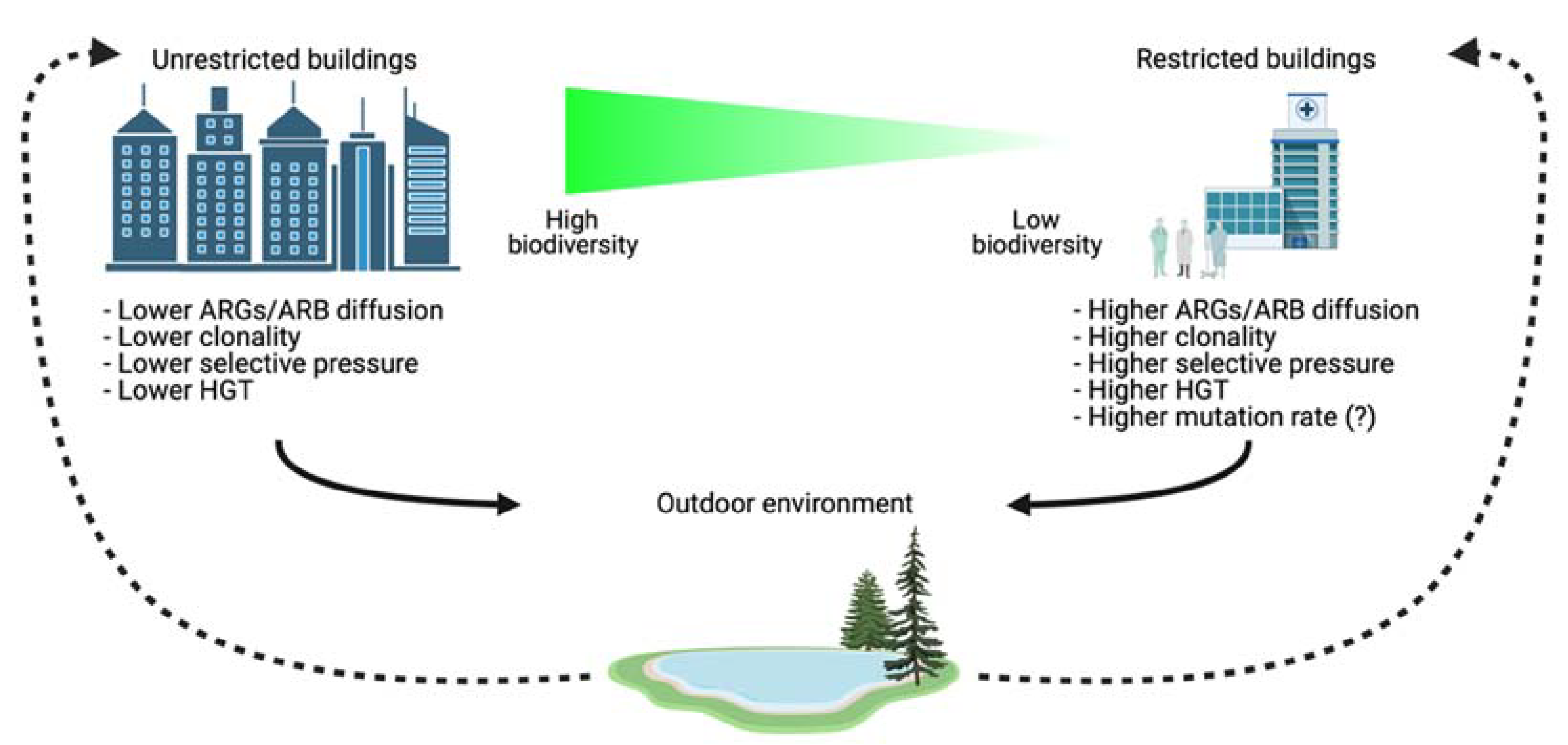
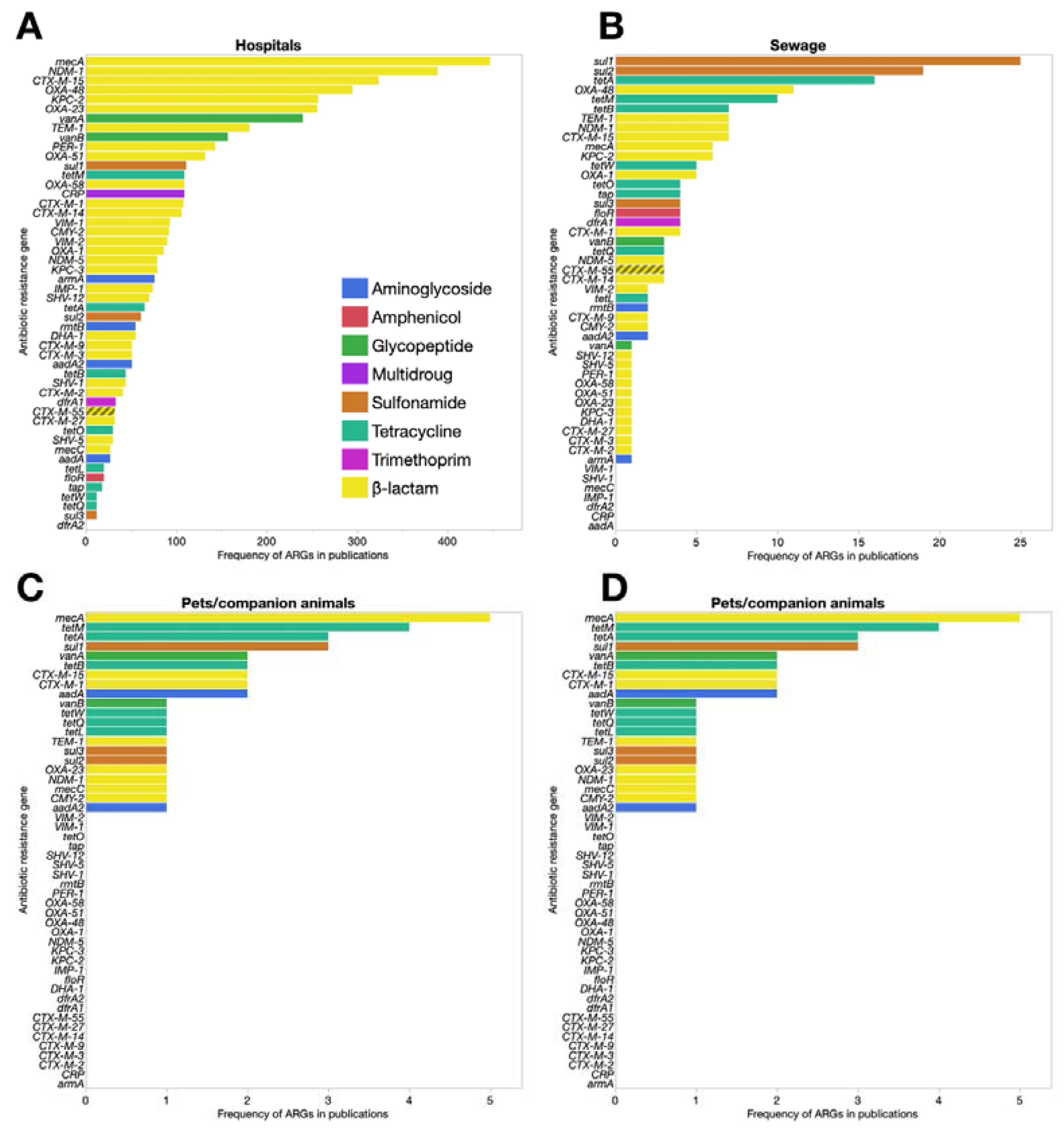
2. Built Environments
3. Transportation
3.1. Air Transport
3.2. Resistomes in Trains and Metro
3.3. Shared Transportation
4. Urban Green Spaces and Parks
5. Companion Animals
6. Urban Wastewater
7. The Urban Network of ARB/ARGs
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pouyat, R.V.; Szlavecz, K.; Yesilonis, I.D.; Groffman, P.M.; Schwarz, K. Characteristics of Urban Soils. Chem. Phys. Biol. 2010, 55, 119–152. [Google Scholar] [CrossRef]
- Ritchie, H. Urbanization; Our World Data; 2018. Available online: https://ourworldindata.org/urbanization (accessed on 13 October 2021).
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Surette, M.D.; Wright, G.D. Lessons from the Environmental Antibiotic Resistome. Annu. Rev. Microbiol. 2017, 71, 309–329. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Westman, E.L.; Wright, G.D. The antibiotic resistome: What’s new? Curr. Opin. Microbiol. 2014, 21, 45–50. [Google Scholar] [CrossRef]
- Brodie, E.L.; DeSantis, T.Z.; Moberg Parker, J.P.; Zubietta, I.X.; Piceno, Y.M.; Andersen, G.L. Urban aerosols harbor diverse and dynamic bacterial populations. Proc. Natl. Acad. Sci. USA 2007, 104, 299–304. [Google Scholar] [CrossRef]
- Lupo, A.; Coyne, S.; Berendonk, T.U. Origin and evolution of antibiotic resistance: The common mechanisms of emergence and spread in water bodies. Front. Microbiol. 2012, 3, 18. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, R.; Chen, B.; Zhang, T.; Hu, L.; Zou, S. Characterization of airborne antibiotic resistance genes from typical bioaerosol emission sources in the urban environment using metagenomic approach. Chemosphere 2018, 213, 463–471. [Google Scholar] [CrossRef]
- Chen, Y.M.; Holmes, E.C.; Chen, X.; Tian, J.H.; Lin, X.D.; Qin, X.C.; Gao, W.H.; Liu, J.; Wu, Z.D.; Zhang, Y.Z. Diverse and abundant resistome in terrestrial and aquatic vertebrates revealed by transcriptional analysis. Sci. Rep. 2020, 10, 18870. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, L.; Li, F.; Huang, S.; Yang, J.; Ma, C.; Zhang, D.; Yu, C.P.; Hu, A. Urban ponds as hotspots of antibiotic resistome in the urban environment. J. Hazard. Mater. 2021, 403, 124008. [Google Scholar] [CrossRef]
- Danko, D.; Bezdan, D.; Afshin, E.E.; Ahsanuddin, S.; Bhattacharya, C.; Butler, D.J.; Chng, K.R.; Donnellan, D.; Hecht, J.; Jackson, K.; et al. A global metagenomic map of urban microbiomes and antimicrobial resistance. Cell 2021, 184, 3376–3393.e17. [Google Scholar] [CrossRef]
- Mahnert, A.; Moissl-Eichinger, C.; Zojer, M.; Bogumil, D.; Mizrahi, I.; Rattei, T.; Martinez, J.L.; Berg, G. Man-made microbial resistances in built environments. Nat. Commun. 2019, 10, 968. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, A.; Kett, S.; Purchase, D.; Marvasi, M. Antibiotic-Resistant Genes and Bacteria as Evolving Contaminants of Emerging Concerns (e-CEC): Is It Time to Include Evolution in Risk Assessment? Antibiotics 2021, 10, 1066. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex-32018D0840-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/en/TXT/?uri=CELEX%3A32018D0840 (accessed on 13 October 2021).
- Lanza, V.F.; Baquero, F.; Martínez, J.L.; Ramos-Ruíz, R.; González-Zorn, B.; Andremont, A.; Sánchez-Valenzuela, A.; Ehrlich, S.D.; Kennedy, S.; Ruppé, E.; et al. In-depth resistome analysis by targeted metagenomics. Microbiome 2018, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Leff, J.W.; Barberán, A.; Bates, S.T.; Betley, J.; Crowther, T.W.; Kelly, E.F.; Oldfield, E.E.; Ashley Shaw, E.; Steenbock, C.; et al. Biogeographic patterns in below-ground diversity in New York City’s Central Park are similar to those observed globally. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141988. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.; Achmon, Y.; Cao, Y.; Liang, X.; Chen, L.; Wang, H.; Siame, B.A.; Leung, K.Y. Distribution of antibiotic resistance genes in the environment. Environ. Pollut. 2021, 285, 117402. [Google Scholar] [CrossRef] [PubMed]
- Custovic, A.; Taggart, S.C.O.; Woodcock, A. House dust mite and cat allergen in different indoor environments. Clin. Exp. Allergy 1994, 24, 1164–1168. [Google Scholar] [CrossRef]
- Höppe, P.; Martinac, I. Indoor climate and air quality. Int. J. Biometeorol. 1998, 42, 1–7. [Google Scholar] [CrossRef]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S.; et al. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef]
- Ben Maamar, S.; Hu, J.; Hartmann, E.M. Implications of indoor microbial ecology and evolution on antibiotic resistance. J. Expo. Sci. Environ. Epidemiol. 2020, 30, 1–15. [Google Scholar] [CrossRef]
- Ben Maamar, S.; Glawe, A.J.; Brown, T.K.; Hellgeth, N.; Hu, J.; Wang, J.P.; Huttenhower, C.; Hartmann, E.M. Mobilizable antibiotic resistance genes are present in dust microbial communities. PLoS Pathog. 2020, 16, e1008211. [Google Scholar] [CrossRef]
- Ho, J.; Yeoh, Y.K.; Barua, N.; Chen, Z.; Lui, G.; Wong, S.H.; Yang, X.; Chan, M.C.; Chan, P.K.S.; Hawkey, P.M.; et al. Systematic review of human gut resistome studies revealed variable definitions and approaches. Gut Microbes 2020, 12, 1700755. [Google Scholar] [CrossRef] [PubMed]
- Peacock, S.; Paterson, G. Mechanisms of Methicillin Resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef] [PubMed]
- Nain, Z.; Karim, M.M. Whole-genome sequence, functional annotation, and comparative genomics of the high biofilm-producing multidrug-resistant Pseudomonas aeruginosa MZ4A isolated from clinical waste. Gene Rep. 2021, 22, 100999. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; Dang, C.; Zhang, M.; Zhao, B.; Cheng, Z.; Chen, L.; Zhong, Z.; Ye, Y.; Xia, Y. A metagenomic-based method to study hospital air dust resistome. Chem. Eng. J. 2021, 406, 126854. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Hartmann, E.M. Anthropogenic chemicals and their impacts on microbes living in buildings. Microb. Biotechnol. 2021, 14, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Lee, S.; Bisesi, M.; Lee, J. Indoor microbiome and antibiotic resistance on floor surfaces: An exploratory study in three different building types. Int. J. Environ. Res. Public Health 2019, 16, 4160. [Google Scholar] [CrossRef]
- Gwenzi, W. The ‘thanato-resistome’—The funeral industry as a potential reservoir of antibiotic resistance: Early insights and perspectives. Sci. Total Environ. 2020, 749, 141120. [Google Scholar] [CrossRef]
- Heß, S.; Kneis, D.; Österlund, T.; Li, B.; Kristiansson, E.; Berendonk, T.U. Sewage from Airplanes Exhibits High Abundance and Diversity of Antibiotic Resistance Genes. Environ. Sci. Technol. 2019, 53, 13898–13905. [Google Scholar] [CrossRef]
- Petersen, T.N.; Rasmussen, S.; Hasman, H.; Carøe, C.; Bælum, J.; Charlotte Schultz, A.; Bergmark, L.; Svendsen, C.A.; Lund, O.; Sicheritz-Pontén, T.; et al. Meta-genomic analysis of toilet waste from long distance flights; A step towards global surveillance of infectious diseases and antimicrobial resistance. Sci. Rep. 2015, 5, 11444. [Google Scholar] [CrossRef]
- Cloran, F.J. Cutaneous infections with community-acquired MRSA in aviators. Aviat. Sp. Environ. Med. 2006, 77, 1271–1274. [Google Scholar]
- Grover, S. Skin in aviation and space environment. Indian J. Dermatol. Venereol. Leprol. 2011, 77, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Ni, Y.; Li, J.; Imamovic, L.; Sarkar, C.; Kobler, M.D.; Heshiki, Y.; Zheng, T.; Kumari, S.; Wong, J.C.Y.; et al. The Environmental Exposures and Inner- and Intercity Traffic Flows of the Metro System May Contribute to the Skin Microbiome and Resistome. Cell Rep. 2018, 24, 1190–1202.e5. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Feng, S.; Chen, H.; Dai, M.; Paterson, D.L.; Zheng, X.; Wu, X.; Zhong, L.L.; Liu, Y.; Xia, Y.; et al. Transmission of mcr-1-Producing Multidrug-resistant Enterobacteriaceae in Public Transportation in Guangzhou, China. Clin. Infect. Dis. 2018, 67, S217–S224. [Google Scholar] [CrossRef]
- Iwao, Y.; Yabe, S.; Takano, T.; Higuchi, W.; Nishiyama, A.; Yamamoto, T. Isolation and molecular characterization of methicillin-resistant Staphylococcus aureus from public transport. Microbiol. Immunol. 2012, 56, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Wang, Y. Characteristics of antibiotic resistance of airborne Staphylococcus isolated from metro stations. Int. J. Environ. Res. Public Health 2013, 10, 2412–2426. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Ou, Q.; Lin, D.; Xu, P.; Li, Y.; Ye, X.; Zhou, J.; Yao, Z. Metro system in Guangzhou as a hazardous reservoir of methicillin-resistant Staphylococci: Findings from a point-prevalence molecular epidemiologic study. Sci. Rep. 2015, 5, 16087. [Google Scholar] [CrossRef][Green Version]
- Lin, J.L.; Peng, Y.; Ou, Q.T.; Lin, D.X.; Li, Y.; Ye, X.H.; Zhou, J.L.; Yao, Z.J. A molecular epidemiological study of methicillin-resistant Staphylococci environmental contamination in railway stations and coach stations in Guangzhou of China. Lett. Appl. Microbiol. 2017, 64, 131–137. [Google Scholar] [CrossRef]
- Wei, T.; Yao, H.; Sun, P.; Cai, W.; Li, X.; Fan, L.; Wei, Q.; Lai, C.; Guo, J. Mitigation of antibiotic resistance in a pilot-scale system treating wastewater from high-speed railway trains. Chemosphere 2020, 245, 125484. [Google Scholar] [CrossRef]
- Gu, J.; Xie, X.-J.; Liu, J.-X.; Shui, J.-R.; Zhang, H.-Y.; Feng, G.-Y.; Liu, X.-Y.; Li, L.-C.; Lan, Q.-W.; Jin, Q.-H.; et al. Prevalence and transmission of antimicrobial-resistant Staphylococci and Enterococci from shared bicycles in Chengdu, China. Sci. Total Environ. 2020, 738, 139735. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, S.; Chen, L.; Liu, Y.; Tan, L.; Shen, J.; Zhang, W. Antimicrobial resistance and molecular characterization of methicillin-resistant coagulase-negative staphylococci from public shared bicycles in Tianjin, China. J. Glob. Antimicrob. Resist. 2019, 19, 231–235. [Google Scholar] [CrossRef]
- Zou, Z.-Y.; Lei, L.; Chen, Q.-Y.; Wang, Y.-Q.; Cai, C.; Li, W.-Q.; Zhang, Z.; Shao, B.; Wang, Y. Prevalence and dissemination risk of antimicrobial-resistant Enterobacteriaceae from shared bikes in Beijing, China. Environ. Int. 2019, 132, 105119. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xie, J.; Li, J.; Zhao, J.; Qiao, S.; Li, Y.; Zeng, J. Shared bicycle microbial community: A potential antibiotic-resistant bacteria warehouse. Folia Microbiol. 2020, 66, 49–58. [Google Scholar] [CrossRef] [PubMed]
- de Keijzer, C.; Gascon, M.; Nieuwenhuijsen, M.J.; Dadvand, P. Long-Term Green Space Exposure and Cognition Across the Life Course: A Systematic Review. Curr. Environ. Health Rep. 2016, 3, 468–477. [Google Scholar] [CrossRef] [PubMed]
- United Nations. World Urbanization Prospects: The 2018 Revision. Available online: https://population.un.org/wup/Publications/Files/WUP2018-Report.pdf (accessed on 13 October 2021).
- Sandifer, P.A.; Sutton-Grier, A.E.; Ward, B.P. Exploring connections among nature, biodiversity, ecosystem services, and human health and well-being: Opportunities to enhance health and biodiversity conservation. Ecosyst. Serv. 2015, 12, 1–15. [Google Scholar] [CrossRef]
- Wood, E.; Harsant, A.; Dallimer, M.; de Chavez, A.C.; McEachan, R.R.C.; Hassall, C. Not all green space is created equal: Biodiversity predicts psychological restorative benefits from urban green space. Front. Psychol. 2018, 9, 2320. [Google Scholar] [CrossRef]
- Knobel, P.; Dadvand, P.; Maneja-Zaragoza, R. A systematic review of multi-dimensional quality assessment tools for urban green spaces. Heal. Place 2019, 59, 102198. [Google Scholar] [CrossRef]
- Charlop-Powers, Z.; Pregitzer, C.C.; Lemetre, C.; Ternei, M.A.; Maniko, J.; Hover, B.M.; Calle, P.Y.; McGuire, K.L.; Garbarino, J.; Forgione, H.M.; et al. Urban park soil microbiomes are a rich reservoir of natural product biosynthetic diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 14811–14816. [Google Scholar] [CrossRef]
- Worsley-Tonks, K.E.L.; Miller, E.A.; Gehrt, S.D.; McKenzie, S.C.; Travis, D.A.; Johnson, T.J.; Craft, M.E. Characterization of antimicrobial resistance genes in Enterobacteriaceae carried by suburban mesocarnivores and locally owned and stray dogs. Zoonoses Public Health 2020, 67, 460–466. [Google Scholar] [CrossRef]
- Mo, S.S.; Urdahl, A.M.; Madslien, K.; Sunde, M.; Nesse, L.L.; Slettemeås, J.S.; Norström, M. What does the fox say? Monitoring antimicrobial resistance in the environment using wild red foxes as an indicator. PLoS ONE 2018, 13, e0198019. [Google Scholar] [CrossRef]
- Reynolds, L.J.; Sala-Comorera, L.; Martin, N.A.; Nolan, T.M.; Stephens, J.H.; Gitto, A.; O’Hare, G.M.P.; O’Sullivan, J.J.; Meijer, W.G. Correlation between antimicrobial resistance and faecal contamination in small urban streams and bathing waters. Sci. Total Environ. 2020, 739, 140242. [Google Scholar] [CrossRef]
- Zhao, H.; Sun, R.; Yu, P.; Alvarez, P.J.J. High levels of antibiotic resistance genes and opportunistic pathogenic bacteria indicators in urban wild bird feces. Environ. Pollut. 2020, 266, 115200. [Google Scholar] [CrossRef] [PubMed]
- Elmberg, J.; Berg, C.; Lerner, H.; Waldenström, J.; Hessel, R. Potential disease transmission from wild geese and swans to livestock, poultry and humans: A review of the scientific literature from a One Health perspective. Infect. Ecol. Epidemiol. 2017, 7, 1300450. [Google Scholar] [CrossRef] [PubMed]
- Agnew, A.; Wang, J.; Fanning, S.; Bearhop, S.; McMahon, B.J. Insights into antimicrobial resistance among long distance migratory East Canadian High Arctic light-bellied Brent geese (Branta bernicla hrota). Ir. Vet. J. 2016, 69, 13. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Linke, H.J.; Wagner, M. Combining industrial and urban water-reuse concepts for increasing the water resources in water-scarce regions. Water Environ. Res. 2020, 92, 1027–1041. [Google Scholar] [CrossRef]
- Wang, J.; Banzhaf, E. Towards a better understanding of Green Infrastructure: A critical review. Ecol. Indic. 2018, 85, 758–772. [Google Scholar] [CrossRef]
- Zalacáin, D.; Martínez-Pérez, S.; Bienes, R.; García-Díaz, A.; Sastre-Merlín, A. Turfgrass biomass production and nutrient balance of an urban park irrigated with reclaimed water. Chemosphere 2019, 237, 124481. [Google Scholar] [CrossRef]
- Zalacáin, D.; Sastre-Merlín, A.; Martínez-Pérez, S.; Bienes, R.; García-Díaz, A. Assessment on micronutrient concentration after reclaimed water irrigation: A CASE study in green areas of Madrid. Irrig. Drain. 2021, 70, 668–678. [Google Scholar] [CrossRef]
- Limayem, A.; Wasson, S.; Mehta, M.; Pokhrel, A.R.; Patil, S.; Nguyen, M.; Chen, J.; Nayak, B. High-Throughput Detection of Bacterial Community and Its Drug-Resistance Profiling From Local Reclaimed Wastewater Plants. Front. Cell. Infect. Microbiol. 2019, 9, 303. [Google Scholar] [CrossRef]
- Han, X.M.; Hu, H.W.; Shi, X.Z.; Wang, J.T.; Han, L.L.; Chen, D.; He, J.Z. Impacts of reclaimed water irrigation on soil antibiotic resistome in urban parks of Victoria, Australia. Environ. Pollut. 2016, 211, 48–57. [Google Scholar] [CrossRef]
- Yan, Z.Z.; Chen, Q.L.; Zhang, Y.J.; He, J.Z.; Hu, H.W. Antibiotic resistance in urban green spaces mirrors the pattern of industrial distribution. Environ. Int. 2019, 132, 105106. [Google Scholar] [CrossRef]
- Ouyang, W.; Gao, B.; Cheng, H.; Zhang, L.; Wang, Y.; Lin, C.; Chen, J. Airborne bacterial communities and antibiotic resistance gene dynamics in PM2.5 during rainfall. Environ. Int. 2020, 134, 105318. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Shin, S.; Jeong, S.H. Clonal spread of extended-spectrum cephalosporin-resistant enterobacteriaceae between companion animals and humans in South Korea. Front. Microbiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [PubMed]
- Dupouy, V.; Abdelli, M.; Moyano, G.; Arpaillange, N.; Bibbal, D.; Cadiergues, M.C.; Lopez-Pulin, D.; Sayah-Jeanne, S.; De Gunzburg, J.; Saint-Lu, N.; et al. Prevalence of beta-lactam and quinolone/fluoroquinolone resistance in enterobacteriaceae from dogs in France and spain—characterization of ESBL/pAmpC isolates, genes, and conjugative plasmids. Front. Vet. Sci. 2019, 6, 279. [Google Scholar] [CrossRef] [PubMed]
- Baede, V.O.; Wagenaar, J.A.; Broens, E.M.; Duim, B.; Dohmen, W.; Nijsse, R.; Timmerman, A.J.; Hordijk, J. Longitudinal study of extended-spectrum-β-lactamase- and AmpC-Producing Enterobacteriaceae in household dogs. Antimicrob. Agents Chemother. 2015, 59, 3117–3124. [Google Scholar] [CrossRef]
- LaLonde-Paul, D.; Cummings, K.J.; Rodriguez-Rivera, L.D.; Wu, J.; Lawhon, S.D. Ciprofloxacin resistance among Campylobacter jejuni isolates obtained from shelter dogs in Texas. Zoonoses Public Health 2019, 66, 337–342. [Google Scholar] [CrossRef]
- Hartantyo, S.H.P.; Chau, M.L.; Fillon, L.; Ariff, A.Z.B.M.; Kang, J.S.L.; Aung, K.T.; Gutiérrez, R.A. Sick pets as potential reservoirs of antibiotic-resistant bacteria in Singapore. Antimicrob. Resist. Infect. Control 2018, 7, 106. [Google Scholar] [CrossRef]
- Joosten, P.; Ceccarelli, D.; Odent, E.; Sarrazin, S.; Graveland, H.; Van Gompel, L.; Battisti, A.; Caprioli, A.; Franco, A.; Wagenaar, J.A.; et al. Antimicrobial usage and resistance in companion animals: A cross-sectional study in three european countries. Antibiotics 2020, 9, 87. [Google Scholar] [CrossRef]
- Schmidt, V.M.; Pinchbeck, G.; McIntyre, K.M.; Nuttall, T.; McEwan, N.; Dawson, S.; Williams, N.J. Routine antibiotic therapy in dogs increases the detection of antimicrobial-resistant faecal Escherichia coli. J. Antimicrob. Chemother. 2018, 73, 3305–3316. [Google Scholar] [CrossRef]
- Toombs-Ruane, L.J.; Benschop, J.; French, N.P.; Biggs, P.J.; Midwinter, A.C.; Marshall, J.C.; Chan, M.; Drinković, D.; Fayaz, A.; Baker, M.G.; et al. Carriage of Extended-Spectrum-Beta-Lactamase- and AmpC Beta-Lactamase-Producing Escherichia coli Strains from Humans and Pets in the Same Households. Appl. Environ. Microbiol. 2020, 86, e01613-20. [Google Scholar] [CrossRef]
- Carvalho, A.C.; Barbosa, A.V.; Arais, L.R.; Ribeiro, P.F.; Carneiro, V.C.; Cerqueira, A.M.F. Resistance patterns, ESBL genes, and genetic relatedness of Escherichia coli from dogs and owners. Braz. J. Microbiol. 2016, 47, 150–158. [Google Scholar] [CrossRef]
- Hong, J.S.; Song, W.; Park, H.-M.; Oh, J.-Y.; Chae, J.-C.; Jeong, S.; Jeong, S.H. Molecular Characterization of Fecal Extended-Spectrum β-Lactamase- and AmpC β-Lactamase-Producing Escherichia coli From Healthy Companion Animals and Cohabiting Humans in South Korea. Front. Microbiol. 2020, 11, 674. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Okada, E.; Shimizu, T.; Kataoka, Y.; Sawada, T.; Takahashi, T. Antimicrobial resistance, virulence profiles, and phylogenetic groups of fecal Escherichia coli isolates: A comparative analysis between dogs and their owners in Japan. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Chae, J.C.; Han, J.I.; Song, W.K.; Lee, C.M.; Park, H.M. Distribution and epidemiological relatedness of methicillin-resistant Staphylococcus aureus isolated from companion dogs, owners, and environments. J. Vet. Med. Sci. 2020, 82, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, C.E.; Todisco, G.; Montani, A.; Profeta, F.; Di Provvido, A.; Foschi, G.; Persiani, T.; Marsilio, F. Reproductive disorders in domestic canaries (Serinus canarius domesticus): A retrospective study on bacterial isolates and their antimicrobial resistance in Italy from 2009 to 2012. Vet. Ital. 2018, 54, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Diren Sigirci, B.; Celik, B.; Halac, B.; Adiguzel, M.C.; Kekec, I.; Metiner, K.; Ikiz, S.; Bagcigil, A.F.; Ozgur, N.Y.; Ak, S.; et al. Antimicrobial resistance profiles of Escherichia coli isolated from companion birds. J. King Saud Univ. Sci. 2020, 32, 1069–1073. [Google Scholar] [CrossRef]
- Varriale, L.; Dipineto, L.; Russo, T.P.; Borrelli, L.; Romano, V.; D’Orazio, S.; Pace, A.; Menna, L.F.; Fioretti, A.; Santaniello, A. Antimicrobial Resistance of Escherichia coli and Pseudomonas aeruginosa from Companion Birds. Antibiotics 2020, 9, 780. [Google Scholar] [CrossRef]
- Sala, A.; Di Ianni, F.; Pelizzone, I.; Bertocchi, M.; Santospirito, D.; Rogato, F.; Flisi, S.; Spadini, C.; Iemmi, T.; Moggia, E.; et al. The prevalence of Pseudomonas aeruginosa and multidrug resistant Pseudomonas aeruginosa in healthy captive ophidian. PeerJ 2019, 7, e6706. [Google Scholar] [CrossRef]
- Colinon, C.; Jocktane, D.; Brothier, E.; Rossolini, G.M.; Cournoyer, B.; Nazaret, S. Genetic analyses of Pseudomonas aeruginosa isolated from healthy captive snakes: Evidence of high interand intrasite dissemination and occurrence of antibiotic resistance genes. Environ. Microbiol. 2010, 12, 716–729. [Google Scholar] [CrossRef]
- Casey, C.L.; Hernandez, S.M.; Yabsley, M.J.; Smith, K.F.; Sanchez, S. The carriage of antibiotic resistance by enteric bacteria from imported tokay geckos (Gekko gecko) destined for the pet trade. Sci. Total Environ. 2015, 505, 299–305. [Google Scholar] [CrossRef]
- Wendt, M.; Silva, B.C.J.; Heo, G.-J. Virulence Factors and Antimicrobial Resistance of Pseudomonas aeruginosa Isolated from Pet Turtles. Asian J. Anim. Vet. Adv. 2017, 12, 205–211. [Google Scholar] [CrossRef][Green Version]
- Alexander, J.; Hembach, N.; Schwartz, T. Evaluation of antibiotic resistance dissemination by wastewater treatment plant effluents with different catchment areas in Germany. Sci. Rep. 2020, 10, 8952. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Iakovides, I.C.; Manoli, K.; Karaolia, P.; Michael-Kordatou, I.; Manaia, C.M.; Fatta-Kassinos, D. Reduction of antibiotic resistance determinants in urban wastewater by ozone: Emphasis on the impact of wastewater matrix towards the inactivation kinetics, toxicity and bacterial regrowth. J. Hazard. Mater. 2021, 420, 126527. [Google Scholar] [CrossRef] [PubMed]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697, 134023. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Vu, H.P.; Nguyen, L.N.; Wang, Q.; Djordjevic, S.P.; Donner, E.; Yin, H.; Nghiem, L.D. Monitoring antibiotic resistance genes in wastewater treatment: Current strategies and future challenges. Sci. Total Environ. 2021, 783, 146964. [Google Scholar] [CrossRef]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I.; et al. Antibiotic resistance in wastewater treatment plants: Tackling the black box. Environ. Int. 2018, 115, 312–324. [Google Scholar] [CrossRef]
- Uluseker, C.; Kaster, K.M.; Thorsen, K.; Basiry, D.; Shobana, S.; Jain, M.; Kumar, G.; Kommedal, R.; Pala-Ozkok, I. A Review on Occurrence and Spread of Antibiotic Resistance in Wastewaters and in Wastewater Treatment Plants: Mechanisms and Perspectives. Front. Microbiol. 2021, 12, 3003. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L.; Wojnárovits, L.; Takács, E. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: An overview. Sci. Total Environ. 2020, 744, 140997. [Google Scholar] [CrossRef]
- Cao, Y.S.; Tang, J.G.; Henze, M.; Yang, X.P.; Gan, Y.P.; Li, J.; Kroiss, H.; Van Loosdrecht, M.C.M.; Zhang, Y.; Daigger, G.T. The leakage of sewer systems and the impact on the ‘black and odorous water bodies’ and WWTPs in China. Water Sci. Technol. 2019, 79, 334–341. [Google Scholar] [CrossRef]
- Besner, M.C.; Prévost, M.; Regli, S. Assessing the public health risk of microbial intrusion events in distribution systems: Conceptual model, available data, and challenges. Water Res. 2011, 45, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.D.J.; Slocum, A.H.; Whittle, A.J. Analytical scaling relations to evaluate leakage and intrusion in intermittent water supply systems. PLoS ONE 2018, 13, e0196887. [Google Scholar] [CrossRef] [PubMed]
- Jalliffier-Verne, I.; Heniche, M.; Madoux-Humery, A.S.; Galarneau, M.; Servais, P.; Prévost, M.; Dorner, S. Cumulative effects of fecal contamination from combined sewer overflows: Management for source water protection. J. Environ. Manag. 2016, 174, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, M.; Tolouei, S.; Autixier, L.; Prévost, M.; Shakibaeinia, A.; Dorner, S. Normalized dynamic behavior of combined sewer overflow discharges for source water characterization and management. J. Environ. Manag. 2019, 249, 109386. [Google Scholar] [CrossRef] [PubMed]
- Tsvetanova, Z.; Tsvetkova, I.; Najdenski, H. Antimicrobial Resistance of Heterotrophic Bacteria in Drinking Water-Associated Biofilms. Water 2022, 14, 944. [Google Scholar] [CrossRef]
- Blyth, W.E.; Shahsavari, E.; Aburto-Medina, A.; Ball, A.S.; Osborn, A.M. Variation in the Structure and Composition of Bacterial Communities within Drinking Water Fountains in Melbourne, Australia. Water 2022, 14, 908. [Google Scholar] [CrossRef]
- Li, S.; Song, W.; Zhou, Y.; Tang, Y.; Gao, Y.; Miao, Z. Spread of extended-spectrum beta-lactamase-producing Escherichia coli from a swine farm to the receiving river. Environ. Sci. Pollut. Res. 2015, 22, 13033–13037. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Wang, L.; Peng, Q.; Li, Y.; Zhou, H.; Li, Q. Molecular characterization of extended-spectrum β-lactamase-producing multidrug resistant Escherichia coli from swine in Northwest China. Front. Microbiol. 2018, 9, 1756. [Google Scholar] [CrossRef]
- Yue, Z.; Zhang, J.; Zhou, Z.; Ding, C.; Wan, L.; Liu, J.; Chen, L.; Wang, X. Pollution characteristics of livestock faeces and the key driver of the spread of antibiotic resistance genes. J. Hazard. Mater. 2021, 409, 124957. [Google Scholar] [CrossRef]
- Durso, L.M.; Cook, K.L. Impacts of antibiotic use in agriculture: What are the benefits and risks? Curr. Opin. Microbiol. 2014, 19, 37–44. [Google Scholar] [CrossRef]
- Miller, D.N.; Jurgens, M.E.; Durso, L.M.; Schmidt, A.M. Simulated Winter Incubation of Soil With Swine Manure Differentially Affects Multiple Antimicrobial Resistance Elements. Front. Microbiol. 2020, 11, 611912. [Google Scholar] [CrossRef] [PubMed]
- Sabia, C.; Stefani, S.; Messi, P.; de Niederhäusern, S.; Bondi, M.; Condò, C.; Iseppi, R.; Anacarso, I. Extended-Spectrum Β-Lactamase and Plasmid-Mediated AMPC Genes in Swine and Ground Pork. J. Food Saf. 2017, 37, e12282. [Google Scholar] [CrossRef]
- Apostolakos, I.; Mughini-Gras, L.; Fasolato, L.; Piccirillo, A. Assessing the occurrence and transfer dynamics of ESBL/pAmpC-producing Escherichia coli across the broiler production pyramid. PLoS One 2019, 14, e0217174. [Google Scholar] [CrossRef] [PubMed]
- Dame-Korevaar, A.; Fischer, E.A.J.; van der Goot, J.; Stegeman, A.; Mevius, D. Transmission routes of ESBL/pAmpC producing bacteria in the broiler production pyramid, a literature review. Prev. Vet. Med. 2019, 162, 136–150. [Google Scholar] [CrossRef]
- Mir, R.A.; Weppelmann, T.A.; Teng, L.; Kirpich, A.; Elzo, M.A.; Driver, J.D.; Jeong, K.C. Colonization dynamics of cefotaxime resistant bacteria in beef cattle raised without cephalosporin antibiotics. Front. Microbiol. 2018, 9, 500. [Google Scholar] [CrossRef]
- Smith, T.C.; Male, M.J.; Harper, A.L.; Kroeger, J.S.; Tinkler, G.P.; Moritz, E.D.; Capuano, A.W.; Herwaldt, L.A.; Diekema, D.J. Methicillin-Resistant Staphylococcus aureus (MRSA) Strain ST398 Is Present in Midwestern U.S. Swine and Swine Workers. PLoS One 2009, 4, e4258. [Google Scholar] [CrossRef]
- Smith, T.C.; Gebreyes, W.A.; Abley, M.J.; Harper, A.L.; Forshey, B.M.; Male, M.J.; Martin, H.W.; Molla, B.Z.; Sreevatsan, S.; Thakur, S.; et al. Methicillin-Resistant Staphylococcus aureus in Pigs and Farm Workers on Conventional and Antibiotic-Free Swine Farms in the USA. PLoS One 2013, 8, e63704. [Google Scholar] [CrossRef]
- Van Cleef, B.A.G.L.; Broens, E.M.; Voss, A.; Huijsdens, X.W.; Züchner, L.; Van Benthem, B.H.B.; Kluytmans, J.A.J.W.; Mulders, M.N.; Van De Giessen, A.W. High prevalence of nasal MRSA carriage in slaughterhouse workers in contact with live pigs in the Netherlands. Epidemiol. Infect. 2010, 138, 756–763. [Google Scholar] [CrossRef]
- Smith, T.C.; Pearson, N. The emergence of Staphylococcus aureus ST398. Vector-Borne Zoonotic Dis. 2011, 11, 327–339. [Google Scholar] [CrossRef]
- United Nations. World Urbanization Prospects—Population Division—United Nations. Available online: https://population.un.org/wup/Download/ (accessed on 1 April 2021).
- Rowe, W.P.M.; Winn, M.D. Indexed variation graphs for efficient and accurate resistome profiling. Bioinformatics 2018, 34, 3601–3608. [Google Scholar] [CrossRef]
- He, Y.; Zhou, X.; Chen, Z.; Deng, X.; Gehring, A.; Ou, H.; Zhang, L.; Shi, X. PRAP: Pan Resistome analysis pipeline. BMC Bioinform. 2020, 21, 20. [Google Scholar] [CrossRef] [PubMed]
- Tamames, J.; Puente-Sánchez, F. SqueezeMeta, A. Highly Portable, Fully Automatic Metagenomic Analysis Pipeline. Front. Microbiol. 2019, 9, 3349. [Google Scholar] [CrossRef] [PubMed]
- Maruzani, R.; Canali, A.; Serafim, V.; Munoz, L.P.; Shah, A.J.; Perito, B.; Marvasi, M. Effect of anthropogenic pollution on the fitness of tetracycline sensitive Shigella flexneri in Thames river water. J. Environ. Chem. Eng. 2018, 6, 19–27. [Google Scholar] [CrossRef]
- Maruzani, R.; Pathak, A.; Ward, M.; Serafim, V.; Munoz, L.P.; Shah, A.J.; Marvasi, M. Antibiotic selective pressure in microcosms: Pollution influences the persistence of multidrug resistant Shigella flexneri 2a YSH6000 strain in polluted river water samples. Environ. Technol. Innov. 2020, 19, 100821. [Google Scholar] [CrossRef]
- Manaia, C.M. Assessing the Risk of Antibiotic Resistance Transmission from the Environment to Humans: Non-Direct Proportionality between Abundance and Risk. Trends Microbiol. 2017, 25, 173–181. [Google Scholar] [CrossRef] [PubMed]
- United Nations. Urban and Rural Populations; United Nations: New York, NY, USA, 2018. [Google Scholar]
- European Medicines Agency. Assessment of the Risk to Public Health from Antimicrobial Resistance due to the Use of an Antimicrobial Veterinary Medicinal Product in Food-Producing Animals. Available online: https://www.ema.europa.eu/en/assessment-risk-public-health-antimicrobial-resistance-due-use-antimicrobial-veterinary-medicinal (accessed on 1 April 2021).
- Ben, Y.; Fu, C.; Hu, M.; Liu, L.; Wong, M.H.; Zheng, C. Human health risk assessment of antibiotic resistance associated with antibiotic residues in the environment: A review. Environ. Res. 2019, 169, 483–493. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; ISBN 9781400881376. [Google Scholar]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-L.; An, X.-L.; Zheng, B.-X.; Gillings, M.; Peñuelas, J.; Cui, L.; Su, J.-Q.; Zhu, Y.-G. Loss of soil microbial diversity exacerbates spread of antibiotic resistance. Soil Ecol. Lett. 2019, 1, 3–13. [Google Scholar] [CrossRef]
- Case, T.J. Invasion resistance arises in strongly interacting species-rich model competition communities. Proc. Natl. Acad. Sci. USA 1990, 87, 9610–9614. [Google Scholar] [CrossRef]
- Barberán, A.; Casamayor, E.O.; Fierer, N. The microbial contribution to macroecology. Front. Microbiol. 2014, 5, 203. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, K.C.; Karaoz, U.; Hanson, C.A.; Santee, C.A.; Bradford, M.A.; Treseder, K.K.; Wallenstein, M.D.; Brodie, E.L. Differential growth responses of soil bacterial taxa to carbon substrates of varying chemical recalcitrance. Front. Microbiol. 2011, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Eisenhauer, N.; Scheu, S.; Jousset, A. Increasing antagonistic interactions cause bacterial communities to collapse at high diversity. Ecol. Lett. 2012, 15, 468–474. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
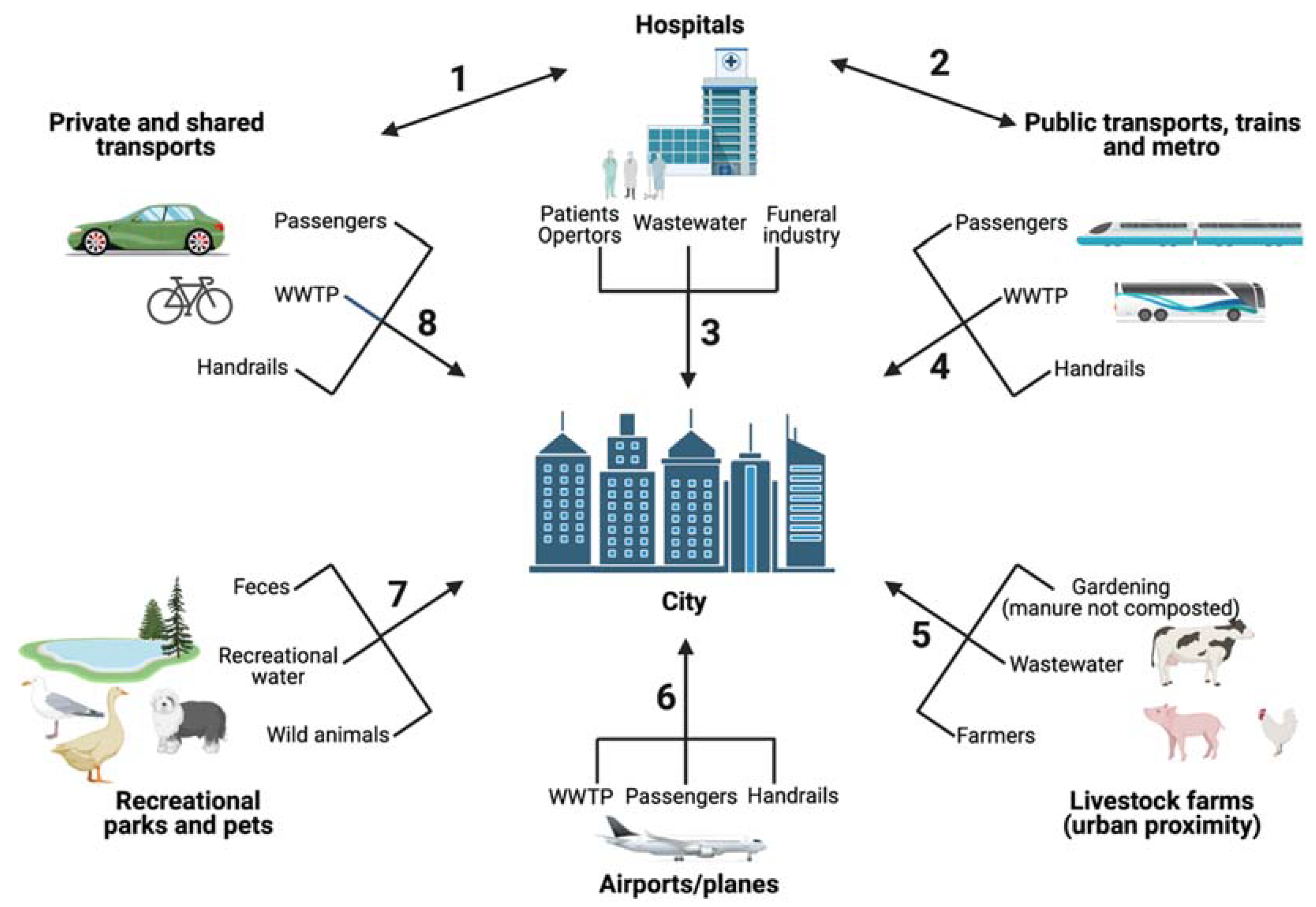
| Route 1 | References |
|---|---|
| 1 | [43] |
| 2 | [35,36] |
| 3 | [12,20,21,22,25,26,28,29] |
| 4 | [34,37,38,39,40] |
| 5 | [100,101,102,103,104,105,106,107,108,109,110,111,112] |
| 6 | [30,31,32,33] |
| 7 | [10,20,63,65,66,67,68,69,72,73,74,75,51,76,77,78,79,80,81,82,83,84,52,53,54,55,56,61,62] |
| 8 | [41,42,43,44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassallo, A.; Kett, S.; Purchase, D.; Marvasi, M. The Bacterial Urban Resistome: Recent Advances. Antibiotics 2022, 11, 512. https://doi.org/10.3390/antibiotics11040512
Vassallo A, Kett S, Purchase D, Marvasi M. The Bacterial Urban Resistome: Recent Advances. Antibiotics. 2022; 11(4):512. https://doi.org/10.3390/antibiotics11040512
Chicago/Turabian StyleVassallo, Alberto, Steve Kett, Diane Purchase, and Massimiliano Marvasi. 2022. "The Bacterial Urban Resistome: Recent Advances" Antibiotics 11, no. 4: 512. https://doi.org/10.3390/antibiotics11040512
APA StyleVassallo, A., Kett, S., Purchase, D., & Marvasi, M. (2022). The Bacterial Urban Resistome: Recent Advances. Antibiotics, 11(4), 512. https://doi.org/10.3390/antibiotics11040512