
Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs



Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of Essential Oils
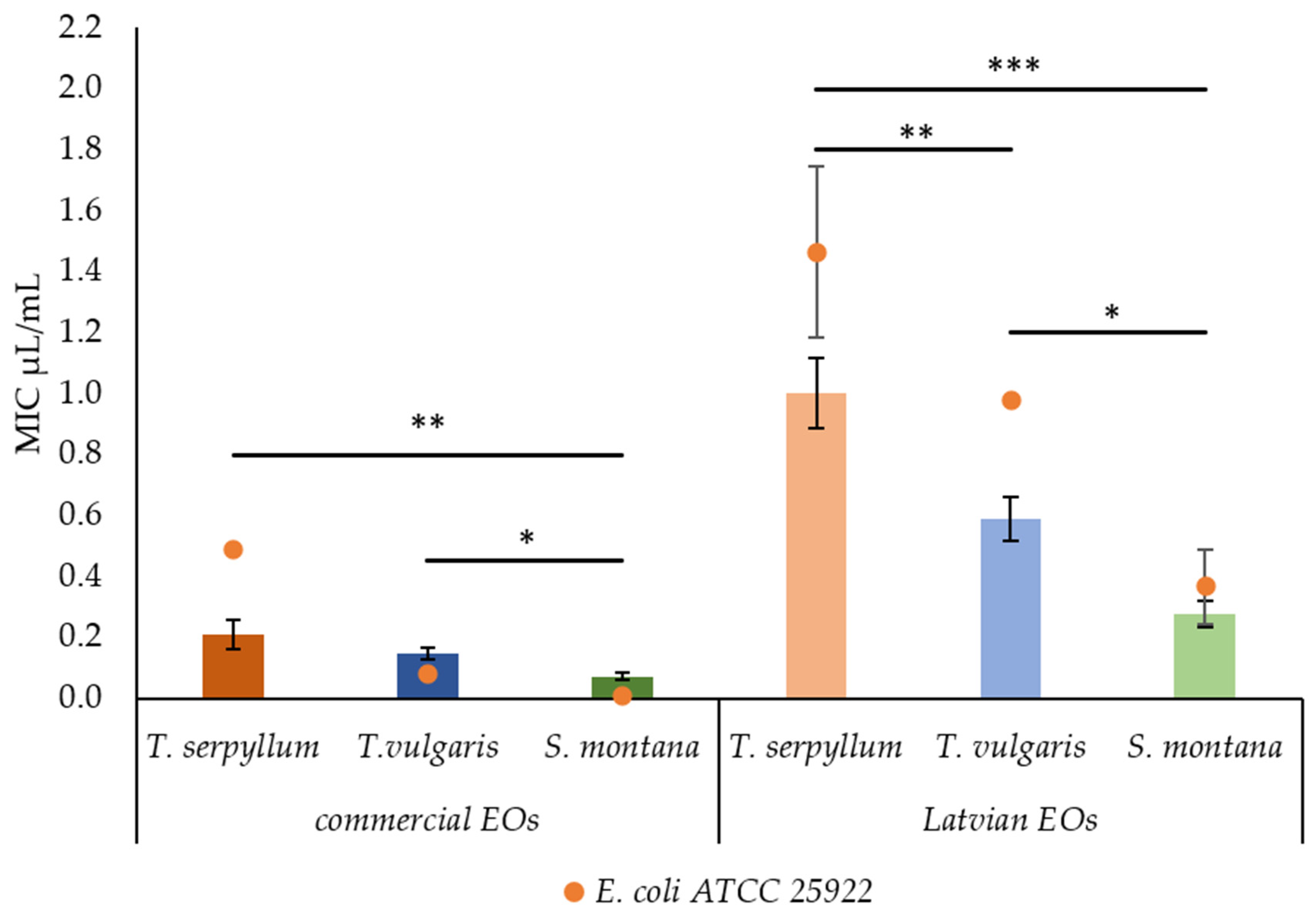
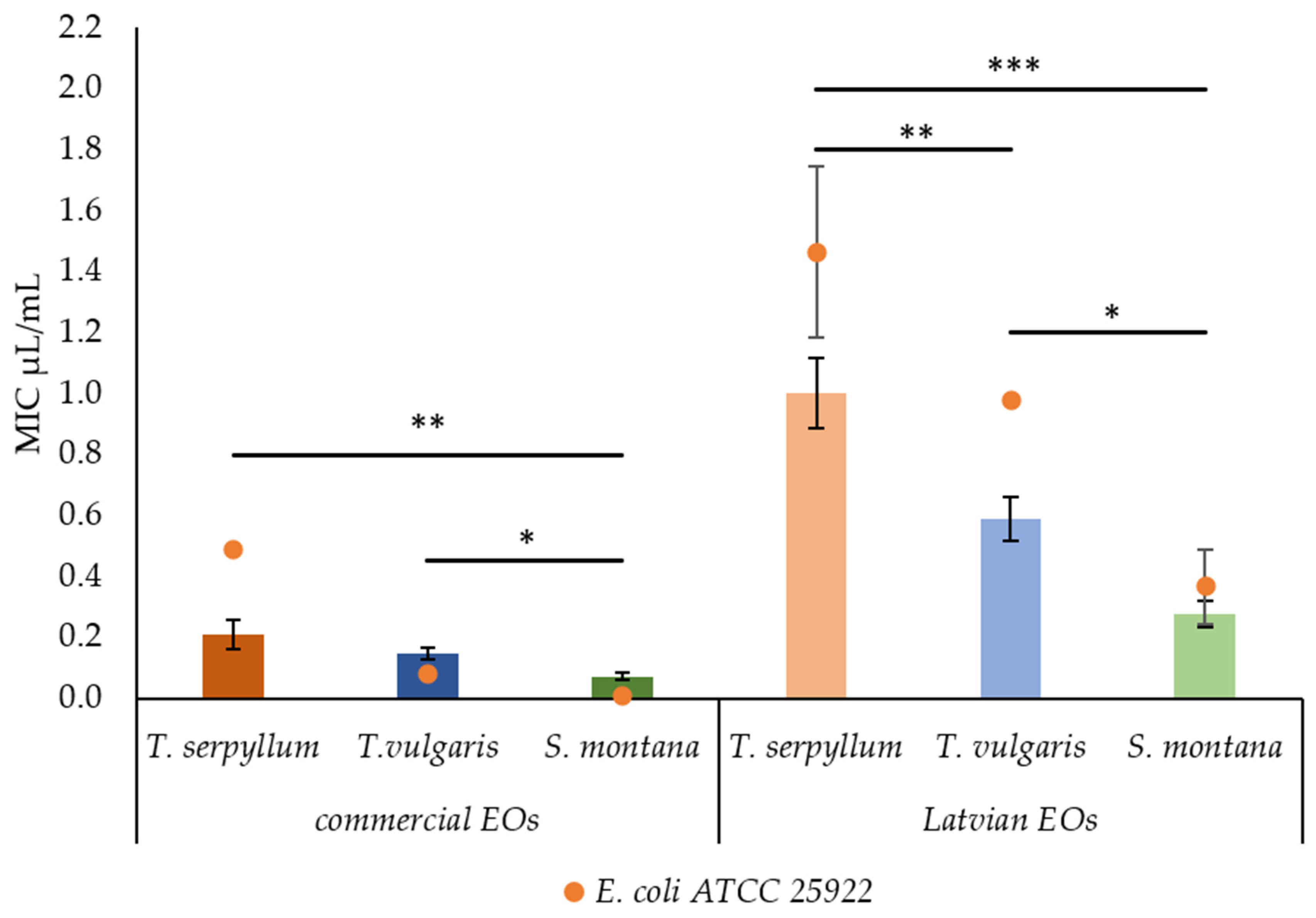
2.2. Antibacterial Activity of Commercial and Latvian EOs
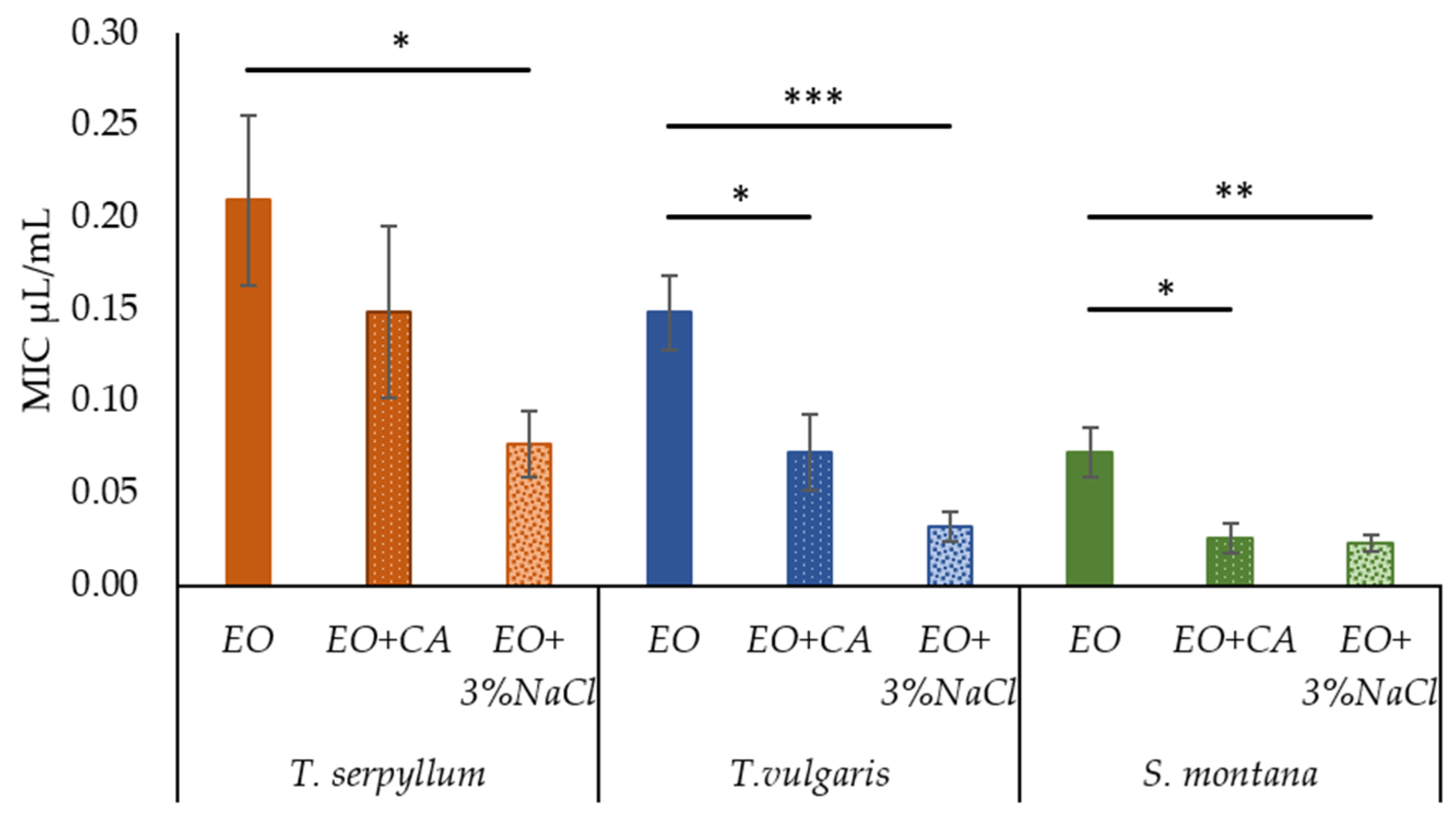
2.3. Effect of Supplementation of Commercial EOs with Caprylic Acid and Sodium Chloride on Antibacterial Activity against Faecal E. coli
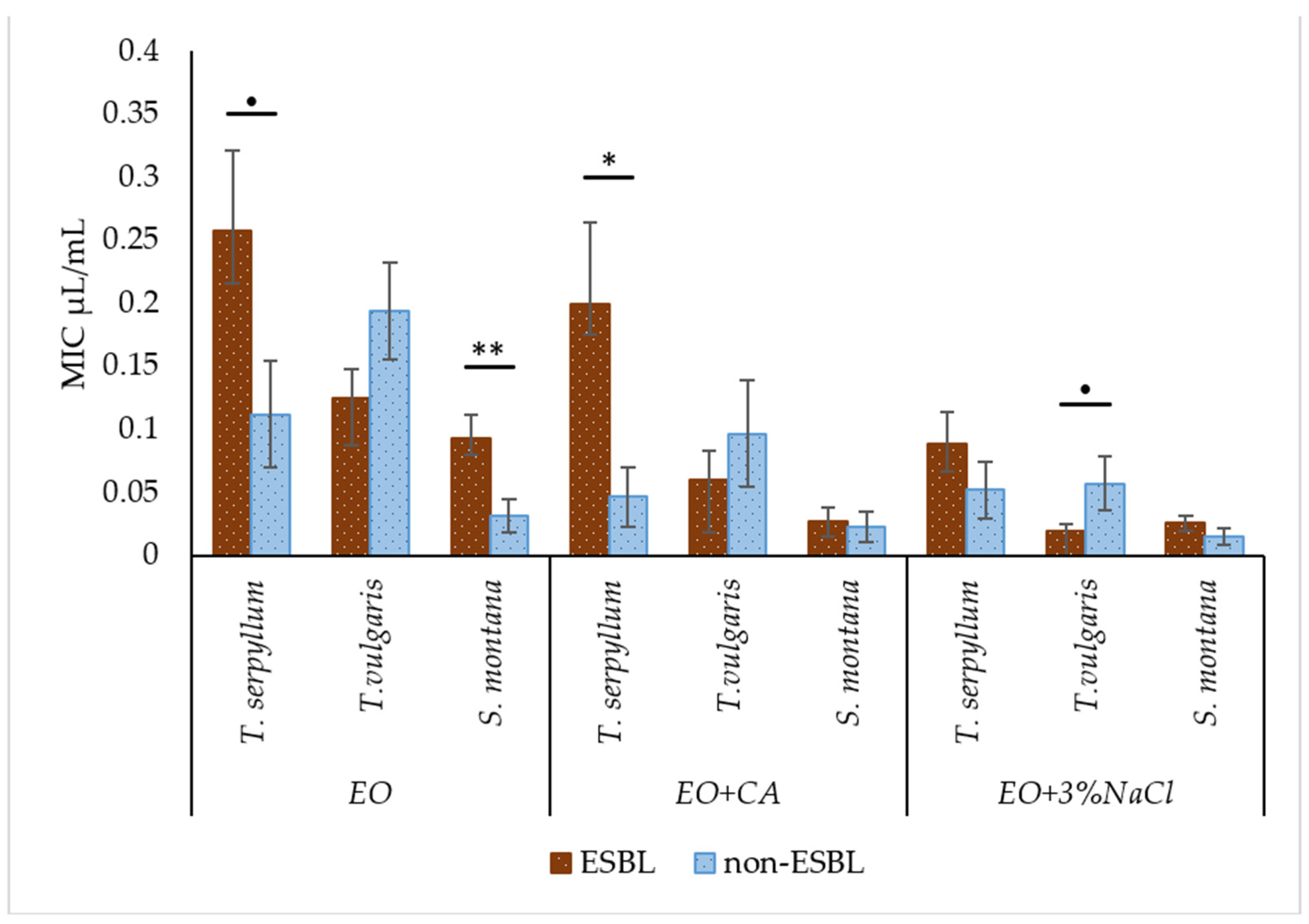
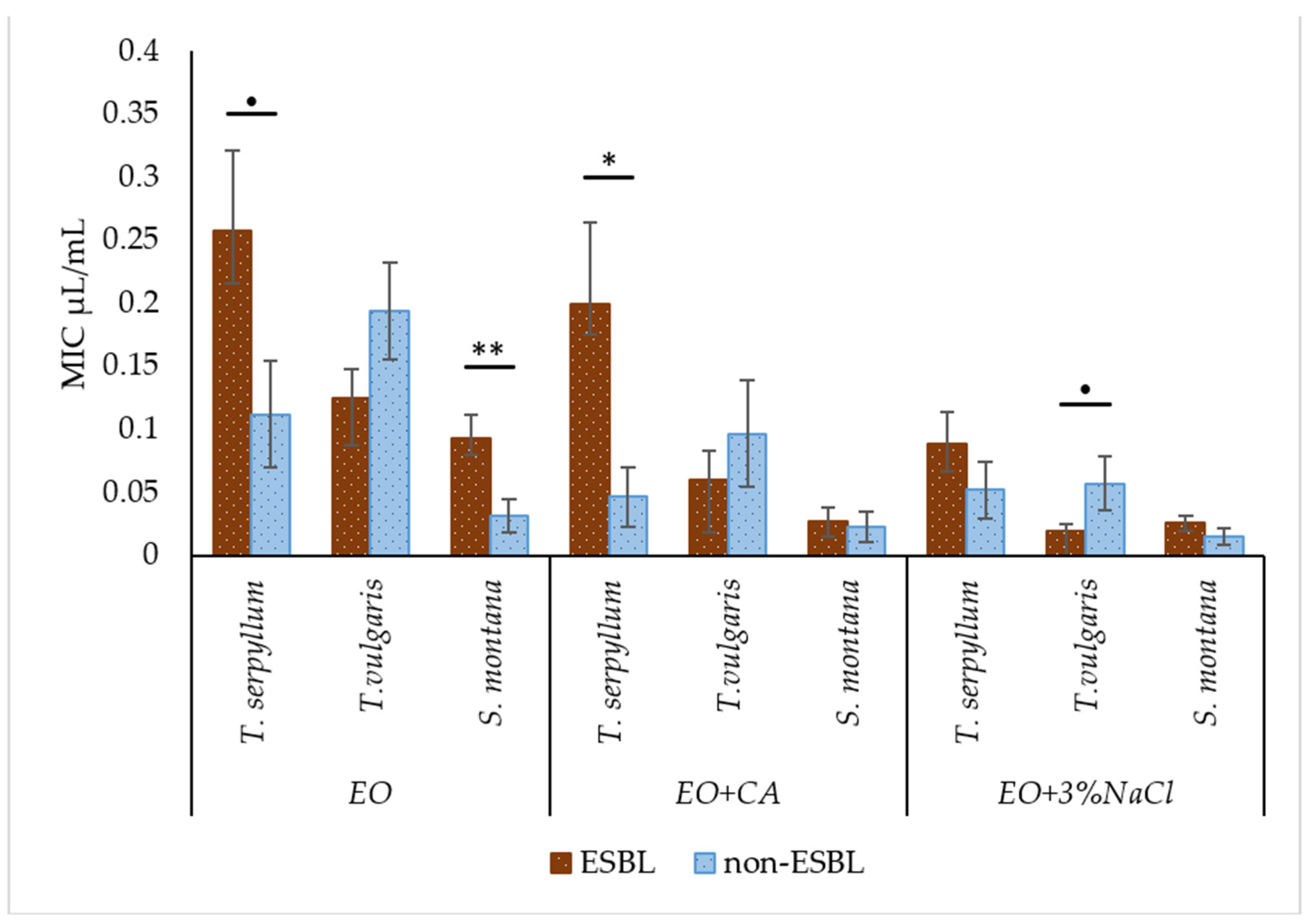
2.4. Comparison of Antibacterial Activity of Essential Oils with and without Supplementation against Faecal ESBL-Producing and Non-ESBL-Producing E. coli
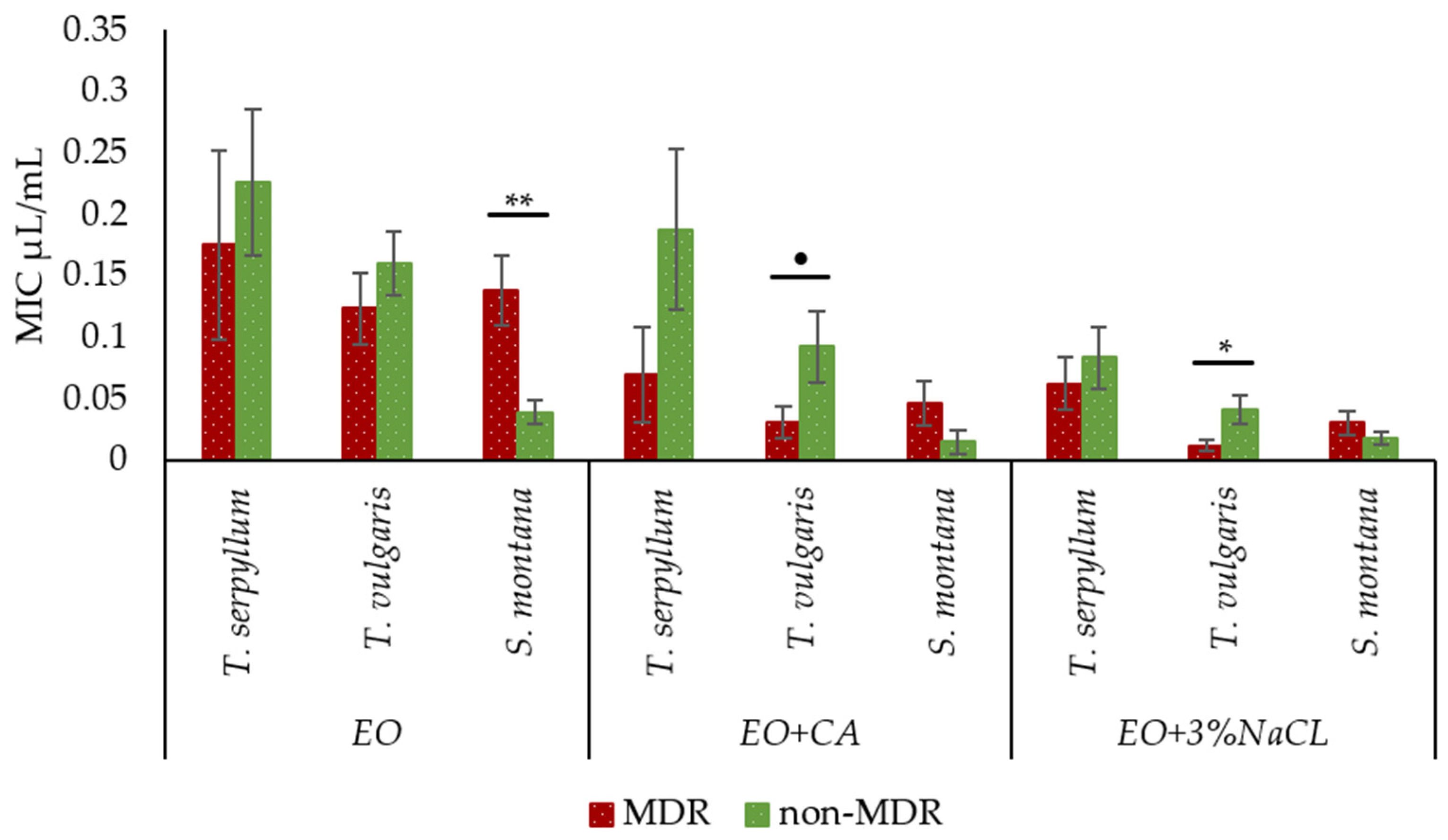
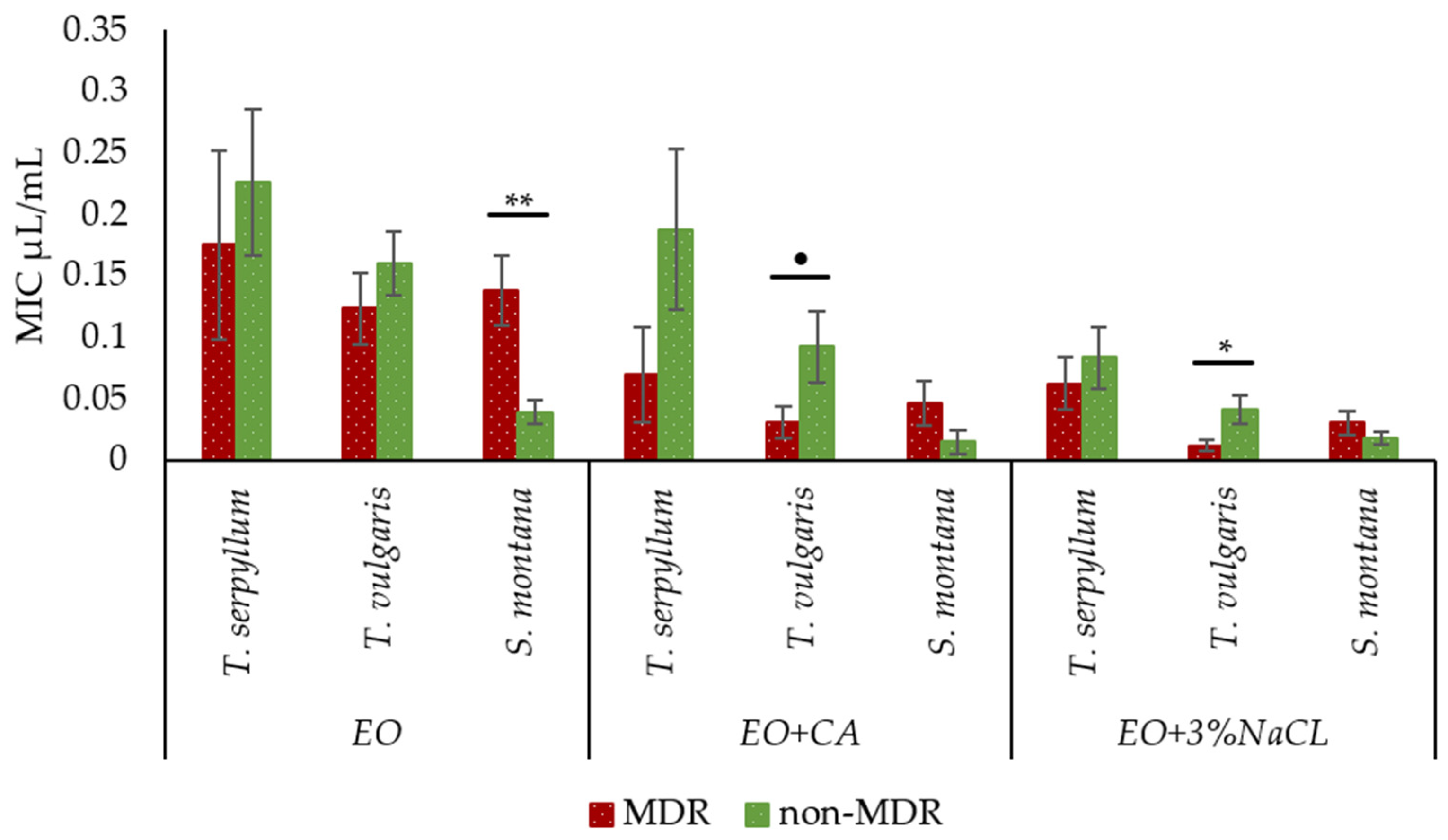
2.5. Comparison of Antibacterial Activity of Essential Oils with and without Supplementation against Faecal MDR and Non-MDR E. coli
3. Discussion
4. Materials and Methods
4.1. Essential Oils (EOs)
4.1.1. Authors-Obtained Essential Oils
4.1.2. Commercial Essential Oils
4.1.3. Preparation of Essential Oils for Analysis of Aroma Volatiles by Gas Chromatography–Mass Spectrometry
4.2. Characterisation of Essential Oils Chemical Composition of by Gas Chromatography-Mass Spectrometry
4.3. Antibacterial Assays
4.3.1. Escherichia coli Strains
4.3.2. Determination of Minimum Inhibitory Concentration (MIC)
4.3.3. Determination of Minimum Bactericidal Concentration (MBC)
4.3.4. Determination of the Effects of EOs Supplementation with NaCl and Caprylic Acid on MICs and MBCs
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramos, S.; Silva, V.; Dapkevicius, M.d.L.E.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, 2239. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, A.; Prasad, J.; Das, S.; Dwivedy, A.K. Essential Oils and Their Application in Food Safety. Front. Sustain. Food Syst. 2021, 5, 133. [Google Scholar] [CrossRef]
- Zhai, H.; Liu, H.; Wang, S.; Wu, J.; Kluenter, A.M. Potential of Essential Oils for Poultry and Pigs. Anim. Nutr. 2018, 4, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Khoury, M.; Stien, D.; Eparvier, V.; Ouaini, N.; El Beyrouthy, M. Report on the Medicinal Use of Eleven Lamiaceae Species in Lebanon and Rationalization of Their Antimicrobial Potential by Examination of the Chemical Composition and Antimicrobial Activity of Their Essential Oils. Evid. Based Complement. Altern. Med. 2016, 2016, 2547169. [Google Scholar] [CrossRef] [Green Version]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial Agents from Plants: Antibacterial Activity of Plant Volatile Oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Ben Arfa, A.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier, P. Antimicrobial Activity of Carvacrol Related to Its Chemical Structure. Lett. Appl. Microbiol. 2006, 43, 149–154. [Google Scholar] [CrossRef]
- Ultee, A.; Bennik, M.H.J.; Moezelaar, R. The Phenolic Hydroxyl Group of Carvacrol Is Essential for Action against the Food-Borne Pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Radenkovs, V.; Kviesis, J.; Juhnevica-Radenkova, K.; Valdovska, A.; Püssa, T.; Klavins, M.; Drudze, I. Valorization of Wild Apple (Malus Spp.) By-Products as a Source of Essential Fatty Acids, Tocopherols and Phytosterols with Antimicrobial Activity. Plants 2018, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial Fatty Acids: An Update of Possible Mechanisms of Action and Implications in the Development of the next-Generation of Antibacterial Agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial Free Fatty Acids: Activities, Mechanisms of Action and Biotechnological Potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanten, G.J.A.; Calder, P.C. Immune Modulation by Parenteral Lipid Emulsions. Am. J. Clin. Nutr. 2007, 85, 1171–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayyan, M.; Devlieger, H.; Jochum, F.; Allegaert, K. Short-Term Use of Parenteral Nutrition With a Lipid Emulsion Containing a Mixture of Soybean Oil, Olive Oil, Medium-Chain Triglycerides, and Fish Oil. J. Parenter. Enter. Nutr. 2012, 36, 81S–94S. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Rhee, M. Synergistic Antimicrobial Activity of Caprylic Acid in Combination with Citric Acid against Both Escherichia coli O157:H7 and Indigenous Microflora in Carrot Juice. Food Microbiol. 2015, 49, 166–172. [Google Scholar] [CrossRef]
- Rosenblatt, J.; Reitzel, R.A.; Vargas-Cruz, N.; Chaftari, A.M.; Hachem, R.; Raad, I. Caprylic and Polygalacturonic Acid Combinations for Eradication of Microbial Organisms Embedded in Biofilm. Front. Microbiol. 2017, 8, 1999. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Singh, H.; Ram, C. Efficacy and Mechanism of Carvacrol with Octanoic Acid against Mastitis Causing Multi-Drug-Resistant Pathogens. Braz. J. Microbiol. 2022, 53, 385–399. [Google Scholar] [CrossRef]
- Henney, J.E.; Taylor, C.L.; Boon, C.S. Preservation and Physical Property Roles of Sodium in Foods. In Strategies to Reduce Sodium Intake in the United States; Henney, J.E., Taylor, C.L., Boon, C.S., Eds.; National Academies Press: Washington, DC, USA, 2010; pp. 91–108. ISBN 978-0-309-15588-5. [Google Scholar]
- Xu, Y.; You, G.; Zhang, M.; Peng, D.; Jiang, Z.; Qi, S.; Yang, S.; Hou, J. Antibiotic Resistance Genes Alternation in Soils Modified with Neutral and Alkaline Salts: Interplay of Salinity Stress and Response Strategies of Microbes. Sci. Total Environ. 2022, 809, 152246. [Google Scholar] [CrossRef]
- Tan, L.; Wang, F.; Liang, M.; Wang, X.; Das, R.; Mao, D.; Luo, Y. Antibiotic Resistance Genes Attenuated with Salt Accumulation in Saline Soil. J. Hazard. Mater. 2019, 374, 35–42. [Google Scholar] [CrossRef]
- Willing, B.P.; Pepin, D.M.; Marcolla, C.S.; Forgie, A.J.; Diether, N.E.; Bourrie, B.C.T. Bacterial Resistance to Antibiotic Alternatives: A Wolf in Sheep’s Clothing? Anim. Front. 2018, 8, 39–47. [Google Scholar] [CrossRef]
- Nieto, G. Biological Activities of Three Essential Oils of the Lamiaceae Family. Medicines 2017, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Kovačević, Z.; Radinović, M.; Čabarkapa, I.; Kladar, N.; Božin, B. Natural Agents against Bovine Mastitis Pathogens. Antibiotics 2021, 10, 205. [Google Scholar] [CrossRef]
- Nikolić, M.; Glamočlija, J.; Ferreira, I.C.F.R.; Calhelha, R.C.; Fernandes, Â.; Marković, T.; Marković, D.; Giweli, A.; Soković, M. Chemical Composition, Antimicrobial, Antioxidant and Antitumor Activity of Thymus serpyllum L., Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. Essential Oils. Ind. Crops Prod. 2014, 52, 183–190. [Google Scholar] [CrossRef]
- Galovičová, L.; Borotová, P.; Valková, V.; Vukovic, N.L.; Vukic, M.; Terentjeva, M.; Štefániková, J.; Ďúranová, H.; Kowalczewski, P.Ł.; Kačániová, M. Thymus serpyllum Essential Oil and Its Biological Activity as a Modern Food Preserver. Plants 2021, 10, 1416. [Google Scholar] [CrossRef]
- Wesolowska, A.; Grzeszczuk, M.; Jadczak, D.; Nawrotek, P.; Struk, M. Comparison of the Chemical Composition and Antimicrobial Activity of Thymus serpyllum Essential Oils. Not. Bot. Horti Agrobot. Cluj Napoca 2015, 43, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Ložionė, K.; Venskutonis, P.R. Chemical Composition of the Essential Oil of Thymus serpyllum L. ssp. serpyllum Growing Wild in Lithuania. J. Essent. Oil Res. 2006, 18, 206–211. [Google Scholar] [CrossRef]
- Raal, A.; Paaver, U.; Arak, E.; Orav, A. Content and Composition of the Essential Oil of Thymus serpyllum L. Growing Wild in Estonia. Medicina 2004, 40, 795–800. [Google Scholar]
- Paaver, U.; Orav, A.; Arak, E.; Mäeorg, U.; Raal, A. Phytochemical Analysis of the Essential Oil of Thymus serpyllum L. Growing Wild in Estonia. Nat. Prod. Res. 2008, 22, 108–115. [Google Scholar] [CrossRef]
- Galovičová, L.; Borotová, P.; Valková, V.; Vukovic, N.L.; Vukic, M.; Štefániková, J.; Ďúranová, H.; Kowalczewski, P.Ł.; Čmiková, N.; Kačániová, M. Thymus vulgaris Essential Oil and Its Biological Activity. Plants 2021, 10, 1959. [Google Scholar] [CrossRef]
- Galambosi, B.; Rey, C.; Vouillamoz, J.F. Suitability of Swiss Herb Cultivars under Finnish Climatic Conditions. Acta Hortic. 2010, 860, 173–180. [Google Scholar] [CrossRef]
- Navarro-Rocha, J.; Andrés, M.F.; Díaz, C.E.; Burillo, J.; González-Coloma, A. Composition and Biocidal Properties of Essential Oil from Pre-Domesticated Spanish Satureja montana. Ind. Crops Prod. 2020, 145, 111958. [Google Scholar] [CrossRef]
- Vitanza, L.; Maccelli, A.; Marazzato, M.; Scazzocchio, F.; Comanducci, A.; Fornarini, S.; Crestoni, M.E.; Filippi, A.; Fraschetti, C.; Rinaldi, F.; et al. Satureja montana L. Essential Oil and Its Antimicrobial Activity Alone or in Combination with Gentamicin. Microb. Pathog. 2019, 126, 323–331. [Google Scholar] [CrossRef]
- Šojić, B.; Pavlić, B.; Tomović, V.; Ikonić, P.; Zeković, Z.; Kocić-Tanackov, S.; Đurović, S.; Škaljac, S.; Jokanović, M.; Ivić, M. Essential Oil versus Supercritical Fluid Extracts of Winter Savory (Satureja montana L.)—Assessment of the Oxidative, Microbiological and Sensory Quality of Fresh Pork Sausages. Food Chem. 2019, 287, 280–286. [Google Scholar] [CrossRef]
- Slavkovska, V.; Jancic, R.; Bojovic, S.; Milosavljevic, S.; Djokovic, D. Variability of Essential Oils of Satureja montana L. and Satureja Kitaibelii Wierzb. Ex Heuff. from the Central Part of the Balkan Peninsula. Phytochemistry 2001, 57, 71–76. [Google Scholar] [CrossRef]
- Ibraliu, A.; Dhillon, B.S.; Faslia, N.; Stich, B. Variability of Essential Oil Composition in Albanian Accessions of Satureja montana L. J. Med. Plants Res. 2010, 4, 1359–1364. [Google Scholar]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Cox, S.D.; Mann, C.M.; Markham, J.L. Interactions between Components of the Essential Oil of Melaleuca alternifolia. J. Appl. Microbiol. 2001, 91, 492–497. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Assessment of Factors Influencing Antimicrobial Activity of Carvacrol and Cymene against Vibrio Cholerae in Food. J. Biosci. Bioeng. 2010, 110, 614–619. [Google Scholar] [CrossRef]
- Kang, L.; Yap, C.W.; Lim, P.F.C.; Chen, Y.Z.; Ho, P.C.; Chan, Y.W.; Wong, G.P.; Chan, S.Y. Formulation Development of Transdermal Dosage Forms: Quantitative Structure-Activity Relationship Model for Predicting Activities of Terpenes That Enhance Drug Penetration through Human Skin. J. Control. Release 2007, 120, 211–219. [Google Scholar] [CrossRef]
- Cosentino, S.; Tuberoso, C.I.G.; Pisano, B.; Satta, M.; Mascia, V.; Arzedi, E.; Palmas, F. In-Vitro Antimicrobial Activity and Chemical Composition of Sardinian Thymus Essential Oils. Lett. Appl. Microbiol. 1999, 29, 130–135. [Google Scholar] [CrossRef]
- Gallucci, M.N.; Oliva, M.M.; Casero, C.N.; Dambolena, J.S.; Luna, A.; Zygadlo, J.; Demo, M.S. Antimicrobial Combined Action of Terpenes against the Food_borne Microorganisms Escherichia coli, Staphylococcus aureus and Bacillus cereus. Flavour Fragr. J. 2009, 24, 348–354. [Google Scholar] [CrossRef]
- Hulánková, R.; Bořilová, G. In Vitro Combined Effect of Oregano Essential Oil and Caprylic Acid against Salmonella Serovars, Escherichia coli O157:H7, Staphylococcus aureus and Listeria monocytogenes. Acta Vet. Brno 2011, 80, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.A.; Rhee, M.S. Highly Enhanced Bactericidal Effects of Medium Chain Fatty Acids (Caprylic, Capric, and Lauric Acid) Combined with Edible Plant Essential Oils (Carvacrol, Eugenol, β-Resorcylic Acid, Trans-Cinnamaldehyde, Thymol, and Vanillin) against Escherichia coli O157:H7. Food Control. 2016, 60, 447–454. [Google Scholar]
- Yoon, B.; Jackman, J.; Valle-González, E.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.H.; Kim, H.W.; Moon, H.; Rhee, M.S. Sodium Chloride Significantly Enhances the Bactericidal Actions of Carvacrol and Thymol against the Halotolerant Species Escherichia coli O157:H7, Listeria monocytogenes, and Staphylococcus aureus. LWT 2020, 122, 109015. [Google Scholar] [CrossRef]
- Hajmeer, M.; Ceylan, E.; Marsden, J.L.; Fung, D.Y.C. Impact of Sodium Chloride on Escherichia coli O157:H7 and Staphylococcus aureus Analysed Using Transmission Electron Microscopy. Food Microbiol. 2006, 23, 446–452. [Google Scholar] [CrossRef]
- Wood, J.M. Bacterial Responses to Osmotic Challenges. J. Gen. Physiol. 2015, 145, 381–388. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. Bacterial Osmoadaptation: The Role of Osmolytes in Bacterial Stress and Virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Benameur, Q.; Gervasi, T.; Pellizzeri, V.; Pľuchtovà, M.; Tali-Maama, H.; Assaous, F.; Guettou, B.; Rahal, K.; Gruľovà, D.; Dugo, G.; et al. Antibacterial Activity of Thymus vulgaris Essential Oil Alone and in Combination with Cefotaxime against BlaESBL Producing Multidrug Resistant Enterobacteriaceae Isolates. Nat. Prod. Res. 2019, 33, 2647–2654. [Google Scholar] [CrossRef]
- Pagès, J.-M.; James, C.E.; Winterhalter, M. The Porin and the Permeating Antibiotic: A Selective Diffusion Barrier in Gram-Negative Bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, S.M.; Abd El-Aziz, A.M.; Hassan, R.; Abdelmegeed, E.S. β-Lactam Resistance Associated with β-Lactamase Production and Porin Alteration in Clinical Isolates of E. coli and K. pneumoniae. PLoS ONE 2021, 16, e0251594. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial Multidrug Efflux Pumps: Mechanisms, Physiology and Pharmacological Exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisluk, G.; Kalily, E.; Yaron, S. Resistance to Essential Oils Affects Survival of Salmonella enterica Serovars in Growing and Harvested Basil. Environ. Microbiol. 2013, 15, 2787–2798. [Google Scholar] [PubMed]
- Kalily, E.; Hollander, A.; Korin, B.; Cymerman, I.; Yaron, S. Mechanisms of Resistance to Linalool in Salmonella Senftenberg and Their Role in Survival on Basil. Environ. Microbiol. 2016, 18, 3673–3688. [Google Scholar] [CrossRef] [PubMed]
- Kalily, E.; Hollander, A.; Korin, B.; Cymerman, I.; Yaron, S. Adaptation of Salmonella enterica Serovar Senftenberg to Linalool and Its Association with Antibiotic Resistance and Environmental Persistence. Appl. Environ. Microbiol. 2017, 83, e03398-16. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Effects of Melaleuca alternifolia (Tea Tree) Essential Oil and the Major Monoterpene Component Terpinen-4-Ol on the Development of Single- and Multistep Antibiotic Resistance and Antimicrobial Susceptibility. Antimicrob. Agents Chemother. 2012, 56, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Becerril, R.; Nerín, C.; Gómez-Lus, R. Evaluation of Bacterial Resistance to Essential Oils and Antibiotics After Exposure to Oregano and Cinnamon Essential Oils. Foodborne Pathog. Dis. 2012, 9, 699–705. [Google Scholar] [CrossRef]
- Bezerra Filho, C.M.; da Silva, L.C.N.; da Silva, M.V.; Løbner-Olesen, A.; Struve, C.; Krogfelt, K.A.; Dos Santos Correia, M.T.; Vilela Oliva, M.L. Antimicrobial and Antivirulence Action of Eugenia brejoensis Essential Oil In Vitro and In Vivo Invertebrate Models. Front. Microbiol. 2020, 11, 424. [Google Scholar] [CrossRef] [Green Version]
- Al-Kandari, F.; Al-Temaimi, R.; van Vliet, A.H.M.; Woodward, M.J. Thymol Tolerance in Escherichia coli Induces Morphological, Metabolic and Genetic Changes. BMC Microbiol. 2019, 19, 294. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Xiong, X.-S.; Yang, Y.-Y.; Wang, J.-J.; Wang, M.-M.; Tang, J.-W.; Liu, Q.-H.; Wang, L.; Gu, B. Effects of NaCl Concentrations on Growth Patterns, Phenotypes Associated With Virulence, and Energy Metabolism in Escherichia coli BW25113. Front. Microbiol. 2021, 12, 705326. [Google Scholar] [CrossRef]
- Hou, H.-S.; Bonku, E.M.; Zhai, R.; Zeng, R.; Hou, Y.-L.; Yang, Z.-H.; Quan, C. Extraction of Essential Oil from Citrus Reticulate Blanco Peel and Its Antibacterial Activity against Cutibacterium acnes (Formerly Propionibacterium acnes). Heliyon 2019, 5, e02947. [Google Scholar] [CrossRef] [Green Version]
- Tabanca, N.; Demirci, B.; Baser, K.H.C.; Aytac, Z.; Ekici, M.; Khan, S.I.; Jacob, M.R.; Wedge, D.E. Chemical Composition and Antifungal Activity of Salvia macrochlamys and Salvia recognita Essential Oils. J. Agric. Food Chem. 2006, 54, 6593–6597. [Google Scholar] [CrossRef] [PubMed]
- Keita, S.M.; Vincent, C.; Schmit, J.-P.; Bélanger, A. Essential Oil Composition of Ocimum basilicum L., O. gratissimum L. and O. suave L. in the Republic of Guinea. Flavour Fragr. J. 2000, 15, 339–341. [Google Scholar] [CrossRef]
- Iwanami, Y.; Tateba, H.; Kodama, N.; Kishino, K. Changes of Lemon Flavor Components in an Aqueous Solution during UV Irradiation. J. Agric. Food Chem. 1997, 45, 463–466. [Google Scholar] [CrossRef]
- Ngassoum, M.B.; Yonkeu, S.; Jirovetz, L.; Buchbauer, G.; Schmaus, G.; Hammerschmidt, F.-J. Chemical Composition of Essential Oils of Lantana camara Leaves and Flowers from Cameroon and Madagascar. Flavour Fragr. J. 1999, 14, 245–250. [Google Scholar] [CrossRef]
- Njoroge, S.M.; Ukeda, H.; Kusunose, H.; Sawamura, M. Japanese SourCitrus Fruits. Part IV. Volatile Compounds of Naoshichi and Tahiti Lime Essential Oils. Flavour Fragr. J. 1996, 11, 25–29. [Google Scholar] [CrossRef]
- Mjøs, S.A. Two-Dimensional Fatty Acid Retention Indices. J. Chromatogr. A 2004, 1061, 201–209. [Google Scholar] [CrossRef]
- Gāliņa, D.; Balins, A.; Valdovska, A. The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia. Antibiotics 2021, 10, 1099. [Google Scholar] [CrossRef]
- Mandrika, I.; Kumar, S.; Zandersone, B.; Eranezhath, S.S.; Petrovska, R.; Liduma, I.; Jezupovs, A.; Pirags, V.; Tracevska, T. Antibacterial and Anti-Inflammatory Potential of Polyherbal Formulation Used in Chronic Wound Healing. Evid. Based Complement. Altern. Med. 2021, 2021, 9991454. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RI | Compound Name | Composition, mg/kg ± SD, (%) | |||||
|---|---|---|---|---|---|---|---|
| Thymus vulgaris | Satureja montana | Thymus serpyllum | |||||
| C | LV | C | LV | C | LV | ||
| 1015 | cis-p-menthane 1 | 4.1 ± 0.1 (0.16) | - | - | - | - | - |
| 1022 | α-pinene 1 | - | - | 32.2 ± 0.5 (0.63) | 50.7 ± 3.9 (0.99) | 56.9 ± 3.0 (1.30) | 20.8 ± 0.6 (0.35) |
| 1023 | β-terpinen 1 | 92.5 ± 4.6 (3.60) | 21.4 ± 1.2 (0.87) | - | - | - | - |
| 1027 | α-thujene 1 | 3.6 ± 0.2 (0.14) | 35.8 ± 1.4 (1.46) | 15.4 ± 0.5 (0.30) | 99.6 ± 8.4 (1.94) | 137.5 ± 12.6 (3.14) | 40.9 ± 3.0 (0.70) |
| 1039 | trans-p-menthane 1 | 4.2 ± 0.3 (0.17) | - | - | - | - | - |
| 1058 | α-fenchene 1 | 10.1 ± 0.5 (0.39) | - | - | - | - | - |
| 1065 | camphene 1 | 49.8 ± 2.0 (1.94) | 15.9 ± 0.7 (0.65) | 30.5 ± 0.6 (0.60) | 20.0 ± 1.8 (0.39) | 13.9 ± 0.7 (0.32) | 17.9 ± 0.9 (0.30) |
| 1107 | β-pinene 1 | 12.6 ± 0.9 (0.49) | 9.3 ± 0.6 (0.38) | 9.4 ± 0.0 (0.19) | 16.7 ± 1.2 (0.32) | 17.1 ± 0.9 (0.39) | 12.1 ± 0.6 (0.21) |
| 1122 | sabinene 1 | - | 5.8 ± 0.4 (0.24) | - | 11.4 ± 0.4 (0.22) | 4.8 ± 0.2 (0.11) | 70.1 ± 3.0 (1.19) |
| 1132 | dehydrosabinene 1 | - | - | t | t | t | - |
| 1136 | carvomenthene 1 | 6.8 ± 0.3 (0.26) | - | - | - | - | - |
| 1149 | δ-3-carene 1 | - | 4.9 ± 0.4 (0.24) | t | 7.5 ± 0.5 (0.15) | 9.9 ± 1.1 (0.23) | t |
| 1165 | β-myrcene + α-phellandrene 1 | 38.8 ± 3.4 (1.51) | 54.3 ± 2.4 (0.20) | 58.0 ± 1.3 (1.14) | 172.8 ± 15.2 (3.36) | 188.0 ± 17.7 (4.30) | 201.7 ± 15.6 (3.43) |
| 1170 | pseudolimonene 1 | 4.4 ± 0.4 (0.17) | - | - | - | - | - |
| 1181 | α-terpinene 1 | 11.2 ± 0.5 (0.44) | 40.6 ± 2.2 (1.66) | 70.3 ± 0.4 (1.39) | 142.8 ± 11.0 (2.78) | 96.8 ± 7.3 (2.21) | 132.6 ± 6.6 (2.25) |
| 1200 | limonene 1 | 91.1 ± 10.1 (3.55) | 13.9 ± 0.6 (0.57) | 16.4 ± 0.3 (0.32) | 44.1 ± 3.0 (0.86) | 18.8 ± 0.9 (0.43) | 25.3 ± 0.7 (0.43) |
| 1210 | 1,8-cineole 2 | 42.8 ± 3.7 (1.67) | 29.4 ± 1.9 (1.20) | 30.6 ± 0.3 (0.60) | 58.4 ± 6.3 (1.14) | 40.5 ± 2.4 (0.93) | 16.3 ± 0.7 (0.28) |
| 1226 | 2-hexenal 6 | - | - | - | t | - | - |
| 1237 | β-cis-ocimene 1 | - | - | 25.7 ± 0.7 (0.51) | 17.0 ± 1.5 (0.33) | 6.9 ± 0.6 (0.16) | 58.9 ± 4.2 (1.00) |
| 1249 | γ-terpinene 1 | 149.0 ± 15.2 (5.81) | 437.8 ± 16.3 (17.83) | 336.7 ± 8.3 (6.64) | 1152.1 ± 50.7(22.42) | 404.4 ± 28.7 (9.24) | 1291.0 ± 18.4 (21.93) |
| 1254 | β-trans-ocimene 1 | - | 3.4 ± 0.1 (0.14) | 5.9 ± 0.2 (0.12) | 9.6 ± 3.2 (0.19) | 6.8 ± 1.0 (0.16) | 18.4 ± 5.7 (0.31) |
| 1258 | 3-octanone 6 | - | 4.9 ± 0.1 (1.20) | - | - | - | 11.3 ± 0.3 (0.19) |
| 1275 | p-cymene 1 | 908.5 ± 35.9 (35.4) | 580.2 ± 22.6 (23.63) | 660.1 ± 21.9 (13.02) | 543.6 ± 42.6 (10.58) | 358.0 ± 30.1 (8.18) | 526.6 ± 29.7 (8.94) |
| 1286 | terpinolene 1 | - | 6.0 ± 0.3 (0.25) | 10.6 ± 0.1 (0.21) | 10.7 ± 0.5 (0.21) | 11.5 ± 0.8 (0.26) | 25.0 ± 1.0 (0.42) |
| 1308 | o-cymene 1 | 4.3 ± 0.1 (0.17) | - | - | - | - | - |
| 1320 | (Z)-3-hexenyl acetate 8 | - | 3.0 ± 0.1 (0.12) | - | t | - | 8.6 ± 0.3 (0.15) |
| 1342 | 4-pentenyl butyrate 8 | - | 7.0 ± 0.3 (0.28) | - | - | - | - |
| 1343 | sulcatone 6 | - | - | - | - | - | 8.4 ± 0.1 (0.14) |
| 1386 | 3-hexen-1-ol 7 | - | 7.6 ± 0.4 (0.31) | - | 14.4 ± 1.3 (0.28) | t | - |
| 1392 | 3-octanol 7 | - | 6.4 ± 0.5 (0.26) | 6.0 ± 0.0 (0.12) | 10.2 ± 0.5 (0.20) | 11.2 ± 0.2 (0.26) | 10.1 ± 0.5 (0.17) |
| 1399 | nonanal 6 | - | - | - | - | - | t |
| 1406 | 1-hepten-3-ol 7 | - | - | - | t | - | - |
| 1407 | fenchone 2 | 3.4 ± 0.1 (0.13) | - | - | - | - | - |
| 1432 | photocitral B 2 | - | - | - | - | - | t |
| 1434 | β-thujone 2 | 5.3 ± 0.3 (0.21) | - | - | - | - | - |
| 1450 | 1-octen-3-ol 7 | - | 28.9 ± 1.9 (1.18) | 65.0 ± 1.2 (1.28) | 161.5 ± 22.0 (3.14) | - | 42.3 ± 1.5 (0.72) |
| 1449 | trans-linalool furanoxide 2 | 4.0 ± 0.2 (0.16) | - | - | - | - | |
| 1453 | α-thujone 2 | 3.7 ± 0.2 (0.14) | - | - | - | - | |
| 1468 | trans-sabinene hydrate 2 | 3.3 ± 0.0 (0.13) | 16.5 ± 1.0 (0.67) | 26.4 ± 0.2 (0.52) | 31.0 ± 3.7 (0.60) | - | 60.7 ± 2.5 (1.03) |
| 1475 | cis-linalool furanoxide 2 | 3.9 ± 0.2 (0.15) | - | - | - | t | - |
| 1477 | isoledene 4 | - | - | 6.9 ± 0.7 (0.14) | - | - | - |
| 1478 | nerol oxide 2 | - | - | - | - | - | t |
| 1493 | α-ylangene 5 | - | - | t | - | - | - |
| 1496 | epiphotocitral A 2 | - | - | - | - | - | t |
| 1503 | α-copaene 5 | 3.2 ± 0.1 (0.13) | - | 10.9 ± 0.6 (0.21) | t | t | - |
| 1507 | decanal 6 | - | - | - | - | - | t |
| 1519 | 1-nonen-3-ol 7 | - | - | t | 5.6 ± 0.4 (0.11) | - | - |
| 1531 | β-bourbonene 4 | - | - | 10.1 ± 0.3 (0.20) | 6.3 ± 0.9 (0.12) | - | 19.2 ± 1.4 (0.33) |
| 1533 | camphor 2 | 8.2 ± 0.7 (0.32) | 13.3 ± 0.7 (0.54) | 5.9 ± 0.2 (0.12) | - | - | - |
| 1542 | α-gurjunene 4 | - | - | t | - | - | - |
| 1548 | β-linalool 2 | 187.5 ± 13.9 (7.31) | 88.0 ± 7.0 (3.58) | 35.2 ± 1.0 (0.69) | 103.5 ± 10.5 (2.01) | 138.7 ± 10.0 (3.17) | 11.2 ± 0.8 (0.19) |
| 1554 | cis-sabinene hydrate 2 | - | 6.9 ± 0.5 (0.28) | 13.6 ± 0.2 (0.27) | 9.5 ± 0.7 (0.19) | 14.8 ± 1.2 (0.34) | - |
| 1555 | isoneral 2 | - | - | - | - | - | 11.6 ± 1.6 (0.20) |
| 1570 | trans-p-menth-2-en-1-ol 2 | - | 4.4 ± 0.2 (0.18) | t | 6.0 ± 0.2 (0.12) | 5.3 ± 0.5 (0.12) | 6.3 ± 0.3 (0.11) |
| 1582 | p-menth-3-en-1-ol 2 | 3.1 ± 0.1 (0.12) | - | - | - | - | - |
| 1582 | isogeranial 2 | - | - | - | - | - | 11.2 ± 0.5 (0.19) |
| 1614 | β-caryophyllene 4 | 42.3 ± 3.6 (1.65) | 79.3 ± 4.9 (3.23) | 728.6 ± 25.6 (14.37) | 173.4 ± 17.4 (3.37) | 149.7 ± 10.3 (3.42) | 735.4 ± 26.9 (12.49) |
| 1586 | β-ylangene 4 | - | t | t | - | t | |
| 1593 | bornyl acetate 8 | 4.6 ± 0.2 (0.18) | 8.0 ± 0.3 (0.32) | 9.6 ± 0.1 (0.19) | - | - | t |
| 1602 | methylthymol 3 | - | 28.9 ± 1.7 (1.18) | 7.1 ± 0.1 (0.14) | t | - | 69.1 ± 3.2 (1.17) |
| 1606 | β-copaene 4 | - | - | t | t | t | t |
| 1618 | β-panasinsene 4 | - | - | 7.8 ± 2.3 (0.15) | - | - | - |
| 1624 | aromandendrene 4 | - | - | 48.5 ± 0.3 (0.96) | - | 11.9 ± 1.0 (0.27) | - |
| 1626 | trans-dihydrocarvone 2 | - | 4.1 ± 0.1 (0.17) | - | - | 8.6 ± 0.4 (0.20) | - |
| 1632 | selina-5,11-diene 4 | - | - | t | - | - | - |
| 1636 | cis-p-menth-2-en-1-ol 2 | - | - | - | t | - | t |
| 1637 | p-menth-8-en-1-ol 2 | 3.8 ± 0.1 (0.14) | - | - | - | - | - |
| 1646 | cis-dihydrocarvone 2 | - | - | - | - | 5.7 ± 0.1 (0.13) | - |
| 1653 | 1-decen-3-ol 7 | - | - | - | - | - | t |
| 1663 | allo-aromadendrene 4 | - | - | 9.5 ± 0.3 (0.19) | - | - | 11.3 ± 0.7 (0.19) |
| 1671 | trans-β-farnesene 4 | - | - | - | - | - | t |
| 1674 | cadina-1(6),4-diene 4 | - | - | t | - | - | - |
| 1678 | isoborneol 2 | 3.6 ± 0.1 (0.14) | - | - | - | - | - |
| 1680 | lavandulol 2 | - | 4.4 ± 0.1 (0.18) | - | - | - | - |
| 1681 | δ-terpineol 2 | - | - | - | t | - | - |
| 1687 | α-humulene 4 | 4.6 ± 0.2 (0.18) | 4.4 ± 0.2 (0.18) | 10.9 ± 0.2 (0.22) | 6.7 ± 0.6 (0.13) | 6.3 ± 0.4 (0.14) | 20.2 ± 2.0 (0.34) |
| 1696 | β-citral 2 | - | 3.5 ± 0.1 (0.32) | - | - | - | 111.6 ± 7.9 (1.90) |
| 1703 | γ-muurolene 4 | - | - | 18.3 ± 0.8 (0.36) | - | - | - |
| 1703 | α-terpineol 2 | 8.5 ± 0.5 (0.33) | 7.9 ± 0.3 (0.32) | 13.8 ± 0.4 (0.27) | 14.1 ± 1.7 (0.27) | 43.7 ± 2.7 (1.00) | 11.1 ± 0.6 (0.19) |
| 1708 | γ-terpineol 2 | 3.2 ± 0.1 (0.13) | - | - | - | - | - |
| 1712 | borneol 2 | 28.0 ± 2.7 (1.09) | 28.9 ± 2.0 (1.18) | 195.9 ± 2.9 (3.86) | 56.7 ± 8.3 (1.10) | 31.1 ± 1.9 (0.71) | 12.3 ± 0.4 (0.21) |
| 1727 | germacrene D 4 | - | 6.1 ± 0.5 (0.17) | - | 35.5 ± 2.9 (0.69) | t | 178.8 ± 17.5 (3.04) |
| 1736 | β-bisabolene 4 | - | - | 93.9 ± 2.7 (1.85) | 38.5 ± 2.8 (0.75) | t | 139.6 ± 15.0 (2.37) |
| 1742 | α-selinene 4 | - | - | t | - | - | - |
| 1746 | α-citral 2 | - | - | - | - | - | 170.1 ± 16.7 (2.89) |
| 1752 | bicyclogermacrene 4 | - | - | - | 20.9 ± 1.8 (0.41) | 15.6 ± 1.7 (0.36) | 34.4 ± 6.6 (0.58) |
| 1754 | carvone 2 | - | - | - | 11.3 ± 0.8 (0.22) | 10.2 ± 0.6 (0.23) | - |
| 1755 | nerol acetate 8 | - | - | - | - | - | 92.2 ± 7.9 (1.57) |
| 1760 | carvone 2 | - | - | 7.5 ± 0.1 (0.15) | - | - | - |
| 1761 | 1-decanol 7 | - | 3.8 ± 0.2 (0.15) | - | - | - | - |
| 1770 | δ-cadinene 4 | - | 3.8 ± 0.1 (0.15) | 25.5 ± 0.2 (0.50) | 6.5 ± 0.2 (0.13) | 6.0 ± 0.1 (0.14) | 22.9 ± 1.8 (0.39) |
| 1775 | γ-cadinene 4 | - | - | 12.0 ± 0.6 (0.24) | t | t | t |
| 1782 | trans-α-bisabolene 4 | - | - | t | t | - | - |
| 1782 | β-sesquiphellandrene 4 | - | - | - | - | - | 7.8 ± 0.3 (0.13) |
| 1797 | cubenene 4 | - | - | t | - | - | - |
| 1801 | p-cumic aldehyde 7 | - | - | t | - | - | - |
| 1805 | cis-geraniol 2 | - | - | - | - | - | 73.2 ± 6.8 (1.24) |
| 1808 | α-cadinene 4 | - | - | t | - | - | - |
| 1823 | geranyl propionate 8 | - | 5.9 ± 0.1 (0.24) | - | - | - | - |
| 1851 | trans-geraniol 2 | - | 5.2 ± 0.2 (0.21) | 6.2 ± 0.2 (0.12) | - | - | 492.5 ± 15.9 (8.37) |
| 1856 | p-cymen-8-ol 7 | - | 3.4 ± 0.1 (0.14) | 7.9 ± 0.5 (0.16) | t | t | - |
| 1861 | thymyl acetate 8 | - | 3.7 ± 0.1 (0.15) | 7.0 ± 0.4 (0.14) | - | - | - |
| 1885 | carvacryl acetate 8 | - | - | 10.1 ± 0.2 (0.20) | 49.5 ± 4.6 (0.96) | t | t |
| 1899 | nerolidol 5 | - | 3.6 ± 0.2 (0.15) | - | - | - | 6.6 ± 0.3 (0.11) |
| 1890 | epi-cubebol 5 | - | - | t | t | - | - |
| 1934 | α-calacorene 4 | - | - | t | - | - | - |
| 1950 | cubebol 5 | - | - | - | - | - | t |
| 2010 | caryophyllene oxide 5 | 9.2 ± 0.6 (0.36) | 12.8 ± 0.6 (0.52) | 23.0 ± 0.1 (0.45) | 11.2 ± 0.5 (0.22) | 7.9 ± 0.5 (0.18) | 51.7 ± 5.3 (0.88) |
| 2044 | trans-nerolidol 5 | - | - | - | - | - | 109.6 ± 7.8 (1.86) |
| 2065 | germacrene D-4-ol 5 | 3.2 ± 0.3 (0.12) | 3.6 ± 0.1 (0.15) | - | - | - | 30.8 ± 2.6 (0.52) |
| 2082 | epi-cubenol 5 | - | - | - | - | - | t |
| 2093 | globulol 5 | - | - | t | - | t | - |
| 2101 | viridiflorol 5 | - | - | t | - | - | - |
| 2106 | p-cymen-7-ol 3 | - | - | t | - | - | - |
| 2118 | spathulenol 5 | - | - | 17.0 ± 0.5 (0.34) | 9.1 ± 0.6 (0.18) | 9.3 ± 0.1 (0.21) | 9.9 ± 0.3 (0.17) |
| 2143 | thymol 3 | 684.2 ± 36.9 (26.66) | 677.6 ± 36.3 (27.59) | 867.3 ± 18.1 (17.10) | 72.6 ± 5.9 (1.41) | 43.9 ± 2.7 (1.00) | 220.3 ± 13.7 (3.74) |
| 2149 | tau-muurolol 5 | - | - | - | - | - | 10.8 ± 0.8 (0.18) |
| 2163 | δ-cadinol 5 | - | - | - | - | - | t |
| 2183 | carvacrol 3 | 78.9 ± 6.2 (3.07) | 34.7 ± 2.2 (1.41) | 1337.3 ± 8.4 (26.37) | 1826.0 ± 69.1 (35.54) | 2334.4 ± 46.1 (53.34) | 453.1 ± 22.3 (7.70) |
| 2219 | α-cadinol 5 | - | - | - | - | - | 21.6 ± 1.4 (0.37) |
| 2311 | caryophylladienol I 5 | - | - | - | - | - | t |
| 2316 | caryophylladienol II 5 | - | - | - | - | - | t |
| Total identified | 2536.9 (99.70) | 2388.7 (99.14) | 5041.9 (99.91) | 5090.2 (99.59) | 4270 (99.6) | 5849.1 (99.77) | |
| monoterpene hydrocarbons 1 | 1391.0 ± 74.4 | 1229.6 ± 49.3 | 1280.5 ± 35.1 | 2302.3 ± 144 | 1335.2 ± 105.9 | 2446.4 ± 90.2 | |
| oxygenated monoterpenes 2 | 312.4 ± 22.9 | 216.7 ± 14.2 | 339.7 ± 6.2 | 308.8 ± 28.5 | 302.5 ± 20.2 | 1008.4 ± 56.5 | |
| aromatic monoterpenoids 3 | 766.4 ± 43.1 | 749.7 ± 40.2 | 2233.8 ± 27 | 1906.1 ± 75 | 2382.8 ± 48.8 | 742.6 ± 39.3 | |
| sesquiterpene hydrocarbons 4 | 50.1 ± 4.0 | 93.7 ± 5.7 | 1029.8 ± 37.1 | 308.8 ± 28.5 | 209.6 ± 14.8 | 1188.5 ± 72.9 | |
| oxygenated sesquiterpenes 5 | 12.4 ± 0.9 | 20.1 ± 0.9 | 51.3 ± 0.9 | 23.7 ± 1.2 | 21.4 ± 0.8 | 263.8 ± 19.7 | |
| aliphatic aldehydes & ketones 6 | - | 4.9 ± 0.1 | 4.4 ± 0.1 | 4.5 ± 0.1 | - | 30.7 ± 0.7 | |
| aliphatic alcohols 7 | - | 46.7 ± 3.1 | 75.9 ± 1.4 | 191.8 ± 24.1 | 14.7 ± 0.4 | 57.3 ± 2.1 | |
| esters 8 | 4.6 ± 0.2 | 27.5 ± 0.9 | 26.6 ± 0.8 | 53.6 ± 4.8 | 3.8 ± 0.9 | 111.5 ± 8.6 | |
| Thymol * | 836.1 ± 44.4 | 828.0 ± 44.0 | 1056.1 ± 21.8 | 100.0 ± 7.1 | 65.36 ± 3.3 | 278.41 ± 16.5 | |
| Essential Oils | MIC μL/mL | MBC 95% μL/mL | MBC 99.5% μL/mL | ||||
|---|---|---|---|---|---|---|---|
| Faecal E. coli | ATCC 25922 | Faecal E. coli | ATCC 25922 | Faecal E. coli | ATCC 25922 | ||
| Commercial (SEU) | T. serpyllum | 0.209 ± 0.0464 | 0.488 ± 0.0000 | 0.209 ± 0.0464 | 0.488 ± 0.0000 | 0.209 ± 0.0464 | 0.488 ± 0.0000 |
| T. vulgaris | 0.148 ± 0.0200 | 0.079 ± 0.0202 | 0.148 ± 0.0200 | 0.079 ± 0.0202 | 0.148 ± 0.0200 | 0.079 ± 0.0202 | |
| S. montana | 0.073 ± 0.0137 | 0.006 ± 0.0031 | 0.076 ± 0.0144 | 0.006 ± 0.0031 | 0.076 ± 0.0144 | 0.006 ± 0.0031 | |
| Latvian (NEU) | T. serpyllum | 1.004 ± 0.1149 | 1.465 ± 0.2819 | 1.221 ± 0.1134 | 1.465 ± 0.2819 | 1.221 ± 0.1134 | 1.465 ± 0.2819 |
| T. vulgaris | 0.590 ± 0.0713 | 0.977 ± 0.0000 | 0.644 ± 0.0763 | 0.977 ± 0.0000 | 0.658 ± 0.0733 | 0.977 ± 0.0000 | |
| S. montana | 0.278 ± 0.0434 | 0.366 ± 0.1221 | 0.278 ± 0.0434 | 0.367 ± 0.1221 | 0.278 ± 0.0434 | 0.366 ± 0.1221 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gāliņa, D.; Radenkovs, V.; Kviesis, J.; Valdovska, A. Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs. Antibiotics 2022, 11, 461. https://doi.org/10.3390/antibiotics11040461
Gāliņa D, Radenkovs V, Kviesis J, Valdovska A. Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs. Antibiotics. 2022; 11(4):461. https://doi.org/10.3390/antibiotics11040461
Chicago/Turabian StyleGāliņa, Daiga, Vitalijs Radenkovs, Jorens Kviesis, and Anda Valdovska. 2022. "Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs" Antibiotics 11, no. 4: 461. https://doi.org/10.3390/antibiotics11040461
APA StyleGāliņa, D., Radenkovs, V., Kviesis, J., & Valdovska, A. (2022). Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs. Antibiotics, 11(4), 461. https://doi.org/10.3390/antibiotics11040461