
Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages

 ,
,  , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
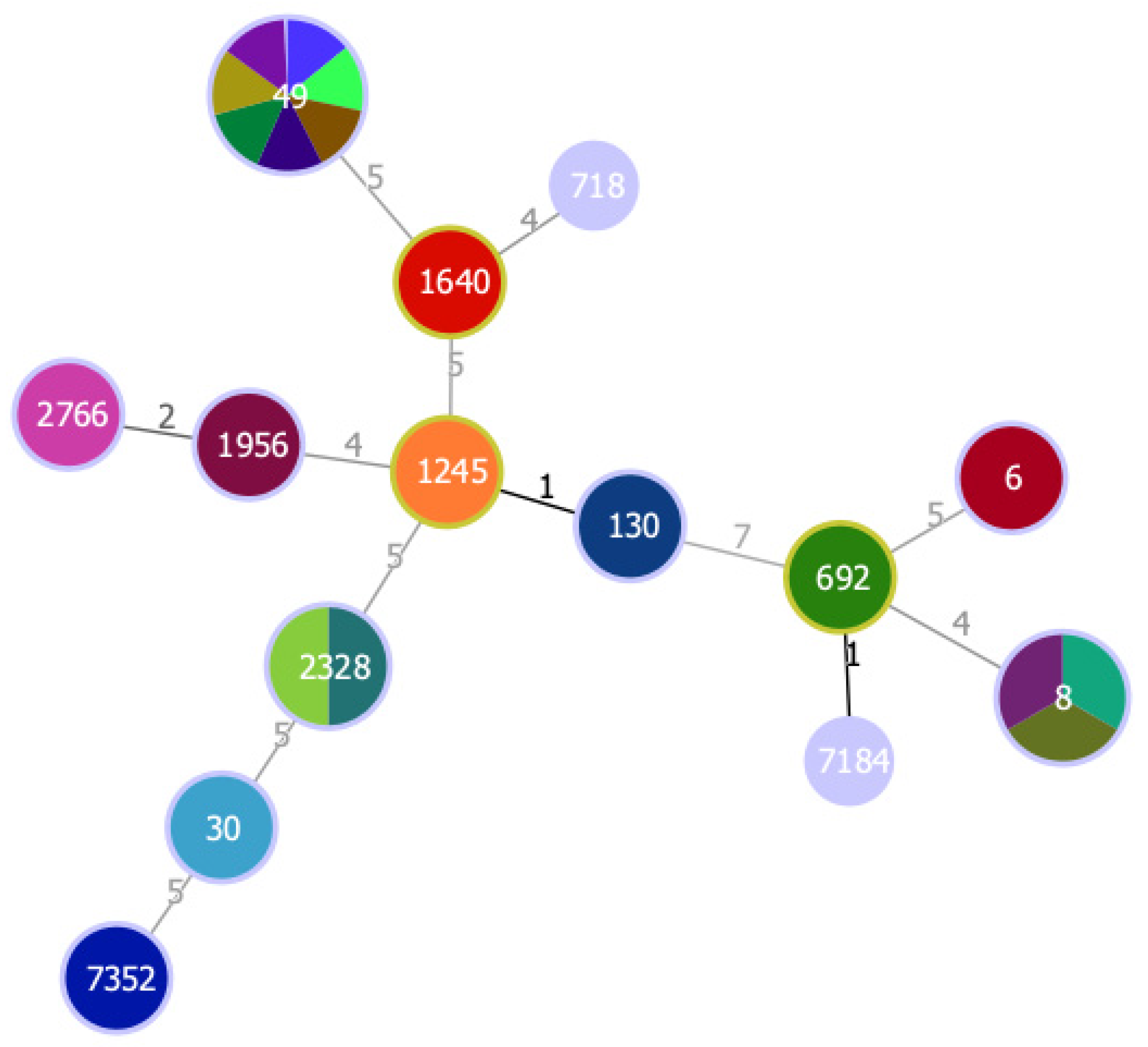
2. Results
2.1. Frequency and Distribution of Staphylococci in Night Prey
2.2. Characterization of CoPS Isolates
2.3. Characterization of CoNS Isolates
3. Discussion
4. Materials and Methods
4.1. Samples and Bacterial Isolates
4.2. Antimicrobial Susceptibility
4.3. Detection of Antimicrobial Resistance and Virulence Genes
4.4. Molecular Typing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- França, A.; Gaio, V.; Lopes, N.; Melo, L.D.R. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Goncheva, M.I.; Flannagan, R.S.; Deecker, S.R.; Guariglia-Oropeza, V.; Ensminger, A.W.; Heinrichs, D.E. Coagulase-negative staphylococci release a purine analog that inhibits Staphylococcus aureus virulence. Nat. Commun. 2021, 12, 1887. [Google Scholar] [CrossRef] [PubMed]
- Natsis, N.E.; Cohen, P.R. Coagulase-negative staphylococcus skin and soft tissue infections. Am. J. Clin. Dermatol. 2018, 19, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Both, A.; Weißelberg, S.; Heilmann, C.; Rohde, H. Emergence of coagulase-negative staphylococci. Expert Rev. Anti. Infect. Ther. 2020, 18, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.H.; Alsallaq, R.; Parsons, J.B.; Ferrolino, J.; Hayden, R.T.; Rubnitz, J.E.; Rafiqullah, I.M.; Robinson, D.A.; Margolis, E.B.; Rosch, J.W. Vancomycin heteroresistance and clinical outcomes in bloodstream infections caused by coagulase-negative staphylococci. Antimicrob. Agents Chemother. 2020, 64, e00944-20. [Google Scholar] [CrossRef] [PubMed]
- Medis, L.S.D.; Dissanayake, D.; Kottahachchi, J.; Weerasekera, M.M.; Namali, D. Detection of biofilm forming ability of coagulase negative Staphylococcus isolated from patients with central venous catheter infections and catheter colonization at a tertiary care hospital in Colombo. In Proceedings of the Research Conference in Health Sciences 2021, Nugegoda, Sri Lanka, 31 March 2021. [Google Scholar]
- Algammal, A.M.; Hetta, H.F.; Elkelish, A.; Alkhalifah, D.H.H.; Hozzein, W.N.; Batiha, G.E.-S.; El Nahhas, N.; Mabrok, M.A. Methicillin-Resistant Staphylococcus aureus (MRSA): One Health Perspective Approach to the Bacterium Epidemiology, Virulence Factors, Antibiotic-Resistance, and Zoonotic Impact. Infect. Drug Resist. 2020, 13, 3255–3265. [Google Scholar] [CrossRef] [PubMed]
- Carretto, E.; Visiello, R.; Nardini, P. Chapter 17—Methicillin Resistance in Staphylococcus aureus. In Pet-to-Man Travelling Staphylococci; Savini, V., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 225–235. ISBN 978-0-12-813547-1. [Google Scholar]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-Encoded Transferable mecB-Mediated Methicillin Resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Urushibara, N.; Aung, M.S.; Kawaguchiya, M.; Kobayashi, N. Novel staphylococcal cassette chromosome mec (SCCmec) type XIV (5A) and a truncated SCCmec element in SCC composite islands carrying speG in ST5 MRSA in Japan. J. Antimicrob. Chemother. 2020, 75, 46–50. [Google Scholar] [CrossRef]
- Silva, V.; Capelo, J.L.; Igrejas, G.; Poeta, P. Molecular Epidemiology of Staphylococcus aureus Lineages in Wild Animals in Europe: A Review. Antibiotics 2020, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Porrero, M.C.; Valverde, A.; Fernández-Llario, P.; Díez-Guerrier, A.; Mateos, A.; Lavín, S.; Cantón, R.; Fernández-Garayzabal, J.-F.; Domínguez, L. Staphylococcus aureus carrying mecC gene in animals and urban wastewater, Spain. Emerg. Infect. Dis. 2014, 20, 899–901. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Ferreira, E.; Manageiro, V.; Reis, L.; Tejedor-Junco, M.T.; Sampaio, A.; Capelo, J.L.; Caniça, M.; Igrejas, G.; Poeta, P. Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains. Antibiotics 2021, 10, 1416. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Soufi, L.; Campanile, F.; Dhaouadi, F.; Sociale, M.; Lazzaro, L.; Cherif, A.; Stefani, S.; Elandoulsi, R.B. Prevalence of meticillin-resistant and -susceptible coagulase-negative staphylococci with the first detection of the mecC gene among cows, humans and manure in Tunisia. Int. J. Antimicrob. Agents 2020, 55, 105826. [Google Scholar] [CrossRef] [PubMed]
- Schwendener, S.; Cotting, K.; Perreten, V. Novel methicillin resistance gene mecD in clinical Macrococcus caseolyticus strains from bovine and canine sources. Sci. Rep. 2017, 7, 43797. [Google Scholar] [CrossRef] [PubMed]
- Úngari, L.P.; Santos, A.L.Q.; Paiva, G.C.M.; Mota, K.C.P.; de Almeida Borges, L.F.; Cury, M.C. Concomitant infection of Haemogregarina sp. and Staphylococcus aureus in free-living yellow-spotted river turtle (Podocnemis unifilis): Case report. Arch. Vet. Sci. 2018, 23, 50–55. [Google Scholar] [CrossRef]
- Fri, J.; Njom, H.A.; Ateba, C.N.; Ndip, R.N. Antibiotic resistance and virulence gene characteristics of methicillin-resistant Staphylococcus aureus (MRSA) isolated from healthy Edible Marine Fish. Int. J. Microbiol. 2020, 2020, 9803903. [Google Scholar] [CrossRef]
- Kukułowicz, A.; Steinka, I.; Siwek, A. Presence of Antibiotic-Resistant Staphylococcus aureus in Fish and Seafood Originating from Points of Sale in the Tri-City Area (Poland). J. Food Prot. 2021, 84, 1911–1914. [Google Scholar] [CrossRef]
- Sousa, M.; Silva, N.; Igrejas, G.; Silva, F.; Sargo, R.; Alegria, N.; Benito, D.; Gómez, P.; Lozano, C.; Gómez-Sanz, E.; et al. Antimicrobial resistance determinants in Staphylococcus spp. recovered from birds of prey in Portugal. Vet. Microbiol. 2014, 171, 436–440. [Google Scholar] [CrossRef]
- Silva, V.; Gabriel, S.I.; Borrego, S.B.; Tejedor-Junco, M.T.; Manageiro, V.; Ferreira, E.; Reis, L.; Caniça, M.; Capelo, J.L.; Igrejas, G.; et al. Antimicrobial Resistance and Genetic Lineages of Staphylococcus aureus from Wild Rodents: First Report of mecC-Positive Methicillin-Resistant S. aureus (MRSA) in Portugal. Animals 2021, 11, 1537. [Google Scholar] [CrossRef]
- Seinige, D.; Von Altrock, A.; Kehrenberg, C. Genetic diversity and antibiotic susceptibility of Staphylococcus aureus isolates from wild boars. Comp. Immunol. Microbiol. Infect. Dis. 2017, 54, 7–12. [Google Scholar] [CrossRef]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as Sentinels of Antimicrobial Resistance in Germany? Front. Vet. Sci. 2021, 7, 1251. [Google Scholar] [CrossRef]
- Mencía-Gutiérrez, A.; Martín-Maldonado, B.; Pastor-Tiburón, N.; Moraleda, V.; González, F.; García-Peña, F.J.; Pérez-Cobo, I.; Revuelta, L.; Marín, M. Prevalence and antimicrobial resistance of Campylobacter from wild birds of prey in Spain. Comp. Immunol. Microbiol. Infect. Dis. 2021, 79, 101712. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, Z.-B.; Zeng, Z.-L.; Yang, X.-W.; Huang, Y.; Liu, J.-H. The role of wildlife (wild birds) in the global transmission of antimicrobial resistance genes. Zool. Res. 2017, 38, 55–80. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; Ramiro, Y.; de la Puente, J.; Fernández-Fernández, R.; Quevedo, M.Á.; Blanco, J.M.; Báguena, G.; et al. Frequency and Characterization of Antimicrobial Resistance and Virulence Genes of Coagulase-Negative Staphylococci from Wild Birds in Spain. Detection of tst-Carrying S. sciuri Isolates. Microorganisms 2020, 8, 1317. [Google Scholar] [CrossRef] [PubMed]
- De Mesquita Souza Saraiva, M.; de Leon, C.M.C.G.; da Silva, N.M.V.; Raso, T.F.; Serafini, P.P.; Givisiez, P.E.N.; Gebreyes, W.A.; de Oliveira, C.J.B. Staphylococcus sciuri as a Reservoir of mecA to Staphylococcus aureus in Non-Migratory Seabirds from a Remote Oceanic Island. Microb. Drug Resist. 2020, 27, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Gambino, D.; Vicari, D.; Vitale, M.; Schirò, G.; Mira, F.; Giglia, M.L.; Riccardi, A.; Gentile, A.; Giardina, S.; Carrozzo, A.; et al. Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy. Microorganisms 2021, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Peri, A. Censusing a tawny owl (Strix aluco) population living at high density merging two consolidated techniques. Écoscience 2018, 25, 249–257. [Google Scholar] [CrossRef]
- Fröhlich, A.; Ciach, M. Noise pollution and decreased size of wooded areas reduces the probability of occurrence of Tawny Owl Strix aluco. Ibis (Lond. 1859) 2018, 160, 634–646. [Google Scholar] [CrossRef]
- Lourenço, R.; Roque, I.; Tomé, R. Relatório do Programa NOCTUA Portugal (2009/10–2019/20); Sociedade Portuguesa para o Estudo das Aves: Lisbon, Portugal, 2021. [Google Scholar]
- Sun, C.-H.; Liu, H.-Y.; Min, X.; Lu, C.-H. Mitogenome of the little owl Athene noctua and phylogenetic analysis of Strigidae. Int. J. Biol. Macromol. 2020, 151, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Lee, J.-H.; Sung, H.-C. A case study of male tawny owl (Strix aluco) vocalizations in South Korea: Call feature, individuality, and the potential use for census. Anim. Cells Syst. (Seoul) 2019, 23, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.P.; Clément, L.; Uva, V.; Goudet, J.; Roulin, A. The Rocky Mountains as a dispersal barrier between barn owl (Tyto alba) populations in North America. J. Biogeogr. 2018, 45, 1288–1300. [Google Scholar] [CrossRef]
- Moysi, M.; Christou, M.; Goutner, V.; Kassinis, N.; Iezekiel, S. Spatial and temporal patterns in the diet of barn owl (Tyto alba) in Cyprus. J. Biol. Res. 2018, 25, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, A.; Baldomà, L.; Molina-López, R.A.; Martin, M.; Darwich, L. Microbiological diagnosis and antimicrobial sensitivity profiles in diseased free-living raptors. Avian Pathol. 2017, 46, 442–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, M.; Silva, N.; Igrejas, G.; Sargo, R.; Benito, D.; Gómez, P.; Lozano, C.; Manageiro, V.; Torres, C.; Caniça, M.; et al. Genetic Diversity and Antibiotic Resistance Among Coagulase-Negative Staphylococci Recovered from Birds of Prey in Portugal. Microb. Drug Resist. 2016, 22, 727–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dipineto, L.; Bossa, L.M.D.L.; Pace, A.; Russo, T.P.; Gargiulo, A.; Ciccarelli, F.; Raia, P.; Caputo, V.; Fioretti, A. Microbiological survey of birds of prey pellets. Comp. Immunol. Microbiol. Infect. Dis. 2015, 41, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; de la Puente, J.; Fernández-Fernández, R.; Ramiro, Y.; Quevedo, M.A.; Blanco, J.M.; Zarazaga, M.; et al. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu(A) in Magpies and Cinereous Vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef]
- Silva, V.; Pereira, J.E.; Maltez, L.; Ferreira, E.; Manageiro, V.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Diversity of methicillin-resistant staphylococci among wild Lepus granatensis: First detection of mecA-MRSA in hares. FEMS Microbiol. Ecol. 2019, 96, fiz204. [Google Scholar] [CrossRef]
- MacFadyen, A.C.; Harrison, E.M.; Ellington, M.J.; Parkhill, J.; Holmes, M.A.; Paterson, G.K. A highly conserved mecC-encoding SCCmec type XI in a bovine isolate of methicillin-resistant Staphylococcus xylosus. J. Antimicrob. Chemother. 2018, 73, 3516–3518. [Google Scholar] [CrossRef]
- Espinosa-Gongora, C.; Harrison, E.M.; Moodley, A.; Guardabassi, L.; Holmes, M.A. MRSA carrying mecC in captive mara. J. Antimicrob. Chemother. 2015, 70, 1622–1624. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Gavier-Widen, D.; Mattsson, R.; Rangstrup-Christensen, L.; Lazaris, A.; Coleman, D.C.; Shore, A.C.; Ehricht, R. Detection of mecC-Positive Staphylococcus aureus (CC130-MRSA-XI) in Diseased European Hedgehogs (Erinaceus europaeus) in Sweden. PLoS One 2013, 8, e66166. [Google Scholar] [CrossRef] [Green Version]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of staphylococcal cassette chromosome mec type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef] [Green Version]
- Haenni, M.; Châtre, P.; Dupieux, C.; Métayer, V.; Maillard, K.; Bes, M.; Madec, J.-Y.; Laurent, F. mecC-positive MRSA in horses. J. Antimicrob. Chemother. 2015, 70, 3401–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet. Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Feßler, A.T.; Thomas, P.; Mühldorfer, K.; Grobbel, M.; Brombach, J.; Eichhorn, I.; Monecke, S.; Ehricht, R.; Schwarz, S. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from zoo and wild animals. Vet. Microbiol. 2018, 218, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Giacinti, G.; Carfora, V.; Caprioli, A.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Amoruso, R.; Iurescia, M.; Stravino, F.; Dottarelli, S.; et al. Prevalence and characterization of methicillin-resistant Staphylococcus aureus carrying mecA or mecC and methicillin-susceptible Staphylococcus aureus in dairy sheep farms in central Italy. J. Dairy Sci. 2017, 100, 7857–7863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; Lozano, C.; Camacho, M.C.; Lima-Barbero, J.-F.; Hernández, J.-M.; Zarazaga, M.; Höfle, Ú.; Torres, C. Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 2015, 71, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in wild small mammals in Spain. J. Antimicrob. Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; Lozano, C.; González-Barrio, D.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. High prevalence of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in a semi-extensive red deer (Cervus elaphus hispanicus) farm in Southern Spain. Vet. Microbiol. 2015, 177, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Huber, C.; Stamm, I.; Ziebuhr, W.; Marincola, G.; Bischoff, M.; Strommenger, B.; Jaschkowitz, G.; Marciniak, T.; Cuny, C.; Witte, W.; et al. Silence as a way of niche adaptation: mecC-MRSA with variations in the accessory gene regulator (agr) functionality express kaleidoscopic phenotypes. Sci. Rep. 2020, 10, 14787. [Google Scholar] [CrossRef]
- Zehra, A.; Singh, R.; Kaur, S.; Gill, J.P.S. Molecular characterization of antibiotic-resistant Staphylococcus aureus from livestock (bovine and swine). Vet. world 2017, 10, 598–604. [Google Scholar] [CrossRef]
- Mrochen, D.M.; Schulz, D.; Fischer, S.; Jeske, K.; El Gohary, H.; Reil, D.; Imholt, C.; Trübe, P.; Suchomel, J.; Tricaud, E.; et al. Wild rodents and shrews are natural hosts of Staphylococcus aureus. Int. J. Med. Microbiol. 2018, 308, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Simpson, V.; Davison, N.; Hudson, L.; Enright, M.; Whatmore, A.M. Staphylococcus aureus ST49 infection in red squirrels. Vet. Rec. 2010, 167, 69. [Google Scholar] [CrossRef] [PubMed]
- Gryz, J.; Krauze-Gryz, D. Changes in the tawny owl Strix aluco diet along an urbanisation gradient. Biologia (Bratisl) 2019, 74, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Rohmer, C.; Wolz, C. The Role of hlb-Converting Bacteriophages in Staphylococcus aureus Host Adaption. Microb. Physiol. 2021, 31, 109–122. [Google Scholar] [CrossRef]
- Jung, P.; Abdelbary, M.M.H.; Kraushaar, B.; Fetsch, A.; Geisel, J.; Herrmann, M.; Witte, W.; Cuny, C.; Bischoff, M. Impact of bacteriophage Saint3 carriage on the immune evasion capacity and hemolytic potential of Staphylococcus aureus CC398. Vet. Microbiol. 2017, 200, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Hossain, M.J.; Liles, M.R.; Panizzi, P. Complete genome sequence of Staphylococcus aureus Tager 104, a sequence type 49 ancestor. Genome Announc. 2013, 1, e00706-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakiewicz, A.; Ziółkowska, G.; Zięba, P.; Gnat, S.; Wojtanowicz-Markiewicz, K.; Trościańczyk, A. Coagulase-positive Staphylococcus isolated from wildlife: Identification, molecular characterization and evaluation of resistance profiles with focus on a methicillin-resistant strain. Comp. Immunol. Microbiol. Infect. Dis. 2016, 44, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST8. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier-Lachance, J.; Arsenault, J.; Usongo, V.; Parent, É.; Labrie, J.; Jacques, M.; Malouin, F.; Archambault, M. Prevalence and characteristics of Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) isolated from chicken meat in the province of Quebec, Canada. PLoS One 2020, 15, e0227183. [Google Scholar] [CrossRef]
- Silva, V.; Monteiro, A.; Porto, M.; Sampaio, A.; Maltez, L.; Pereira, J.E.; Aonofriesei, F.; Capelo, J.L.; Igrejas, G.; Poeta, P. Molecular Diversity of Methicillin-Resistant and -Susceptible Staphylococcus aureus Detected in Animals: A Focus on Aquatic Animals. Diversity 2021, 13, 417. [Google Scholar] [CrossRef]
- Thapaliya, D.; Dalman, M.; Kadariya, J.; Little, K.; Mansell, V.; Taha, M.Y.; Grenier, D.; Smith, T.C. Characterization of Staphylococcus aureus in Goose Feces from State Parks in Northeast Ohio. Ecohealth 2017, 14, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Porrero, M.C.; Mentaberre, G.; Sánchez, S.; Fernández-Llario, P.; Casas-Díaz, E.; Mateos, A.; Vidal, D.; Lavín, S.; Fernández-Garayzábal, J.-F.; Domínguez, L. Carriage of Staphylococcus aureus by Free-Living Wild Animals in Spain. Appl. Environ. Microbiol. 2014, 80, 4865–4870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, M.; Silva, N.; Manageiro, V.; Ramos, S.; Coelho, A.; Gonçalves, D.; Caniça, M.; Torres, C.; Igrejas, G.; Poeta, P. First report on MRSA CC398 recovered from wild boars in the north of Portugal. Are we facing a problem? Sci. Total Environ. 2017, 596–597, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.; Dorothee, G.; Veronika, B.; Annette, B.; Julia, K.; André, G.; Stefan, W.; Birte, H.; Bauerfeind, S.S.; Paula, D.; et al. Molecular Epidemiology of Staphylococcus aureus in the General Population in Northeast Germany: Results of the Study of Health in Pomerania (SHIP-TREND-0). J. Clin. Microbiol. 2016, 54, 2774–2785. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus Isolates in European Wildlife. PLoS One 2016, 11, e0168433. [Google Scholar] [CrossRef] [Green Version]
- Mama, O.M.; Gómez, P.; Ruiz-Ripa, L.; Gómez-Sanz, E.; Zarazaga, M.; Torres, C. Antimicrobial Resistance, Virulence, and Genetic Lineages of Staphylococci from Horses Destined for Human Consumption: High Detection of S. aureus Isolates of Lineage ST1640 and Those Carrying the lukPQ Gene. Animals 2019, 9, 900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fountain, K.; Blackett, T.; Butler, H.; Carchedi, C.; Schilling, A.-K.; Meredith, A.; Gibbon, M.J.; Lloyd, D.H.; Loeffler, A.; Feil, E.J. Fatal exudative dermatitis in island populations of red squirrels (Sciurus vulgaris): Spillover of a virulent Staphylococcus aureus clone (ST49) from reservoir hosts. Microb. Genom. 2021, 7, 565. [Google Scholar] [CrossRef]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Loncaric, I.; Kübber-Heiss, A.; Posautz, A.; Ruppitsch, W.; Lepuschitz, S.; Schauer, B.; Feßler, A.T.; Krametter-Frötscher, R.; Harrison, E.M.; Holmes, M.A.; et al. Characterization of mecC gene-carrying coagulase-negative Staphylococcus spp. isolated from various animals. Vet. Microbiol. 2019, 230, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Tsubakishita, S.; Kuwahara-Arai, K.; Sasaki, T.; Hiramatsu, K. Origin and molecular evolution of the determinant of methicillin resistance in staphylococci. Antimicrob. Agents Chemother. 2010, 54, 4352–4359. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sybille, S.; Vincent, P. New MLSB Resistance Gene erm(43) in Staphylococcus lentus. Antimicrob. Agents Chemother. 2012, 56, 4746–4752. [Google Scholar] [CrossRef] [Green Version]
- Edslev, S.M.; Clausen, M.-L.; Agner, T.; Stegger, M.; Andersen, P.S. Genomic analysis reveals different mechanisms of fusidic acid resistance in Staphylococcus aureus from Danish atopic dermatitis patients. J. Antimicrob. Chemother. 2018, 73, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, C.A.R.; Pereira, I.A.; Rodrigues, D.P.; Siciliano, S. Staphylococcus spp. isolated from wild birds apprehended in the local illegal trade in Rio de Janeiro, Brazil, and relevance in public health. Lett. Appl. Microbiol. 2018, 67, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Regecová, I.; Pipová, M.; Jevinová, P.; Kmet’, V.; Výrostková, J.; Sopková, D. Antimicrobial Resistance of Coagulase-negative Species of Staphylococci Isolated from the Meat of Wild Pheasants (Phasianus Colchicus). Ital. J. Anim. Sci. 2014, 13, 3476. [Google Scholar] [CrossRef]
- EUCAST European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint tables for interpretation of MICs and Zone diameters Version 8.0; EUCAST European Committee on Antimicrobial Susceptibility Testing (EUCAST): Växjö, Sweden, 2018. [Google Scholar]
- CLSI Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Silva, V.; Vieira-Pinto, M.; Saraiva, C.; Manageiro, V.; Reis, L.; Ferreira, E.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Prevalence and Characteristics of Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) CC398 Isolated from Quails (Coturnix Coturnix Japonica) Slaughtered for Human Consumption. Animals 2021, 11, 2038. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, D.; Claus, H.H.H.H.; Witte, W.; Rothgänger, J.; Claus, H.H.H.H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [Green Version]
- Shopsin, B.; Gomez, M.; Montgomery, S.O.; Smith, D.H.; Waddington, M.; Dodge, D.E.; Bost, D.A.; Riehman, M.; Naidich, S.; Kreiswirth, B.N. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 1999, 37, 3556–3563. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Owl Species | Number of Samples | Number of CoPS | Number of CoNS |
|---|---|---|---|
| Tyto alba (Barn owl) | 41 | 7 | 9 |
| Bubo bubo (Eurasian eagle-owl) | 5 | 1 | 4 |
| Athene noctua (Little owl) | 25 | 8 | 8 |
| Strix aluco (Tawny owl) | 43 | 9 | 20 |
| Total | 114 | 25 | 41 |
| Isolate | Owl Species | Antimicrobial Resistance | Virulence Factors | Molecular Typing | ||||
|---|---|---|---|---|---|---|---|---|
| Phenotype | Genotype | IEC Type | Other Genes | ST (CC) | spa | agr | ||
| VS2960 | Athene noctua | PEN, FOX | mecC, blaZ-SCCmecXI | hla, hlb, etd2 | 1245 (130) | t843 | III | |
| VS2961 | Strix aluco | PEN, FD | hla, hlb | 49 (49) | t208 | II | ||
| VS2962 | Strix aluco | PEN | blaZ | hla, hlb | 49 (49) | t208 | II | |
| VS2963 | Strix aluco | Susceptible | hla, hlb | 49 (49) | t208 | II | ||
| VS2964 | Strix aluco | Susceptible | hla, hlb | 49 (49) | t9811 | II | ||
| VS2965 | Athene noctua | Susceptible | hla, hlb | 49 (49) | t20169 | II | ||
| VS2966 | Athene noctua | Susceptible | hla, hlb | 49 (49) | t208 | II | ||
| VS2967 | Athene noctua | Susceptible | E | hla | 49 (49) | t208 | II | |
| VS2968 | Strix aluco | PEN | blaZ | B | hla | 8 (8) | t121 | I |
| VS2969 | Athene noctua | PEN, CIP | blaZ | B | hla | 8 (8) | t121 | I |
| VS2970 | Athene noctua | PEN, FD | blaZ | B | hla | 8 (8) | t121 | I |
| VS2971 | Tyto alba | Susceptible | hla, hlb | 2328 (133) | t3750 | III | ||
| VS2972 | Tyto alba | PEN, FD | hla, hlb | 2328 (133) | t3750 | III | ||
| VS2973 | Tyto alba | PEN, TET, FD | tetK | hla, hlb | 2766 (121) | t12364 | IV | |
| VS2974 | Bubo bubo | ERY | ermA | B | hla | 718 | t11333 | II |
| VS2975 | Athene noctua | PEN | blaZ | hla, hlb | 30 (30) | t9413 | III | |
| VS2976 | Tyto alba | TET | tetK | hla | 692 | t1422 | I | |
| VS2977 | Tyto alba | Susceptible | hla, hlb | 1956 (121) | t20223 | IV | ||
| VS2978 | Strix aluco | PEN, CIP, CD, FD | blaZ, mphC | hla, hlb | 130 (130) | t843 | III | |
| VS2979 | Strix aluco | Susceptible | hla, hlb | 1640 | t9853 | IV | ||
| VS2980 | Athene noctua | Susceptible | hla, hlb | 6 (5) | t16615 | I | ||
| VS2981 | Tyto alba | Susceptible | hla, hlb | 7184 | t2247 | I | ||
| VS2982 | Strix aluco | Susceptible | E | hla | 7352 | t2143 | I | |
| Isolate | Staphylococci Species | Owl Species | Antimicrobial Resistance | |
|---|---|---|---|---|
| Phenotype | Genotype | |||
| VS2985 | S. epidermidis | Bubo bubo | PEN, FD | blaZ, fusB |
| VS2986 | S. epidermidis | Bubo bubo | PEN, ERY, FD | blaZ, msr(A/B), mphC, fusB |
| VS2987 | S. sciuri | Strix aluco | Susceptible | |
| VS2988 | S. sciuri | Tyto alba | Susceptible | |
| VS2989 | S. sciuri | Tyto alba | PEN | mecA |
| VS2990 | S. sciuri | Strix aluco | PEN | mecA |
| VS2991 | S. sciuri | Tyto alba | PEN, CD, TET, FD | mecA, mphC, tetK |
| VS2992 | S. sciuri | Tyto alba | PEN, CD, FD | |
| VS2993 | S. sciuri | Tyto alba | PEN, FOX, CD | mecA, mphC |
| VS2994 | S. sciuri | Strix aluco | PEN | mecA |
| VS2995 | S. sciuri | Strix aluco | PEN, CD, FD | |
| VS2996 | S. sciuri | Strix aluco | Susceptible | |
| VS2997 | S. sciuri | Bubo bubo | Susceptible | |
| VS2998 | S. sciuri | Strix aluco | Susceptible | |
| VS2999 | S. sciuri | Strix aluco | PEN, FD | |
| VS3000 | S. sciuri | Athene noctua | PEN, FOX, CD, FD | mecA, mphC |
| VS3001 | S. sciuri | Athene noctua | Susceptible | |
| VS3002 | S. sciuri | Strix aluco | PEN | |
| VS3003 | S. sciuri | Strix aluco | PEN, FD | mecA |
| VS3004 | S. sciuri | Athene noctua | PEN, FOX, CD, TET, FD | mecA |
| VS3005 | S. sciuri | Athene noctua | Susceptible | |
| VS3006 | S. sciuri | Tyto alba | Susceptible | |
| VS3007 | S. lentus | Strix aluco | CD | mphC |
| VS3008 | S. lentus | Strix aluco | PEN, CD, TET, FD | tetK |
| VS3009 | S. lentus | Strix aluco | PEN, CD, TET | tetK |
| VS3010 | S. lentus | Strix aluco | CD, TET | mphC |
| VS3011 | S. lentus | Strix aluco | Susceptible | |
| VS3012 | S. lentus | Strix aluco | Susceptible | |
| VS3013 | S. lentus | Strix aluco | TET | tetK |
| VS3014 | S. lentus | Tyto alba | Susceptible | |
| VS3015 | S. lentus | Strix aluco | PEN, CD, FD | mecA |
| VS3016 | S. lentus | Athene noctua | FD | |
| VS3017 | S. lentus | Athene noctua | mecA | |
| VS3018 | S. vitulinus | Tyto alba | Susceptible | mecA |
| VS3019 | S. vitulinus | Athene noctua | PEN, FD | mecA |
| VS3020 | S. haemolyticus | Athene noctua | Susceptible | |
| VS3021 | S. haemolyticus | Bubo bubo | PEN, FOX, CIP, ERY, CD, TET, SXT | mphC, msr(A/B), tetO |
| VS3022 | S. saprophyticus | Tyto alba | Susceptible | mecA |
| VS3023 | S. xylosus | Strix aluco | PEN, FOX, ERY, CD, TET, C, FD | mphC, tetM, tetL |
| VS3024 | S. xylosus | Strix aluco | ERY | |
| VS3025 | S. succinus | Strix aluco | PEN | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.; Lopes, A.F.; Soeiro, V.; Caniça, M.; Manageiro, V.; Pereira, J.E.; Maltez, L.; Capelo, J.L.; Igrejas, G.; Poeta, P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics 2022, 11, 240. https://doi.org/10.3390/antibiotics11020240
Silva V, Lopes AF, Soeiro V, Caniça M, Manageiro V, Pereira JE, Maltez L, Capelo JL, Igrejas G, Poeta P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics. 2022; 11(2):240. https://doi.org/10.3390/antibiotics11020240
Chicago/Turabian StyleSilva, Vanessa, Ana Filipa Lopes, Vanessa Soeiro, Manuela Caniça, Vera Manageiro, José Eduardo Pereira, Luís Maltez, José Luis Capelo, Gilberto Igrejas, and Patrícia Poeta. 2022. "Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages" Antibiotics 11, no. 2: 240. https://doi.org/10.3390/antibiotics11020240
APA StyleSilva, V., Lopes, A. F., Soeiro, V., Caniça, M., Manageiro, V., Pereira, J. E., Maltez, L., Capelo, J. L., Igrejas, G., & Poeta, P. (2022). Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics, 11(2), 240. https://doi.org/10.3390/antibiotics11020240