
Antimicrobial and Biocide Resistance among Feline and Canine Staphylococcus aureus and Staphylococcus pseudintermedius Isolates from Diagnostic Submissions

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Origin of the Isolates
2.2. Antimicrobial Susceptibility of Feline S. aureus Isolates
2.3. Antimicrobial Susceptibility of Canine S. aureus Isolates
2.4. Antimicrobial Susceptibility of Feline S. pseudintermedius Isolates
2.5. Antimicrobial Susceptibility of Canine S. pseudintermedius Isolates
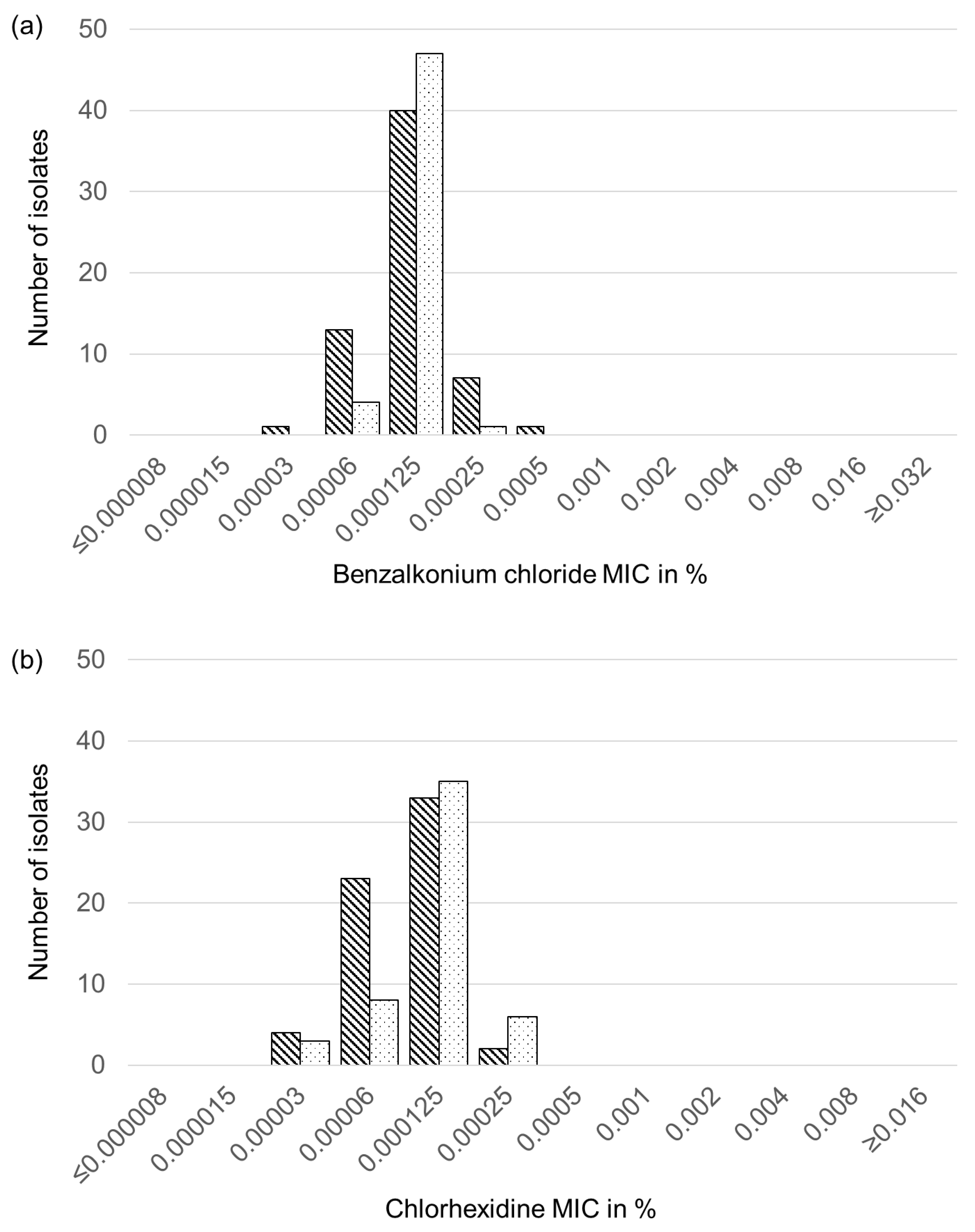
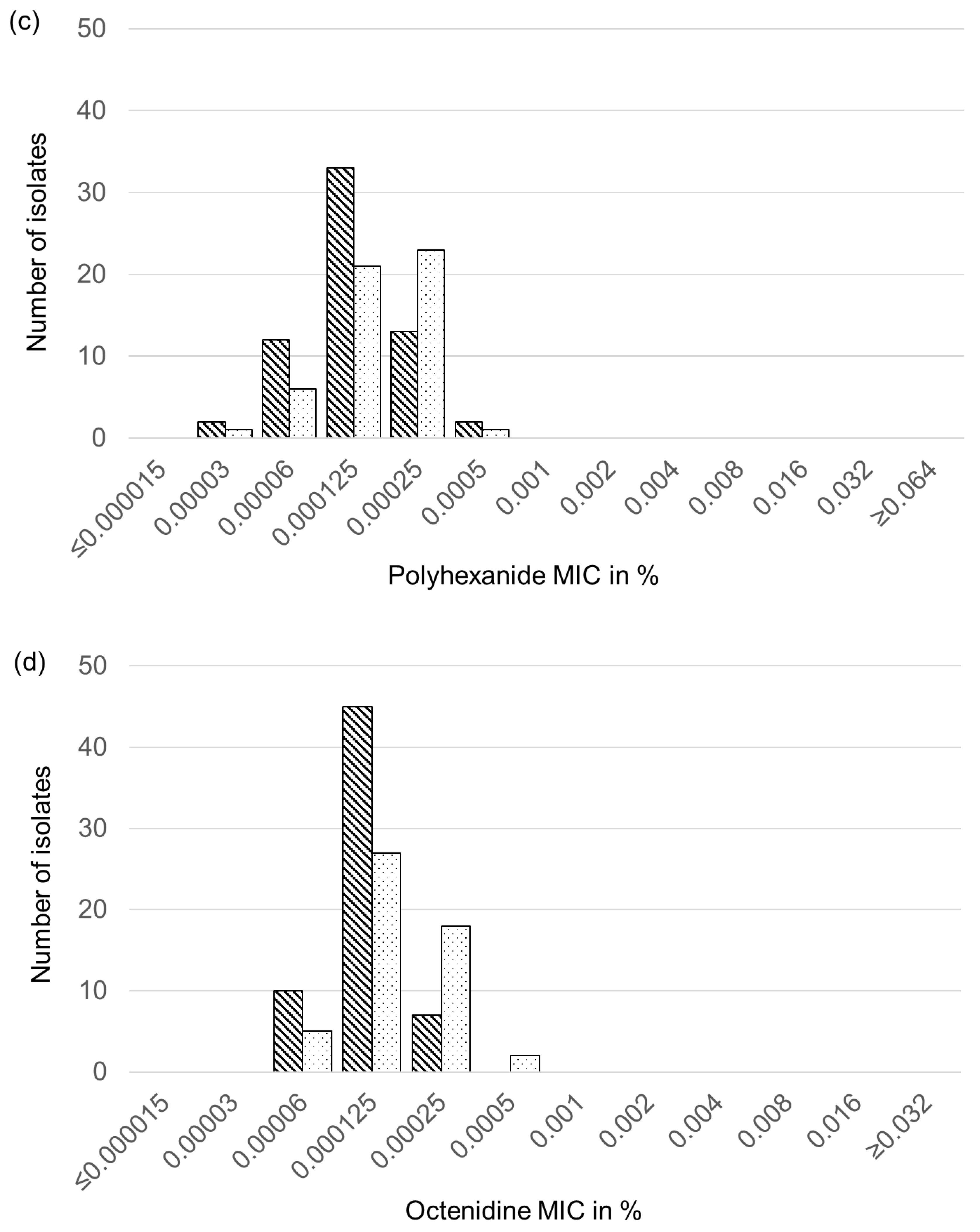
2.6. Biocide Susceptibility of Feline and Canine S. aureus and S. pseudintermedius Isolates
3. Discussion
4. Materials and Methods
4.1. Identification of S. aureus and S. pseudintermedius
4.2. Antimicrobial Susceptibility Testing and Detection of Resistance Genes
4.3. Calculation of the MAR Index
4.4. Biocide Susceptibility Testing
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haag, A.F.; Fitzgerald, J.R.; Penadés, J.R. Staphylococcus aureus in Animals. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.A.; Helbig, K.J. The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next? Vet. Sci. 2021, 8, 11. [Google Scholar] [CrossRef]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet animals as reservoirs of antimicrobial-resistant bacteria. J. Antimicrob. Chemother. 2004, 54, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, A.; Lloyd, D.H. Companion animals: A reservoir for methicillin-resistant Staphylococcus aureus in the community? Epidemiol. Infect. 2010, 138, 595–605. [Google Scholar] [CrossRef]
- Sing, A.; Tuschak, C.; Hörmansdorfer, S. Methicillin-resistant Staphylococcus aureus in a family and its pet cat. N. Engl. J. Med. 2008, 358, 1200–1201. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Hermes, J.; Cuny, C.; Wieler, L.H.; Vincze, S.; Abou Elnaga, Y.; Stamm, I.; Kopp, P.A.; Kohn, B.; Witte, W.; et al. Sharing more than friendship—nasal colonization with coagulase-positive staphylococci (CPS) and co-habitation aspects of dogs and their owners. PLoS ONE 2012, 7, e35197. [Google Scholar] [CrossRef]
- Blondeau, L.D.; Deutscher, M.; Rubin, J.E.; Deneer, H.; Kanthan, R.; Sanche, S.; Blondeau, J.M. Urinary tract infection in a human male patient with Staphylococcus pseudintermedius transmission from the family dog. J. Chemother. 2021, 8, 1–4. [Google Scholar] [CrossRef]
- Hackmann, C.; Gastmeier, P.; Schwarz, S.; Lübke-Becker, A.; Bischoff, P.; Leistner, R. Pet husbandry as a risk factor for colonization or infection with MDR organisms: A systematic meta-analysis. J. Antimicrob. Chemother. 2021, 76, 1392–1405. [Google Scholar] [CrossRef]
- Schwarz, S.; Feßler, A.T.; Loncaric, I.; Wu, C.; Kadlec, K.; Wang, Y.; Shen, J. Antimicrobial Resistance among Staphylococci of Animal Origin. Microbiol Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Cain, C.L. Antimicrobial resistance in staphylococci in small animals. Vet. Clin. N. Am. Small Anim. Pract. 2013, 43, 19–40. [Google Scholar] [CrossRef]
- Malik, S.; Peng, H.; Barton, M.D. Antibiotic resistance in staphylococci associated with cats and dogs. J. Appl. Microbiol. 2005, 99, 1283–1293. [Google Scholar] [CrossRef]
- Kadlec, K.; Schwarz, S. Antimicrobial resistance of Staphylococcus pseudintermedius. Vet. Dermatol. 2012, 23, 276-e55. [Google Scholar] [CrossRef]
- Weese, J.S.; van Duijkeren, E. Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary medicine. Vet. Microbiol. 2010, 140, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Tedin, K.; Lübke-Becker, A. Multidrug-resistant opportunistic pathogens challenging veterinary infection control. Vet Microbiol. 2017, 200, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ripa, L.; Simón, C.; Ceballos, S.; Ortega, C.; Zarazaga, M.; Torres, C.; Gómez-Sanz, E. S. pseudintermedius and S. aureus lineages with transmission ability circulate as causative agents of infections in pets for years. BMC Vet. Res. 2021, 17, 42. [Google Scholar] [CrossRef]
- Davis, J.A.; Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Brousse, J.H.; Gustafson, J.; Kucher, M. Carriage of methicillin-resistant staphylococci by healthy companion animals in the US. Lett. Appl. Microbiol. 2014, 59, 1–8. [Google Scholar] [CrossRef]
- Perreten, V.; Kadlec, K.; Schwarz, S.; Grönlund Andersson, U.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A.; et al. Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: An international multicentre study. J. Antimicrob. Chemother. 2010, 65, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, K.; Schwarz, S.; Perreten, V.; Grönlund Andersson, U.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A.; et al. Molecular analysis of methicillin-resistant Staphylococcus pseudintermedius of feline origin from different European countries and North America. J. Antimicrob. Chemother. 2010, 65, 1826–1828. [Google Scholar] [CrossRef]
- Ruzauskas, M.; Couto, N.; Pavilonis, A.; Klimiene, I.; Siugzdiniene, R.; Virgailis, M.; Vaskeviciute, L.; Anskiene, L.; Pomba, C. Characterization of Staphylococcus pseudintermedius isolated from diseased dogs in Lithuania. Pol. J. Vet. Sci. 2016, 19, 7–14. [Google Scholar] [CrossRef][Green Version]
- Loncaric, I.; Tichy, A.; Handler, S.; Szostak, M.P.; Tickert, M.; Diab-Elschahawi, M.; Spergser, J.; Künzel, F. Prevalence of Methicillin-Resistant Staphylococcus sp. (MRS) in Different Companion Animals and Determination of Risk Factors for Colonization with MRS. Antibiotics 2019, 8, 36. [Google Scholar] [CrossRef]
- Saputra, S.; Jordan, D.; Worthing, K.A.; Norris, J.M.; Wong, H.S.; Abraham, R.; Trott, D.J.; Abraham, S. Antimicrobial resistance in coagulase-positive staphylococci isolated from companion animals in Australia: A one year study. PLoS ONE 2017, 12, e0176379. [Google Scholar] [CrossRef]
- Couto, N.; Monchique, C.; Belas, A.; Marques, C.; Gama, L.T.; Pomba, C. Trends and molecular mechanisms of antimicrobial resistance in clinical staphylococci isolated from companion animals over a 16 year period. J. Antimicrob. Chemother. 2016, 71, 1479–1487. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; CLSI Supplement VET01S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Couto, N.; Belas, A.; Rodrigues, C.; Schwarz, S.; Pomba, C. Acquisition of the fexA and cfr genes in Staphylococcus pseudintermedius during florfenicol treatment of canine pyoderma. J. Glob. Antimicrob. Resist. 2016, 7, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Kwiecień, E.; Rzewuska, M.; Binek, M. Molecular characterization of high-level mupirocin resistance in methicillin-resistant staphylococci isolated from companion animals. Vet. Microbiol. 2021, 259, 109160. [Google Scholar] [CrossRef]
- Borio, S.; Colombo, S.; La Rosa, G.; De Lucia, M.; Damborg, P.; Guardabassi, L. Effectiveness of a combined (4% chlorhexidine digluconate shampoo and solution) protocol in MRS and non-MRS canine superficial pyoderma: A randomized, blinded, antibiotic-controlled study. Vet. Dermatol. 2015, 26, 339–344.e72. [Google Scholar] [CrossRef]
- Loeffler, A.; Cobb, M.A.; Bond, R. Comparison of a chlorhexidine and a benzoyl peroxide shampoo as sole treatment in canine superficial pyoderma. Vet. Rec. 2011, 169, 249. [Google Scholar] [CrossRef]
- Jeffers, J.G. Topical therapy for drug-resistant pyoderma in small animals. Vet. Clin. N. Am. Small Anim. Pract. 2013, 43, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ortega Morente, E.; Fernández-Fuentes, M.A.; Grande Burgos, M.J.; Abriouel, H.; Pérez Pulido, R.; Gálvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Schug, A.R.; Bartel, A.; Scholtzek, A.D.; Meurer, M.; Brombach, J.; Hensel, V.; Fanning, S.; Schwarz, S.; Feßler, A.T. Biocide susceptibility testing of bacteria: Development of a broth microdilution method. Vet. Microbiol. 2020, 248, 108791. [Google Scholar] [CrossRef]
- Langsrud, S.; Sidhu, M.; Heir, E.; Holck, A. Bacterial disinfectant resistance—A challenge for the food industry. Int. Biodeterior. Biodegrad. 2003, 51, 283–290. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Ussery, D.; Nielsen, L.N.; Ingmer, H. Review and phylogenetic analysis of qac genes that reduce susceptibility to quaternary ammonium compounds in Staphylococcus species. Eur. J. Microbiol. Immunol. 2015, 5, 44–61. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- CLSI. Understanding Susceptibility Test Data as a Component of Antimicrobial Stewardship in Veterinary Settings Performance, 1st ed.; CLSI Report VET09; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Assessing the antimicrobial susceptibility of bacteria obtained from animals. Vet. Microbiol. 2010, 141, 1–4. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Generation, Presentation, and Application of Antimicrobial Susceptibility Data for Bacteria of Animal Origin, a Report, 1st ed.; CLSI Document VET05-R; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Gómez-Sanz, E.; Kadlec, K.; Feßler, A.T.; Zarazaga, M.; Torres, C.; Schwarz, S. A novel fexA variant from a canine Staphylococcus pseudintermedius isolate that does not confer florfenicol resistance. Antimicrob. Agents Chemother. 2013, 57, 5763–5766. [Google Scholar] [CrossRef]
- Feßler, A.; Scott, C.; Kadlec, K.; Ehricht, R.; Monecke, S.; Schwarz, S. Characterization of methicillin-resistant Staphylococcus aureus ST398 from cases of bovine mastitis. J. Antimicrob. Chemother. 2010, 65, 619–625. [Google Scholar] [CrossRef]
- Schwarz, S.; Alesík, E.; Werckenthin, C.; Grobbel, M.; Lübke-Becker, A.; Wieler, L.H.; Wallmann, J. Antimicrobial susceptibility of coagulase-positive and coagulase-variable staphylococci from various indications of swine, dogs and cats as determined in the BfT-GermVet monitoring program 2004–2006. Berl. Münch. Tierärztl. Wochenschr. 2007, 120, 372–379. [Google Scholar]
- Walther, B.; Wieler, L.H.; Vincze, S.; Antão, E.M.; Brandenburg, A.; Stamm, I.; Kopp, P.A.; Kohn, B.; Semmler, T.; Lübke-Becker, A. MRSA variant in companion animals. Emerg. Infect. Dis. 2012, 18, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; Burnens, A.P.; Frey, J.; Kuhnert, P.; Nicolet, J. Molecular epidemiology and genetic linkage of macrolide and aminoglycoside resistance in Staphylococcus intermedius of canine origin. Vet. Microbiol. 2001, 79, 155–169. [Google Scholar] [CrossRef]
- EMEA. Commitee for Veterinary Medicinal Products Benzalkonium Chloride Summary Report. The European Agency for the Evaluation of Medicinal Products. EMEA/MRL/306/97. 1997. Available online: https://www.ema.europa.eu/en/documents/mrl-report/benzalkonium-chloride-summary-report-committee-veterinary-medicinal-products_en.pdf (accessed on 1 December 2021).
- Scholtzek, A.D.; Hanke, D.; Walther, B.; Eichhorn, I.; Stöckle, S.D.; Klein, K.S.; Gehlen, H.; Lübke-Becker, A.; Schwarz, S.; Feßler, A.T. Molecular Characterization of Equine Staphylococcus aureus Isolates Exhibiting Reduced Oxacillin Susceptibility. Toxins 2019, 11, 535. [Google Scholar] [CrossRef] [PubMed]
- Monecke, M.; Feßler, A.T.; Burgold-Voigt, S.; Krüger, H.; Mühldorfer, K.; Wibbelt, G.; Liebler-Tenorio, E.M.; Reinicke, M.; Braun, S.D.; Hanke, D.; et al. Staphylococcus aureus isolates from Eurasian Beavers (Castor fiber) carry a novel phage-borne bicomponent leukocidin related to the Panton-Valentine leukocidin. Sci. Rep. 2021, 11, 24394. [Google Scholar] [CrossRef]
- Roberts, M.C.; Feßler, A.T.; Monecke, S.; Ehricht, R.; No, D.; Schwarz, S. Molecular Analysis of Two Different MRSA Clones ST188 and ST3268 From Primates (Macaca spp.) in a United States Primate Center. Front Microbiol. 2018, 9, 2199. [Google Scholar] [CrossRef]
- Walther, B.; Klein, K.S.; Barton, A.K.; Semmler, T.; Huber, C.; Merle, R.; Tedin, K.; Mitrach, F.; Lübke-Becker, A.; Gehlen, H. Equine Methicillin-Resistant Sequence Type 398 Staphylococcus aureus (MRSA) Harbor Mobile Genetic Elements Promoting Host Adaptation. Front. Microbiol. 2018, 9, 2516. [Google Scholar] [CrossRef]
- Murugaiyan, J.; Walther, B.; Stamm, I.; Abou-Elnaga, Y.; Brueggemann-Schwarze, S.; Vincze, S.; Wieler, L.H.; Lübke-Becker, A.; Semmler, T.; Roesler, U. Species differentiation within the Staphylococcus intermedius group using a refined MALDI-TOF MS database. Clin. Microbiol. Infect. 2014, 20, 1007–1015. [Google Scholar] [CrossRef]
- Schnellmann, C.; Gerber, V.; Rossano, A.; Jaquier, V.; Panchaud, Y.; Doherr, M.G.; Thomann, A.; Straub, R.; Perreten, V. Presence of new mecA and mph(C) variants conferring antibiotic resistance in Staphylococcus spp. isolated from the skin of horses before and after clinic admission. J. Clin. Microbiol. 2006, 44, 4444–4454. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of staphylococcal cassette chromosome mec type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef]
- Pang, Y.; Bosch, T.; Roberts, M.C. Single polymerase chain reaction for the detection of tetracycline-resistant determinants Tet K and Tet L. Mol. Cell. Probes 1994, 8, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C.; Pang, Y.; Riley, D.E.; Hillier, S.L.; Berger, R.C.; Krieger, J.N. Detection of Tet M and Tet O tetracycline resistance genes by polymerase chain reaction. Mol. Cell. Probes 1993, 7, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Lüthje, P.; Schwarz, S. Molecular basis of resistance to macrolides and lincosamides among staphylococci and streptococci from various animal sources collected in the resistance monitoring program BfT-GermVet. Int. J. Antimicrob. Agents 2007, 29, 528–535. [Google Scholar] [CrossRef]
- Li, J.; Li, B.; Wendlandt, S.; Schwarz, S.; Wang, Y.; Wu, C.; Ma, Z.; Shen, J. Identification of a novel vga(E) gene variant that confers resistance to pleuromutilins, lincosamides and streptogramin A antibiotics in staphylococci of porcine origin. J. Antimicrob. Chemother. 2014, 69, 919–923. [Google Scholar] [CrossRef]
- Hauschild, T.; Vuković, D.; Dakić, I.; Jezek, P.; Djukić, S.; Dimitrijević, V.; Stepanović, S.; Schwarz, S. Aminoglycoside resistance in members of the Staphylococcus sciuri group. Microb. Drug Resist. 2007, 13, 77–84. [Google Scholar] [CrossRef]
- López, M.; Kadlec, K.; Schwarz, S.; Torres, C. First detection of the staphylococcal trimethoprim resistance gene dfrK and the dfrK-carrying transposon Tn559 in enterococci. Microb. Drug Resist. 2012, 13, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Huynh, T.M.; Bourdon, N.; Auzou, M.; Leclercq, R. Trimethoprim resistance genes in vancomycin-resistant Enterococcus faecium clinical isolates from France. Int. J. Antimicrob. Agents 2009, 34, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antimicrobial Agent(s) | Number of Isolates for Which the MIC Value (mg/L) Is | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |
| Penicillin G | 5 | 5 | - | - | 5 | 1 | 3 | 4 | 6 | 3 | 5 | 3 | 3 | |||||
| Ampicillin | 6 | 3 | 6 | 2 | 2 | 7 | 5 | 4 | 3 | 5 | - | - | ||||||
| Amoxicillin-clavulanic acid (2:1) | 1 | 1 | 5 | 11 | 2 | 8 | 3 | 4 | - | 6 | 2 | - | - | |||||
| Oxacillin | - | - | 4 | 2 | 8 | 12 | 3 | - | - | 1 | 13 | |||||||
| Cephalothin | - | 6 | 13 | 10 | 3 | 2 | 1 | 1 | 4 | 2 | - | 1 | - | |||||
| Cefotaxime | - | - | - | - | 1 | 2 | 1 | 12 | 13 | 3 | 2 | 1 | 8 | |||||
| Cefoperazone | - | - | - | 3 | 2 | 15 | 9 | 4 | 2 | - | 8 | |||||||
| Ceftiofur | - | - | - | 3 | - | 20 | 6 | 5 | 1 | - | 1 | 3 | 4 | |||||
| Florfenicol | - | - | - | - | 19 | 21 | 3 | - | - | - | - | - | - | |||||
| Erythromycin | - | - | - | 1 | 17 | 11 | 2 | - | - | 1 | - | - | 10 | |||||
| Tylosin | - | - | - | 1 | 10 | 21 | 3 | - | - | - | - | - | 8 | |||||
| Tilmicosin | - | - | - | 1 | 15 | 18 | 1 | - | - | - | - | - | 8 | |||||
| Clindamycin | - | 7 | 22 | 5 | - | - | - | - | - | - | - | 1 | 8 | |||||
| Streptomycin | - | - | - | 1 | 4 | 24 | 11 | 1 | - | - | 1 | - | 1 | |||||
| Gentamicin | 2 | 19 | 16 | 5 | - | - | - | - | 1 | - | - | - | - | |||||
| Neomycin | 4 | 14 | 17 | 5 | - | - | - | 1 | - | 2 | - | |||||||
| Ciprofloxacin | - | - | - | - | 3 | 11 | 16 | 2 | 1 | - | 1 | 3 | 6 | |||||
| Enrofloxacin | - | - | - | 2 | 18 | 10 | 1 | 2 | 1 | 3 | 1 | - | 5 | |||||
| Marbofloxacin | - | - | - | - | - | 12 | 19 | 2 | - | - | 1 | 4 | 5 | |||||
| Nalidixic acid | - | - | - | - | - | - | - | - | 7 | 13 | 11 | 5 | 7 | |||||
| Trimethoprim-sulfamethoxazole (1:19) | - | - | 30 | 7 | 1 | 2 | 1 | 1 | - | 1 | - | - | - | |||||
| Tetracycline | 17 | 20 | 1 | 1 | - | - | - | 1 | 1 | 1 | 1 | - | - | |||||
| Doxycycline | 12 | 23 | 2 | 1 | 2 | 1 | - | 2 | - | - | - | - | - | |||||
| Linezolid | - | - | - | - | 6 | 28 | 7 | 2 | - | - | - | - | - | |||||
| Tiamulin | - | - | - | 7 | 18 | 17 | - | - | - | - | - | - | 1 | |||||
| Vancomycin | - | - | - | - | 1 | 9 | 32 | 1 | - | - | - | - | - | |||||
| Quinupristin-dalfopristin | - | - | - | 2 | 12 | 24 | 4 | - | 1 | - | - | |||||||
| Antimicrobial Agent (s) | Number of Isolates for Which the MIC Value (mg/L) Is | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |
| Penicillin G | - | 2 | - | - | 2 | 1 | 1 | - | - | 4 | 5 | 4 | - | |||||
| Ampicillin | - | - | 2 | 2 | 2 | 2 | 1 | - | 4 | 2 | 4 | - | - | |||||
| Amoxicillin-clavulanic acid (2:1) | - | - | - | 7 | - | 4 | 1 | 1 | 1 | 3 | 2 | - | - | |||||
| Oxacillin | - | - | - | 2 | 5 | 4 | 1 | - | - | - | 7 | |||||||
| Cephalothin | - | 2 | 6 | 4 | 1 | 1 | 1 | 1 | - | 1 | - | 2 | - | |||||
| Cefotaxime | - | - | - | - | - | 2 | - | 5 | 5 | 1 | 1 | 1 | 4 | |||||
| Cefoperazone | - | - | - | 2 | - | 7 | 3 | 2 | - | - | 5 | |||||||
| Ceftiofur | - | - | - | 2 | - | 6 | 5 | 1 | - | 1 | - | 2 | 2 | |||||
| Florfenicol | - | - | - | - | 9 | 8 | 2 | - | - | - | - | - | - | |||||
| Erythromycin | - | - | - | - | 6 | 7 | - | - | - | - | - | - | 6 | |||||
| Tylosin | - | - | - | - | 2 | 13 | - | - | 1 | - | - | - | 3 | |||||
| Tilmicosin | - | - | - | - | 3 | 11 | 1 | 1 | - | - | - | - | 3 | |||||
| Clindamycin | 1 | 2 | 11 | 1 | - | - | - | - | 1 | 1 | - | - | 2 | |||||
| Streptomycin | - | - | - | - | 1 | 9 | 6 | - | - | 1 | 1 | - | 1 | |||||
| Gentamicin | 2 | 9 | 4 | 1 | - | - | - | - | 2 | - | 1 | - | - | |||||
| Neomycin | - | 7 | 6 | 2 | - | - | 1 | 2 | - | - | 1 | |||||||
| Ciprofloxacin | - | - | - | - | 1 | 6 | 7 | - | - | - | - | 1 | 4 | |||||
| Enrofloxacin | - | - | - | - | 13 | 1 | - | - | - | 2 | 1 | 1 | 1 | |||||
| Marbofloxacin | - | - | - | - | - | 7 | 7 | - | - | - | - | 1 | 4 | |||||
| Nalidixic acid | - | - | - | - | - | - | - | - | 2 | 11 | 2 | 1 | 3 | |||||
| Trimethoprim-sulfamethoxazole (1:19) | - | - | 15 | 2 | 1 | - | 1 | - | - | - | - | - | - | |||||
| Tetracycline | 10 | 5 | - | 1 | - | - | - | - | 2 | 1 | - | - | - | |||||
| Doxycycline | 10 | 5 | - | 1 | - | 2 | - | 1 | - | - | - | - | - | |||||
| Linezolid | - | - | - | - | 3 | 14 | 1 | 1 | - | - | - | - | - | |||||
| Tiamulin | - | - | 3 | 1 | 7 | 8 | - | - | - | - | - | - | - | |||||
| Vancomycin | - | - | - | - | - | 1 | 16 | 2 | - | - | - | - | - | |||||
| Quinupristin-dalfopristin | - | - | - | - | 4 | 14 | 1 | - | - | - | - | |||||||
| Antimicrobial Agent (s) | Number of Isolates for Which the MIC Value (mg/L) Is | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |
| Penicillin G | 1 | - | - | - | - | 1 | - | 2 | 1 | 2 | 1 | - | 3 | |||||
| Ampicillin | - | 1 | - | 2 | 1 | 3 | 1 | - | - | 1 | - | 2 | - | |||||
| Amoxicillin-clavulanic acid (2:1) | - | - | 2 | 6 | - | 1 | - | - | 1 | - | 1 | - | - | |||||
| Oxacillin | - | - | 1 | 2 | 5 | - | - | - | 1 | - | 2 | |||||||
| Cephalothin | 1 | 6 | 1 | 1 | - | - | - | 1 | - | - | 1 | - | - | |||||
| Cefotaxime | - | - | - | - | 4 | 4 | - | - | 1 | - | - | - | 2 | |||||
| Cefoperazone | - | - | - | 7 | 1 | 1 | - | 1 | - | - | 1 | |||||||
| Ceftiofur | - | - | - | 6 | 2 | - | - | 1 | - | - | - | 1 | 1 | |||||
| Florfenicol | - | - | - | - | 7 | 4 | - | - | - | - | - | - | - | |||||
| Erythromycin | - | - | - | - | 5 | - | - | - | - | - | - | - | 6 | |||||
| Tylosin | - | - | - | 4 | 1 | - | - | - | - | - | - | - | 6 | |||||
| Tilmicosin | - | - | - | 1 | 4 | - | - | - | - | - | - | - | 6 | |||||
| Clindamycin | - | - | 5 | - | - | - | - | 1 | - | - | 1 | 1 | 3 | |||||
| Streptomycin | - | - | - | 1 | 4 | - | - | - | 1 | 2 | 1 | - | 2 | |||||
| Gentamicin | 7 | 2 | - | - | - | - | - | 1 | 1 | - | - | - | - | |||||
| Neomycin | 4 | 1 | - | - | - | - | 4 | 2 | - | - | - | |||||||
| Ciprofloxacin | - | - | - | 2 | 4 | 3 | - | - | - | - | - | - | 2 | |||||
| Enrofloxacin | - | - | - | 3 | 6 | - | - | - | - | - | - | 1 | 1 | |||||
| Marbofloxacin | - | - | - | - | 1 | 7 | 1 | - | - | - | - | 1 | 1 | |||||
| Nalidixic acid | - | - | - | - | - | - | - | - | 3 | 6 | - | 2 | - | |||||
| Trimethoprim-sulfamethoxazole (1:19) | - | - | - | 6 | - | 3 | - | - | - | 1 | 1 | - | - | |||||
| Tetracycline | 5 | 2 | - | - | - | - | - | - | 2 | 2 | - | - | - | |||||
| Doxycycline | 6 | 1 | - | - | - | - | 2 | 2 | - | - | - | - | - | |||||
| Linezolid | - | - | - | - | 1 | 10 | - | - | - | - | - | - | - | |||||
| Tiamulin | - | 1 | 7 | 3 | - | - | - | - | - | - | - | - | - | |||||
| Vancomycin | - | - | - | - | - | - | 10 | 1 | - | - | - | - | - | |||||
| Quinupristin-dalfopristin | - | - | - | 2 | 8 | 1 | - | - | - | - | - | - | - | |||||
| Antimicrobial Agent (s) | Number of Isolates for Which the MIC Value (mg/L) Is | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |
| Penicillin G | 8 | 4 | - | - | 2 | 3 | 4 | 2 | 5 | 4 | 7 | 4 | 3 | 3 | ||||
| Ampicillin | 2 | 2 | 2 | 4 | 10 | 13 | 3 | - | 2 | - | 1 | 2 | - | |||||
| Amoxicillin-clavulanic acid (2:1) | 1 | 1 | 13 | 21 | - | - | 2 | - | 1 | 1 | 1 | - | - | |||||
| Oxacillin | - | - | 1 | 16 | 19 | - | - | - | - | 1 | 4 | |||||||
| Cephalothin | 9 | 27 | - | 1 | 1 | - | 1 | - | - | - | 2 | - | - | |||||
| Cefotaxime | - | - | - | 1 | 14 | 21 | - | - | - | 1 | 1 | - | 3 | |||||
| Cefoperazone | - | - | 4 | 32 | - | - | 2 | 1 | - | - | 2 | |||||||
| Ceftiofur | - | - | 2 | 34 | - | - | - | 1 | 1 | - | - | 1 | 2 | |||||
| Florfenicol | - | - | - | - | 34 | 7 | - | - | - | - | - | - | - | |||||
| Erythromycin | - | - | - | 4 | 29 | - | - | - | - | - | - | - | 8 | |||||
| Tylosin | - | - | - | 31 | 2 | - | - | - | - | 1 | - | 1 | 6 | |||||
| Tilmicosin | - | - | - | 3 | 31 | - | - | - | 1 | - | - | - | 6 | |||||
| Clindamycin | - | 7 | 27 | 1 | - | - | 1 | - | - | 1 | 1 | - | 3 | |||||
| Streptomycin | - | - | - | 2 | 20 | 11 | - | - | - | 2 | 2 | - | 4 | |||||
| Gentamicin | 22 | 15 | - | - | - | - | 2 | 1 | 1 | - | - | - | - | |||||
| Neomycin | 20 | 13 | - | - | - | - | 3 | 5 | - | - | - | |||||||
| Ciprofloxacin | - | 1 | 2 | 3 | 21 | 4 | 3 | - | - | - | 1 | 2 | 4 | |||||
| Enrofloxacin | - | 1 | 3 | 6 | 20 | 2 | 2 | - | - | - | 3 | 2 | 2 | |||||
| Marbofloxacin | - | - | - | 3 | 3 | 22 | 6 | - | - | - | - | 3 | 4 | |||||
| Nalidixic acid | - | - | - | - | - | - | - | 3 | 11 | 18 | 2 | 5 | 2 | |||||
| Trimethoprim-sulfamethoxazole (1:19) | - | 1 | 9 | 13 | 2 | 13 | - | - | - | 2 | 1 | - | - | |||||
| Tetracycline | 18 | 13 | - | - | - | - | - | - | 5 | 5 | - | - | - | |||||
| Doxycycline | 27 | 4 | - | - | - | - | 4 | 6 | - | - | - | - | - | |||||
| Linezolid | - | - | - | - | 5 | 35 | 1 | - | - | - | - | - | - | |||||
| Tiamulin | - | 1 | 22 | 17 | 1 | - | - | - | - | - | - | - | - | |||||
| Vancomycin | - | - | - | - | - | 6 | 34 | 1 | - | - | - | - | - | |||||
| Quinupristin-dalfopristin | - | - | - | 5 | 31 | 5 | - | - | - | - | - | - | - | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feßler, A.T.; Scholtzek, A.D.; Schug, A.R.; Kohn, B.; Weingart, C.; Schink, A.-K.; Bethe, A.; Lübke-Becker, A.; Schwarz, S. Antimicrobial and Biocide Resistance among Feline and Canine Staphylococcus aureus and Staphylococcus pseudintermedius Isolates from Diagnostic Submissions. Antibiotics 2022, 11, 127. https://doi.org/10.3390/antibiotics11020127
Feßler AT, Scholtzek AD, Schug AR, Kohn B, Weingart C, Schink A-K, Bethe A, Lübke-Becker A, Schwarz S. Antimicrobial and Biocide Resistance among Feline and Canine Staphylococcus aureus and Staphylococcus pseudintermedius Isolates from Diagnostic Submissions. Antibiotics. 2022; 11(2):127. https://doi.org/10.3390/antibiotics11020127
Chicago/Turabian StyleFeßler, Andrea T., Anissa D. Scholtzek, Angela R. Schug, Barbara Kohn, Christiane Weingart, Anne-Kathrin Schink, Astrid Bethe, Antina Lübke-Becker, and Stefan Schwarz. 2022. "Antimicrobial and Biocide Resistance among Feline and Canine Staphylococcus aureus and Staphylococcus pseudintermedius Isolates from Diagnostic Submissions" Antibiotics 11, no. 2: 127. https://doi.org/10.3390/antibiotics11020127
APA StyleFeßler, A. T., Scholtzek, A. D., Schug, A. R., Kohn, B., Weingart, C., Schink, A.-K., Bethe, A., Lübke-Becker, A., & Schwarz, S. (2022). Antimicrobial and Biocide Resistance among Feline and Canine Staphylococcus aureus and Staphylococcus pseudintermedius Isolates from Diagnostic Submissions. Antibiotics, 11(2), 127. https://doi.org/10.3390/antibiotics11020127