
Molecular Determinants for OMF Selectivity in Tripartite RND Multidrug Efflux Systems

 , ,
, ,  , , ,
, , ,  ,
,
Abstract
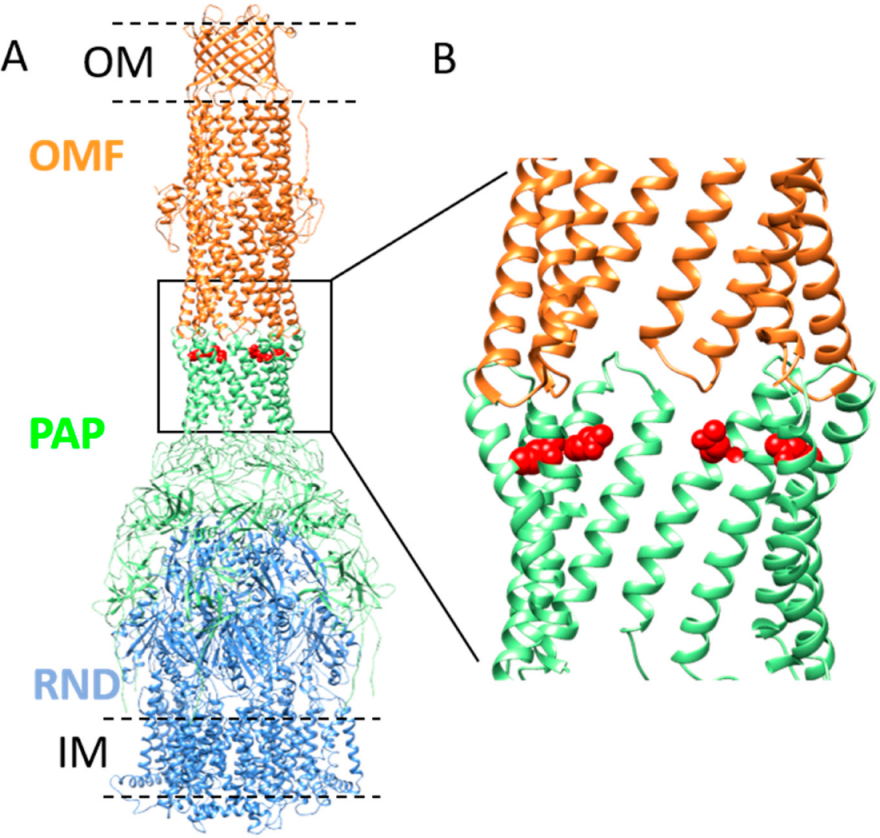
1. Introduction
2. Results
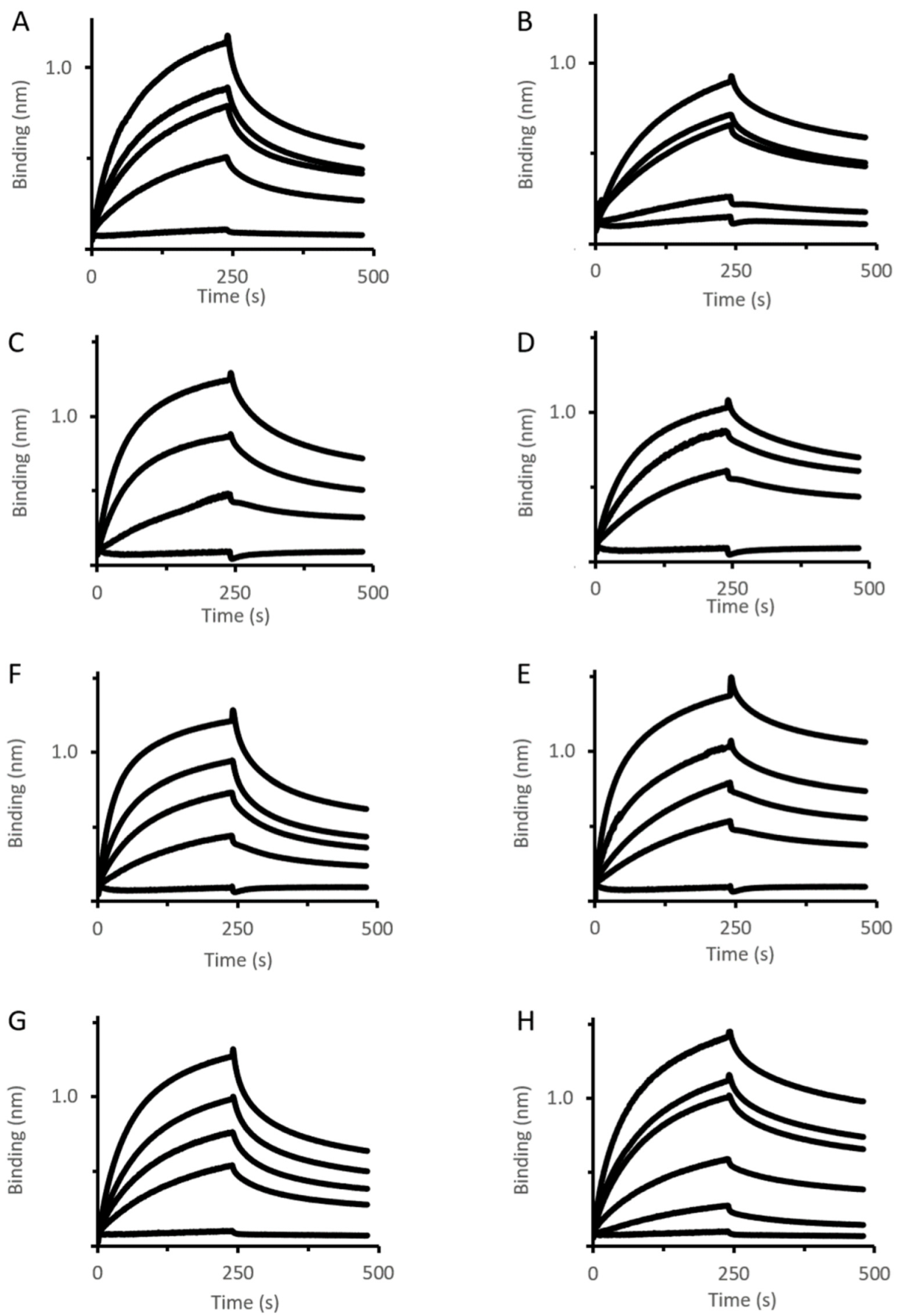
2.1. Analysis of Mexa Binding to OMF by Biolayer Interferometry
2.2. Analysis of Oligomerization State of MexAQ93R
2.3. Binding Analysis of MexA Variants to MexB Using BLI
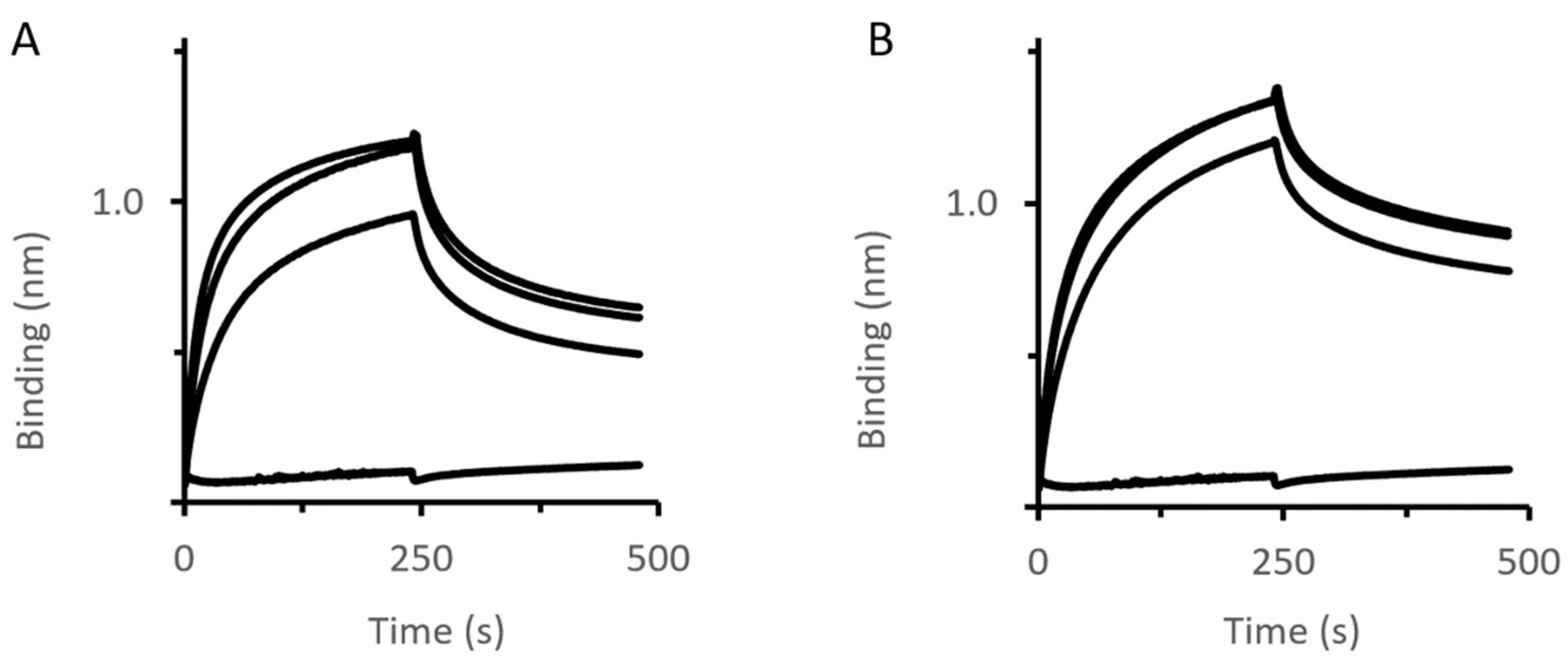
2.4. Impact of MexAQ93R on the Formation of Tripartite Complexes
2.5. Impact of an OprM Variant on the Formation of Tripartite Complexes
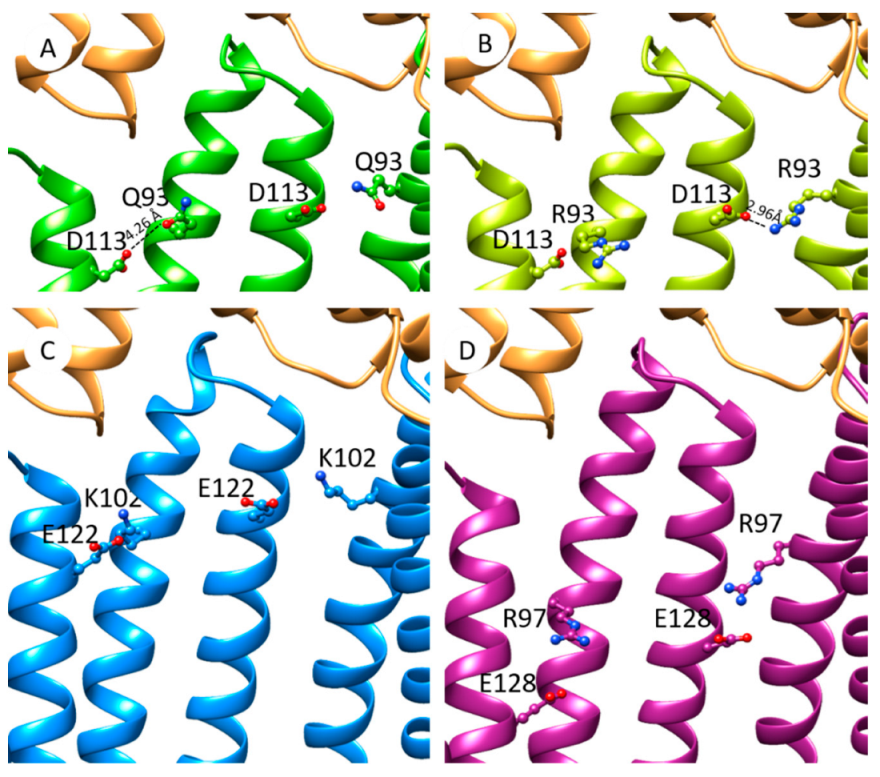
2.6. The Increase in Antibiotic Resistance Is Related to a Q93R Mutation When Associated with D113 Residue
3. Discussion
4. Materials and Methods
4.1. Material and Reagents
4.2. Protein Preparation
4.3. Membrane Protein Stabilization with Amphipols
4.4. Binding Analysis Using BLI
4.5. Formation of Tripartite Complexes
4.6. Analysis of MexA Oligomerisation State
4.7. Electron Microscopy Acquisition and Image Analysis
4.8. Model Simulation and Score Evaluation
4.9. Measurement of Antibiotic Susceptibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zgurskaya, H.I.; Nikaido, H. Bypassing the Periplasm: Reconstitution of the AcrAB Multidrug Efflux Pump of Escherichia Coli. Proc. Natl. Acad. Sci. USA 1999, 96, 7190–7195. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Venter, H. Reversing Resistance to Counter Antimicrobial Resistance in the World Health Organisation’s Critical Priority of Most Dangerous Pathogens. Biosci. Rep. 2019, 39, BSR20180474. [Google Scholar] [CrossRef]
- Symmons, M.F.; Marshall, R.L.; Bavro, V.N. Architecture and Roles of Periplasmic Adaptor Proteins in Tripartite Efflux Assemblies. Front. Microbiol. 2015, 6, 513. [Google Scholar] [CrossRef] [PubMed]
- Alav, I.; Kobylka, J.; Kuth, M.S.; Pos, K.M.; Picard, M.; Blair, J.M.A.; Bavro, V.N. Structure, Assembly, and Function of Tripartite Efflux and Type 1 Secretion Systems in Gram-Negative Bacteria. Chem. Rev. 2021, 121, 5479–5596. [Google Scholar] [CrossRef]
- Schweizer, H.P. Efflux as a Mechanism of Resistance to Antimicrobials in Pseudomonas Aeruginosa and Related Bacteria: Unanswered Questions. Genet. Mol. Res. 2003, 2, 48–62. [Google Scholar]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate Specificities of MexAB-OprM, MexCD-OprJ, and MexXY-OprM Efflux Pumps in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef]
- Maseda, H.; Yoneyama, H.; Nakae, T. Assignment of the Substrate-Selective Subunits of the MexEF-OprN Multidrug Efflux Pump of Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 658–664. [Google Scholar] [CrossRef]
- Phan, G.; Picard, M.; Broutin, I. Focus on the Outer Membrane Factor OprM, the Forgotten Player from Efflux Pumps Assemblies. Antibiotics 2015, 4, 544–566. [Google Scholar] [CrossRef]
- Srikumar, R.; Li, X.Z.; Poole, K. Inner Membrane Efflux Components Are Responsible for Beta-Lactam Specificity of Multidrug Efflux Pumps in Pseudomonas Aeruginosa. J. Bacteriol. 1997, 179, 7875–7881. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, H.; Ocaktan, A.; Gotoh, N.; Nishino, T.; Nakae, T. Subunit Swapping in the Mex-Extrusion Pumps in Pseudomonas Aeruginosa. Biochem. Biophys. Res. Commun. 1998, 244, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Sykes, E.M.E.; Li, Y.; Kumar, A. MexXY RND Pump of Pseudomonas Aeruginosa PA7 Effluxes Bi-Anionic β-Lactams Carbenicillin and Sulbenicillin When It Partners with the Outer Membrane Factor OprA but Not with OprM. Microbiology 2020, 166, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Vediyappan, G.; Borisova, T.; Fralick, J.A. Isolation and Characterization of VceC Gain-of-Function Mutants That Can Function with the AcrAB Multiple-Drug-Resistant Efflux Pump of Escherichia Coli. J. Bacteriol. 2006, 188, 3757–3762. [Google Scholar] [CrossRef]
- Bai, J.; Mosley, L.; Fralick, J.A. Evidence That the C-Terminus of OprM Is Involved in the Assembly of the VceAB-OprM Efflux Pump. FEBS Lett. 2010, 584, 1493–1497. [Google Scholar] [CrossRef]
- Weeks, J.W.; Nickels, L.M.; Ntreh, A.T.; Zgurskaya, H.I. Non-Equivalent Roles of Two Periplasmic Subunits in the Function and Assembly of Triclosan Pump TriABC from Pseudomonas Aeruginosa. Mol. Microbiol. 2015, 98, 343–356. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, G.; Hryc, C.F.; Blaza, J.N.; Serysheva, I.I.; Schmid, M.F.; Chiu, W.; Luisi, B.F.; Du, D. An Allosteric Transport Mechanism for the AcrAB-TolC Multidrug Efflux Pump. elife 2017, 6, e24905. [Google Scholar] [CrossRef]
- Tsutsumi, K.; Yonehara, R.; Ishizaka-Ikeda, E.; Miyazaki, N.; Maeda, S.; Iwasaki, K.; Nakagawa, A.; Yamashita, E. Structures of the Wild-Type MexAB-OprM Tripartite Pump Reveal Its Complex Formation and Drug Efflux Mechanism. Nat. Commun. 2019, 10, 1520. [Google Scholar] [CrossRef]
- Glavier, M.; Puvanendran, D.; Salvador, D.; Decossas, M.; Phan, G.; Garnier, C.; Frezza, E.; Cece, Q.; Schoehn, G.; Picard, M.; et al. Antibiotic Export by MexB Multidrug Efflux Transporter Is Allosterically Controlled by a MexA-OprM Chaperone-like Complex. Nat. Commun. 2020, 11, 4948. [Google Scholar] [CrossRef]
- Lobedanz, S.; Bokma, E.; Symmons, M.F.; Koronakis, E.; Hughes, C.; Koronakis, V. A Periplasmic Coiled-Coil Interface Underlying TolC Recruitment and the Assembly of Bacterial Drug Efflux Pumps. Proc. Natl. Acad. Sci. USA 2007, 104, 4612–4617. [Google Scholar] [CrossRef]
- Bokma, E.; Koronakis, E.; Lobedanz, S.; Hughes, C.; Koronakis, V. Directed Evolution of a Bacterial Efflux Pump: Adaptation of the E. Coli TolC Exit Duct to the Pseudomonas MexAB Translocase. FEBS Lett. 2006, 580, 5339–5343. [Google Scholar] [CrossRef]
- Krishnamoorthy, G.; Tikhonova, E.B.; Zgurskaya, H.I. Fitting Periplasmic Membrane Fusion Proteins to Inner Membrane Transporters: Mutations That Enable Escherichia Coli AcrA to Function with Pseudomonas Aeruginosa MexB. J. Bacteriol. 2008, 190, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.L.; Bavro, V.N. Mutations in the TolC Periplasmic Domain Affect Substrate Specificity of the AcrAB-TolC Pump. Front. Mol. Biosci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Tikhonova, E.B.; Yamada, Y.; Zgurskaya, H.I. Sequential Mechanism of Assembly of Multidrug Efflux Pump AcrAB-TolC. Chem. Biol. 2011, 18, 454–463. [Google Scholar] [CrossRef]
- Weeks, J.W.; Celaya-Kolb, T.; Pecora, S.; Misra, R. AcrA Suppressor Alterations Reverse the Drug Hypersensitivity Phenotype of a TolC Mutant by Inducing TolC Aperture Opening. Mol. Microbiol. 2010, 75, 1468–1483. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hayashi, K.; Nakashima, R.; Sakurai, K.; Kitagawa, K.; Yamasaki, S.; Nishino, K.; Yamaguchi, A. AcrB-AcrA Fusion Proteins That Act as Multidrug Efflux Transporters. J. Bacteriol. 2016, 198, 332–342. [Google Scholar] [CrossRef]
- Stegmeier, J.F.; Polleichtner, G.; Brandes, N.; Hotz, C.; Andersen, C. Importance of the Adaptor (Membrane Fusion) Protein Hairpin Domain for the Functionality of Multidrug Efflux Pumps. Biochemistry 2006, 45, 10303–10312. [Google Scholar] [CrossRef] [PubMed]
- Symmons, M.F.; Bokma, E.; Koronakis, E.; Hughes, C.; Koronakis, V. The Assembled Structure of a Complete Tripartite Bacterial Multidrug Efflux Pump. Proc. Natl. Acad. Sci. USA 2009, 106, 7173–7178. [Google Scholar] [CrossRef] [PubMed]
- Eda, S.; Maseda, H.; Yoshihara, E.; Nakae, T. Assignment of the Outer-Membrane-Subunit-Selective Domain of the Membrane Fusion Protein in the Tripartite Xenobiotic Efflux Pump of Pseudomonas Aeruginosa. FEMS Microbiol. Lett. 2006, 254, 101–107. [Google Scholar] [CrossRef]
- Ferrandez, Y.; Monlezun, L.; Phan, G.; Benabdelhak, H.; Benas, P.; Ulryck, N.; Falson, P.; Ducruix, A.; Picard, M.; Broutin, I. Stoichiometry of the MexA-OprM Binding, as Investigated by Blue Native Gel Electrophoresis. Electrophoresis 2012, 33, 1282–1287. [Google Scholar] [CrossRef]
- Akama, H.; Matsuura, T.; Kashiwagi, S.; Yoneyama, H.; Narita, S.-I.; Tsukihara, T.; Nakagawa, A.; Nakae, T. Crystal Structure of the Membrane Fusion Protein, MexA, of the Multidrug Transporter in Pseudomonas Aeruginosa. J. Biol. Chem. 2004, 279, 25939–25942. [Google Scholar] [CrossRef] [PubMed]
- Higgins, M.K.; Bokma, E.; Koronakis, E.; Hughes, C.; Koronakis, V. Structure of the Periplasmic Component of a Bacterial Drug Efflux Pump. Proc. Natl. Acad. Sci. USA 2004, 101, 9994–9999. [Google Scholar] [CrossRef]
- Daury, L.; Orange, F.; Taveau, J.-C.; Verchère, A.; Monlezun, L.; Gounou, C.; Marreddy, R.K.R.; Picard, M.; Broutin, I.; Pos, K.M.; et al. Tripartite Assembly of RND Multidrug Efflux Pumps. Nat. Commun. 2016, 7, 10731. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein Structure and Function Prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y. I-TASSER Server: New Development for Protein Structure and Function Predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A Unified Platform for Automated Protein Structure and Function Prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for Rigid and Symmetric Docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef]
- Yoshihara, E.; Eda, S.; Kaitou, S. Functional Interaction Sites of OprM with MexAB in the Pseudomonas Aeruginosa Multidrug Efflux Pump. FEMS Microbiol. Lett. 2009, 299, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Bhagavathi, R.; Tran, P.; Muzzarelli, K.; Wang, D.; Fralick, J.A. Evidence That the C-Terminal Region Is Involved in the Stability and Functionality of OprM in E. coli. Microbiol. Res. 2014, 169, 425–431. [Google Scholar] [CrossRef]
- Yamanaka, H.; Izawa, H.; Okamoto, K. Carboxy-Terminal Region Involved in Activity of Escherichia Coli TolC. J. Bacteriol. 2001, 183, 6961–6964. [Google Scholar] [CrossRef]
- Yamanaka, H.; Nomura, T.; Morisada, N.; Shinoda, S.; Okamoto, K. Site-Directed Mutagenesis Studies of the Amino Acid Residue at Position 412 of Escherichia Coli TolC Which Is Required for the Activity. Microb. Pathog. 2002, 33, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Ntsogo Enguéné, Y.V.; Phan, G.; Garnier, C.; Ducruix, A.; Prangé, T.; Broutin, I. Xenon for Tunnelling Analysis of the Efflux Pump Component OprN. PLoS ONE 2017, 12, e0184045. [Google Scholar] [CrossRef] [PubMed]
- Bosco, M.; Damian, M.; Chauhan, V.; Roche, M.; Guillet, P.; Fehrentz, J.-A.; Bonneté, F.; Polidori, A.; Banères, J.-L.; Durand, G. Biotinylated Non-Ionic Amphipols for GPCR Ligands Screening. Methods 2020, 180, 69–78. [Google Scholar] [CrossRef]
- Rouser, G.; Fleischer, S.; Yamamoto, A. Two Dimensional Then Layer Chromatographic Separation of Polar Lipids and Determination of Phospholipids by Phosphorus Analysis of Spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef]
- Tang, G.; Peng, L.; Baldwin, P.R.; Mann, D.S.; Jiang, W.; Rees, I.; Ludtke, S.J. EMAN2: An Extensible Image Processing Suite for Electron Microscopy. J. Struct. Biol. 2007, 157, 38–46. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. Geometry-Based Flexible and Symmetric Protein Docking. Proteins 2005, 60, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Blundell, T.L. Comparative Protein Modelling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Analyte | koff (10−3 s−1) | kon (102 M−1s−1) | KD (µM) |
|---|---|---|---|---|
| OprMwt | MexAwt | 2.15 | 1.80 | 12.0 |
| OprM∆473−485 | MexAwt | 4.58 | 1.03 | 44.0 |
| OprN | MexAwt | 3.58 | 1.77 | 20.0 |
| TolC | MexAwt | 5.8 | 1.27 | 45.8 |
| OprMwt | MexAQ93R | 2.66 | 0.81 | 32.9 |
| OprM∆473−485 | MexAQ93R | 2.63 | 1.02 | 25.7 |
| OprN | MexAQ93R | 2.38 | 0.88 | 26.9 |
| TolC | MexAQ93R | 1.91 | 1.08 | 17.8 |
| Ligand | Analyte | koff (10−3 s−1) | kon (102 M−1s−1) | KD (µM) |
|---|---|---|---|---|
| MexB | MexAwt | 5.5 | 2.50 | 23.0 |
| MexB | MexAQ93R | 3.0 | 1.73 | 17.4 |
| PAP | ||
|---|---|---|
| OMF | MexAwt | MexAQ93R |
| OprMwt | 1146 ± 59 | 1981 ± 156 *a |
| OprM∆473−485 | 0 | 589 ± 15 *b, **c |
| OprN | 0 | 10 ± 0.3 **b |
| TolC | 0 | 164 ± 3 **b |
| Minimal Inhibitory Concentration (MIC, µg/mL) | ||
|---|---|---|
| Strain | Ticarcillin | Aztreonam |
| PAO1 | 32 | 4 |
| PAO1 pUCP24-mexAB-oprM wt | 64 | 8 |
| PAO1 pUCP24-mexA D113A mexB-oprM | 32 | 4 |
| PAO1 pUCP24-mexA Q93R mexB-oprM | 128 | 32 |
| PAO1 pUCP24-mexA D113A + Q93R mexB-oprM | 32 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyer, E.; Dessolin, J.; Lustig, M.; Decossas, M.; Phan, G.; Cece, Q.; Durand, G.; Dubois, V.; Sansen, J.; Taveau, J.-C.; et al. Molecular Determinants for OMF Selectivity in Tripartite RND Multidrug Efflux Systems. Antibiotics 2022, 11, 126. https://doi.org/10.3390/antibiotics11020126
Boyer E, Dessolin J, Lustig M, Decossas M, Phan G, Cece Q, Durand G, Dubois V, Sansen J, Taveau J-C, et al. Molecular Determinants for OMF Selectivity in Tripartite RND Multidrug Efflux Systems. Antibiotics. 2022; 11(2):126. https://doi.org/10.3390/antibiotics11020126
Chicago/Turabian StyleBoyer, Esther, Jean Dessolin, Margaux Lustig, Marion Decossas, Gilles Phan, Quentin Cece, Grégory Durand, Véronique Dubois, Joris Sansen, Jean-Christophe Taveau, and et al. 2022. "Molecular Determinants for OMF Selectivity in Tripartite RND Multidrug Efflux Systems" Antibiotics 11, no. 2: 126. https://doi.org/10.3390/antibiotics11020126
APA StyleBoyer, E., Dessolin, J., Lustig, M., Decossas, M., Phan, G., Cece, Q., Durand, G., Dubois, V., Sansen, J., Taveau, J.-C., Broutin, I., Daury, L., & Lambert, O. (2022). Molecular Determinants for OMF Selectivity in Tripartite RND Multidrug Efflux Systems. Antibiotics, 11(2), 126. https://doi.org/10.3390/antibiotics11020126