

Combined Anti-Bacterial Actions of Lincomycin and Freshly Prepared Silver Nanoparticles: Overcoming the Resistance to Antibiotics and Enhancement of the Bioactivity


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials Procurement
2.2. Isolation and Identification of Bacterial Isolates
2.3. Antibiotics Susceptibility Test
2.4. Synthesis of AgNPs
2.5. Preparation of Lincomycin Antibiotic Stock Solution
2.6. Characterization of Prepared AgNPs
2.7. Combined Effects of the Co-Delivered AgNPs–Lincomycin Mixture
2.8. Biofilm Formation Test
2.9. Evaluation of the MIC, and MBC
2.10. DPPH (1, 1-Diphenyl-2-picryl-hydrazyl) Assays
2.11. Growth Curve Tests
2.12. Analyses of Genes Expression Levels
2.12.1. Extraction of Total RNA
2.12.2. Reverse Transcription (RT)-PCR
2.12.3. Quantitative Real-Time PCR
2.13. Statistical Analyses
3. Results and Discussion
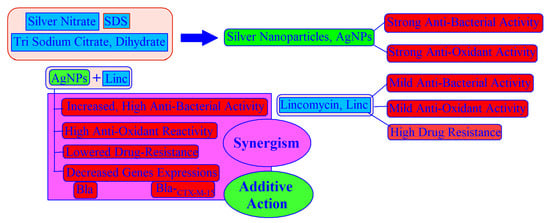
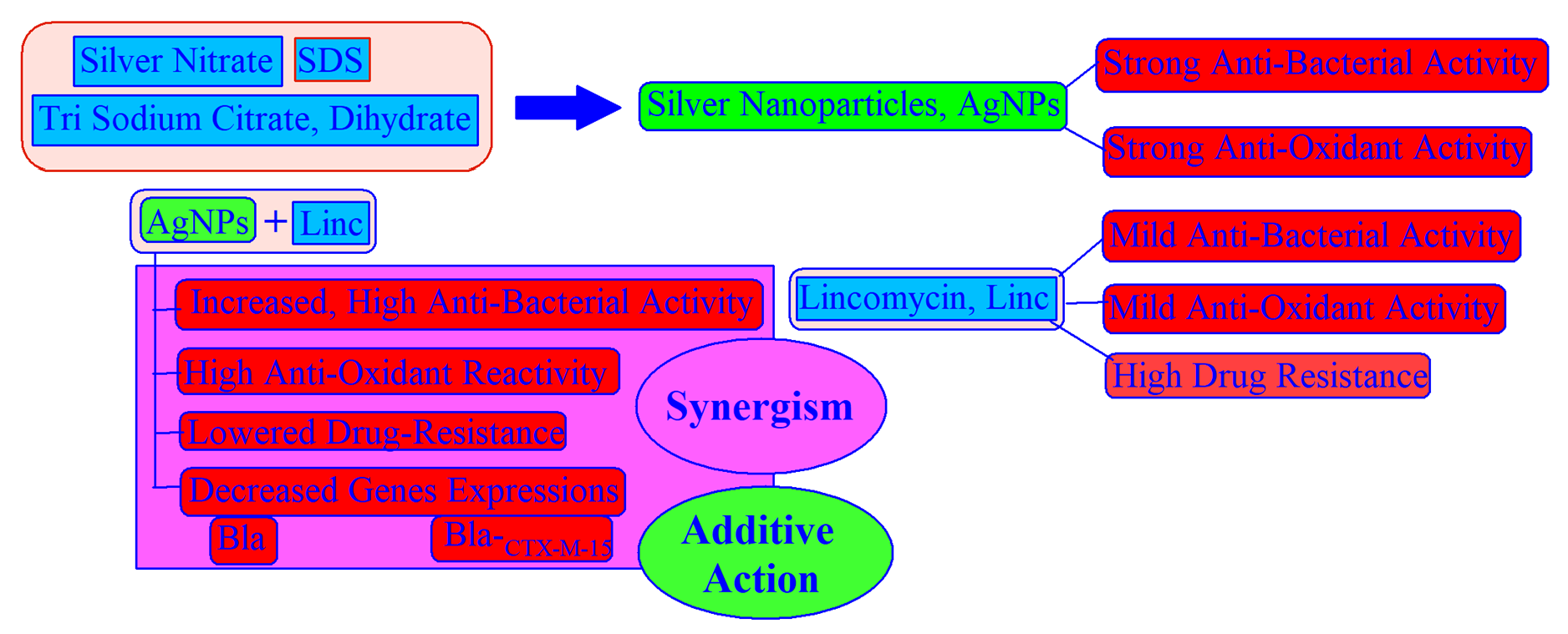
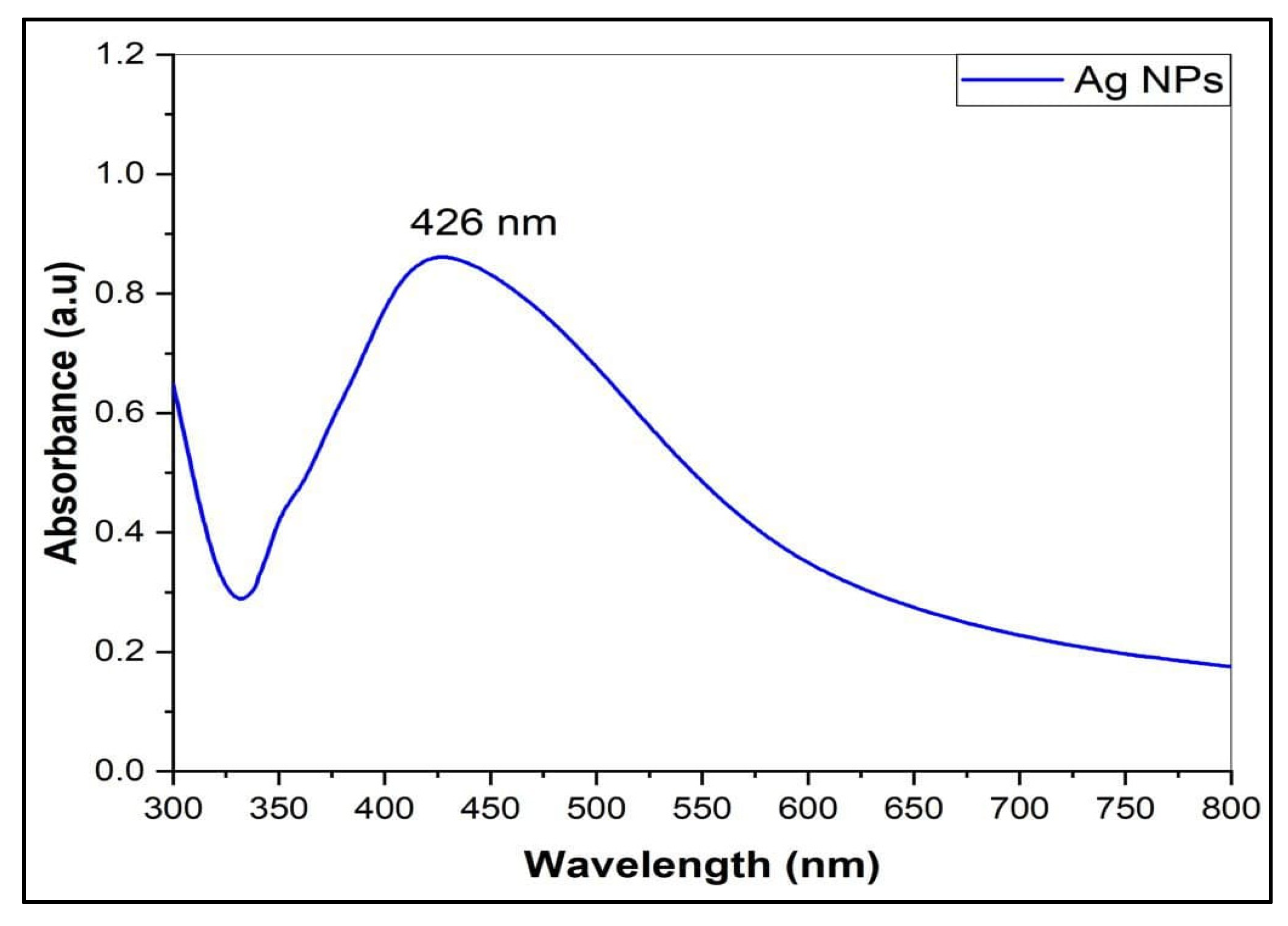
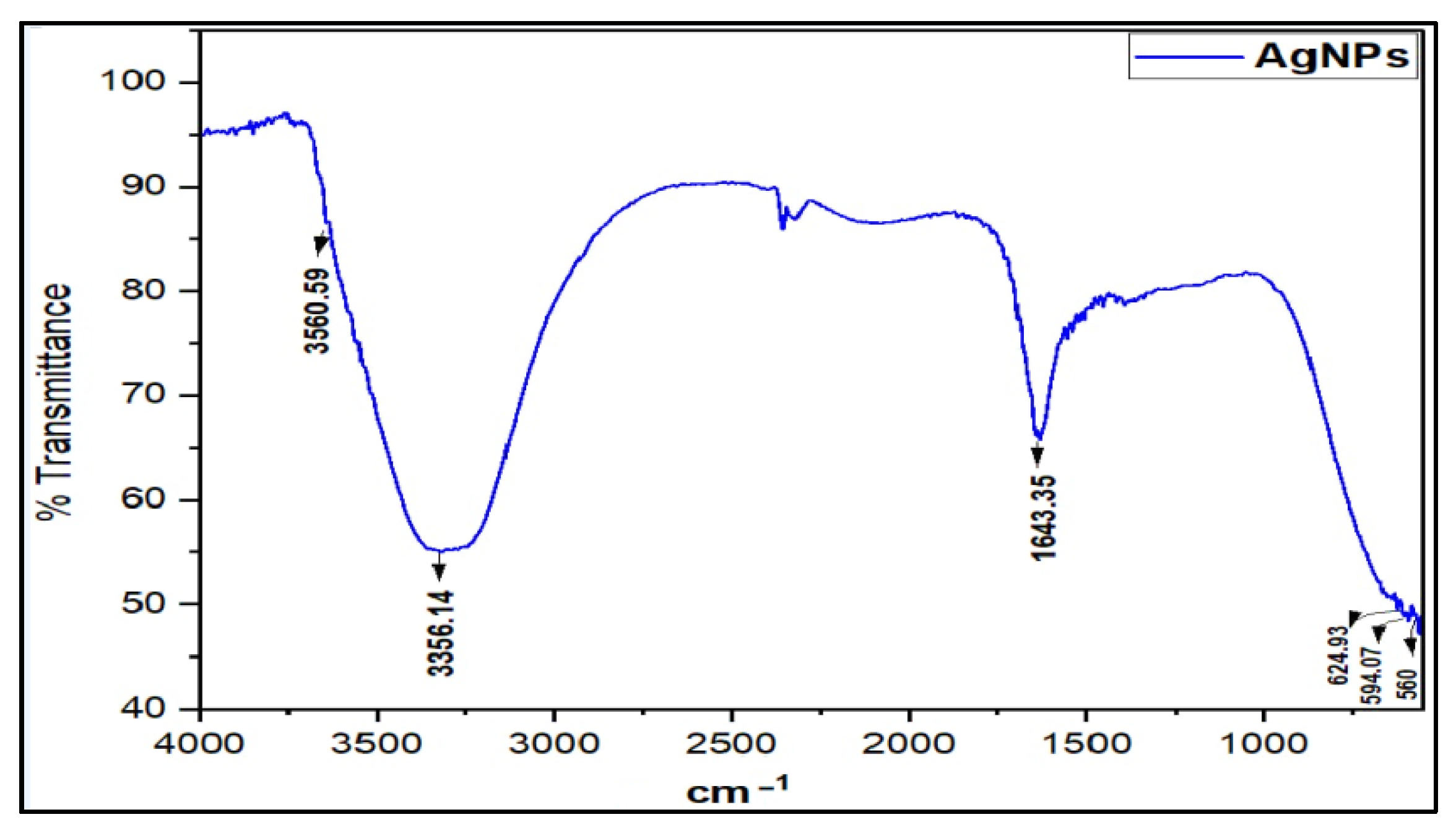
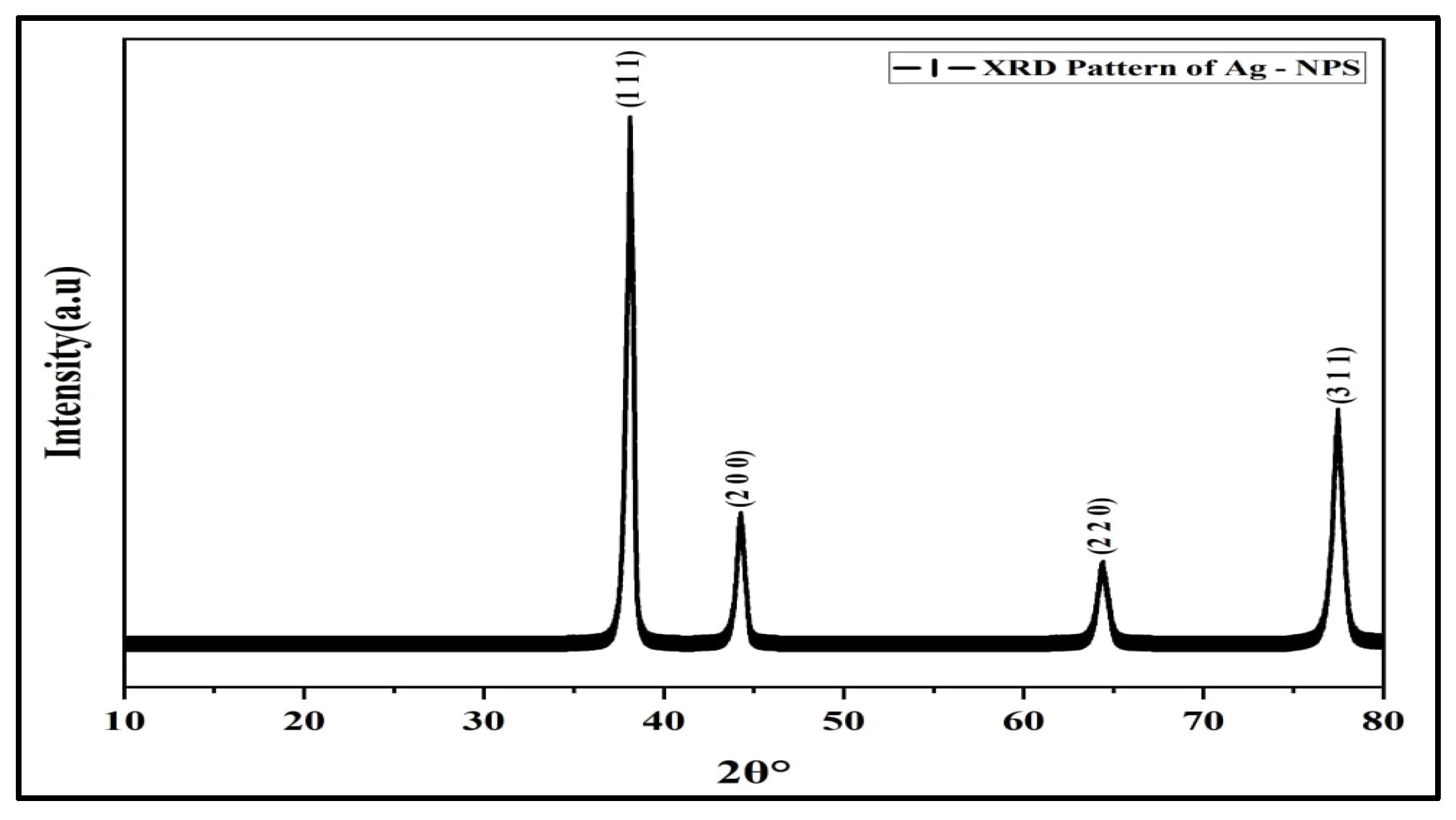
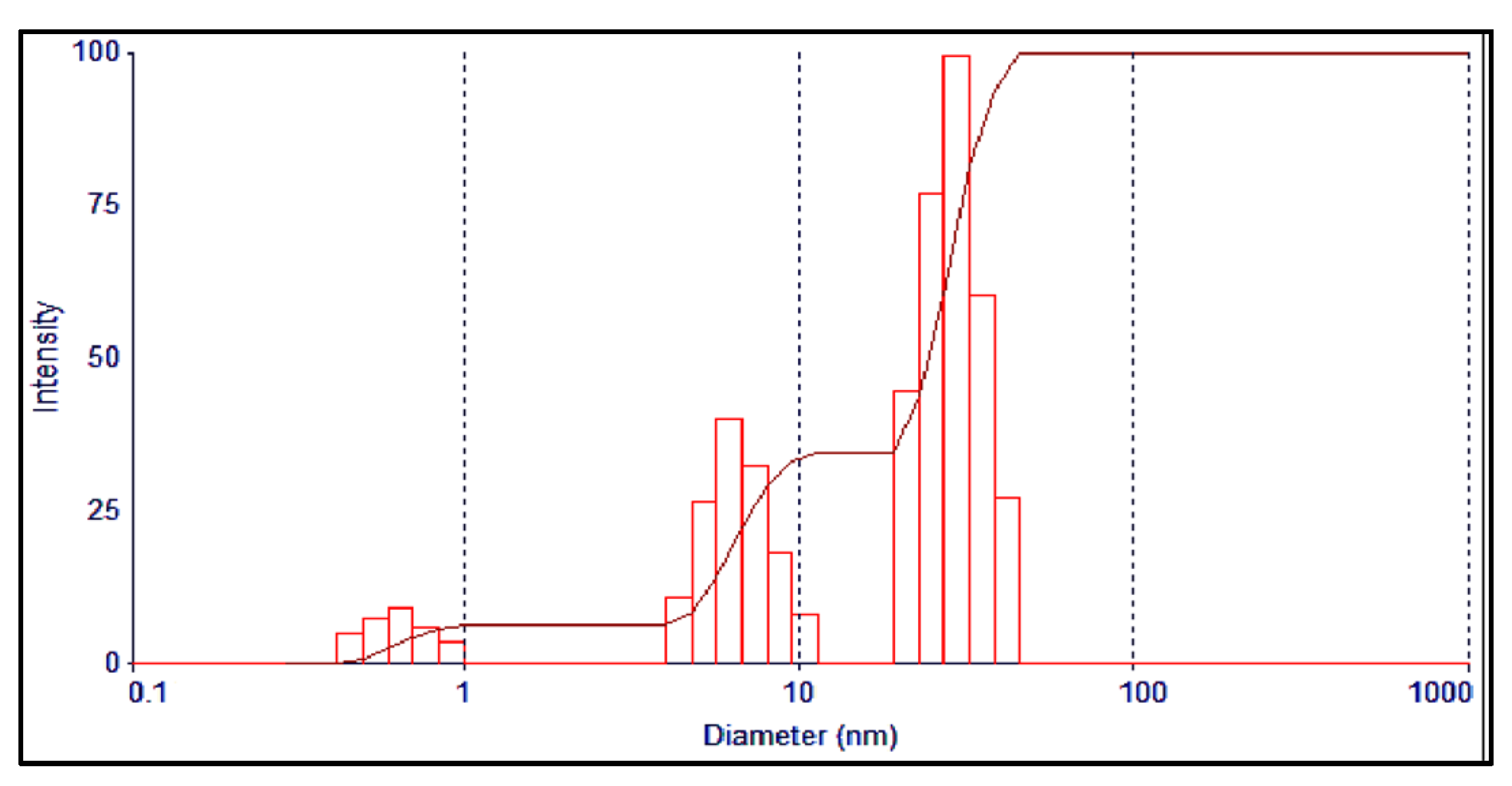
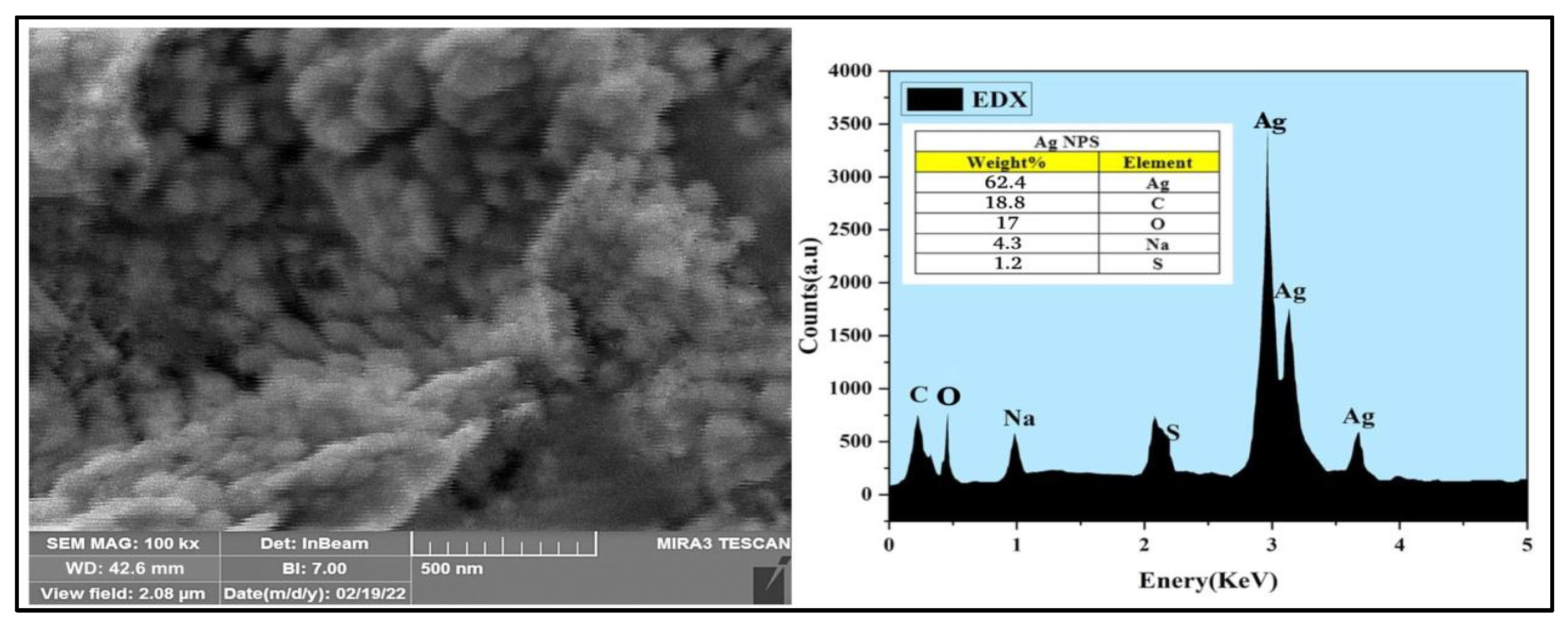
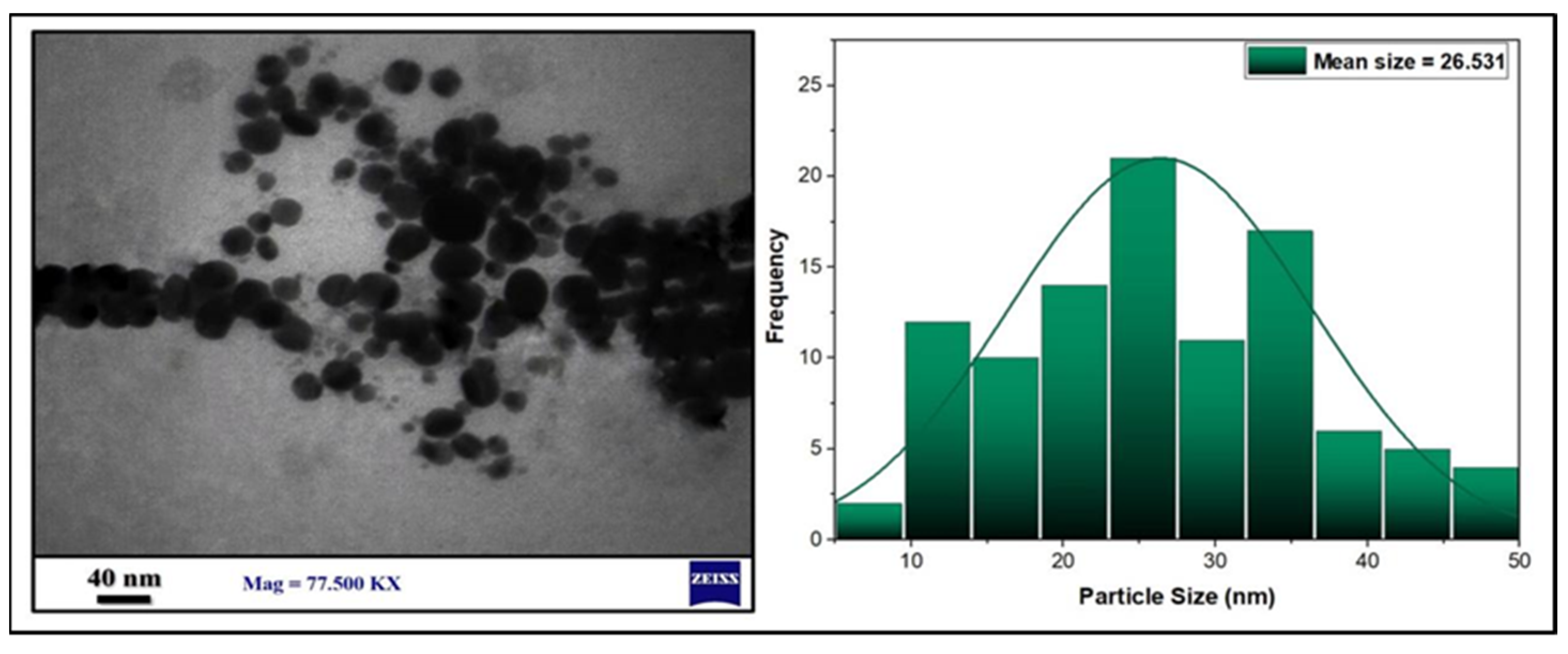
3.1. Preparation and Characterization of AgNPs
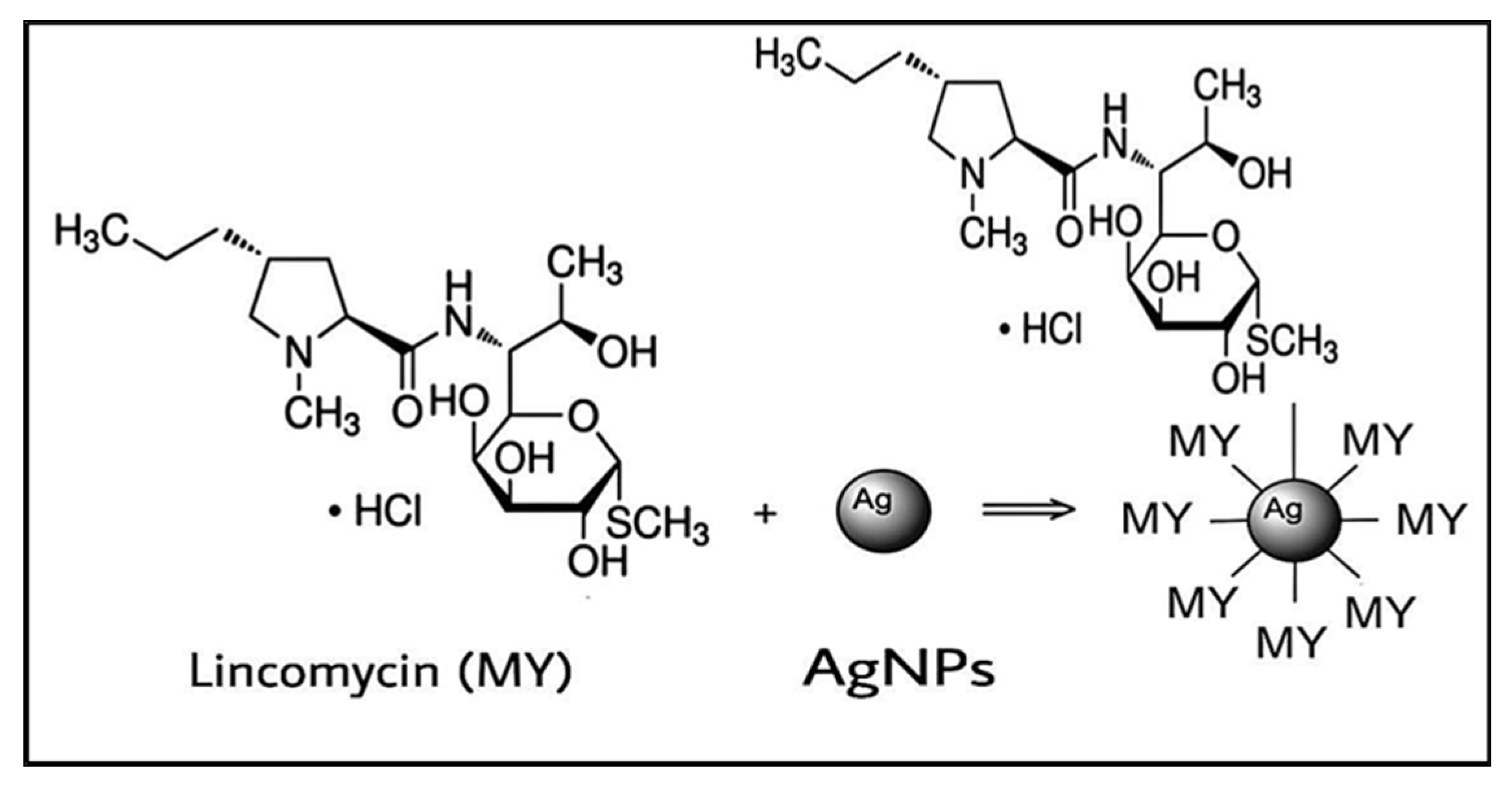
3.2. Plausible Mechanism of AgNPs Conjugation with Lincomycin
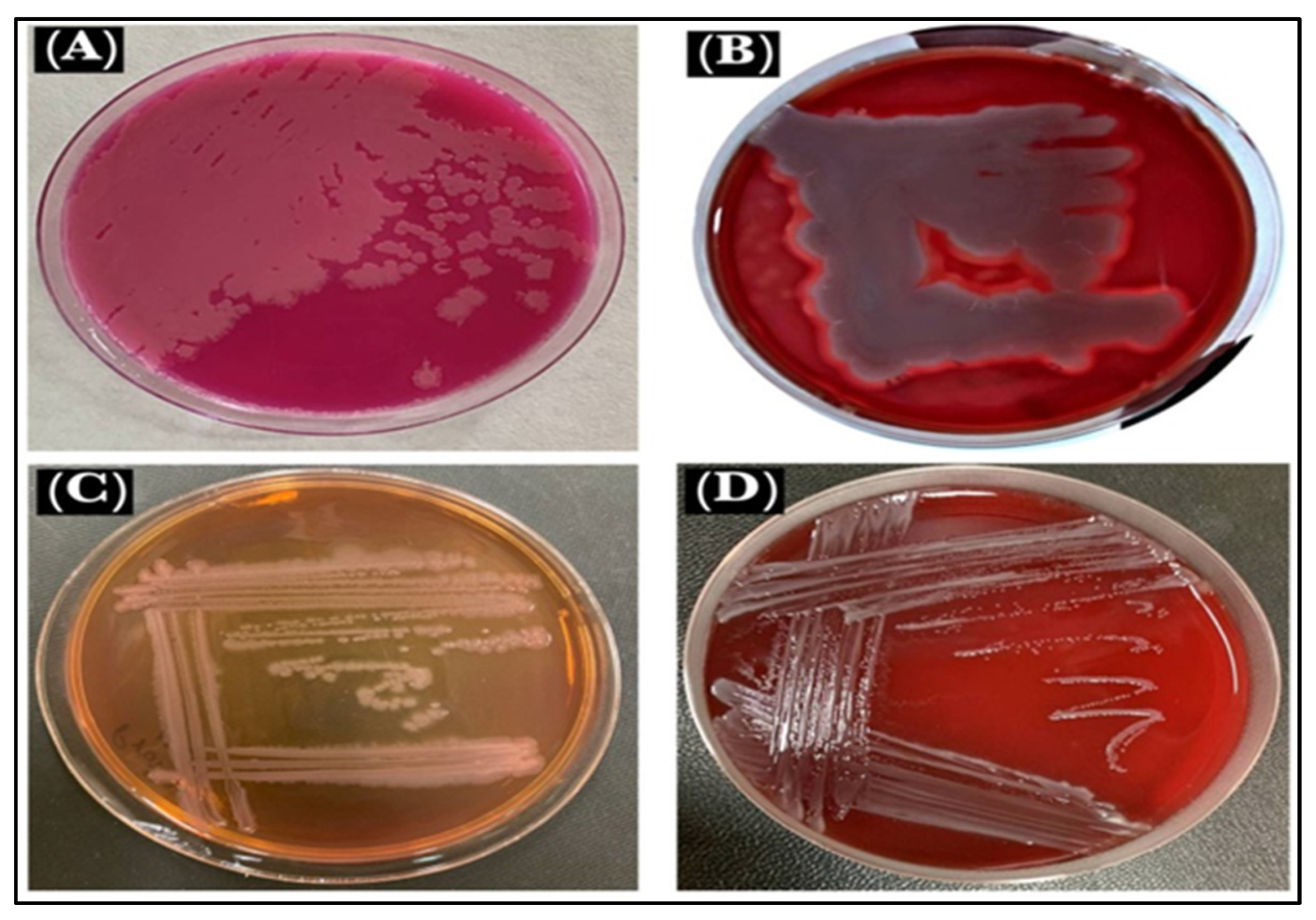
3.3. Culture Identifications
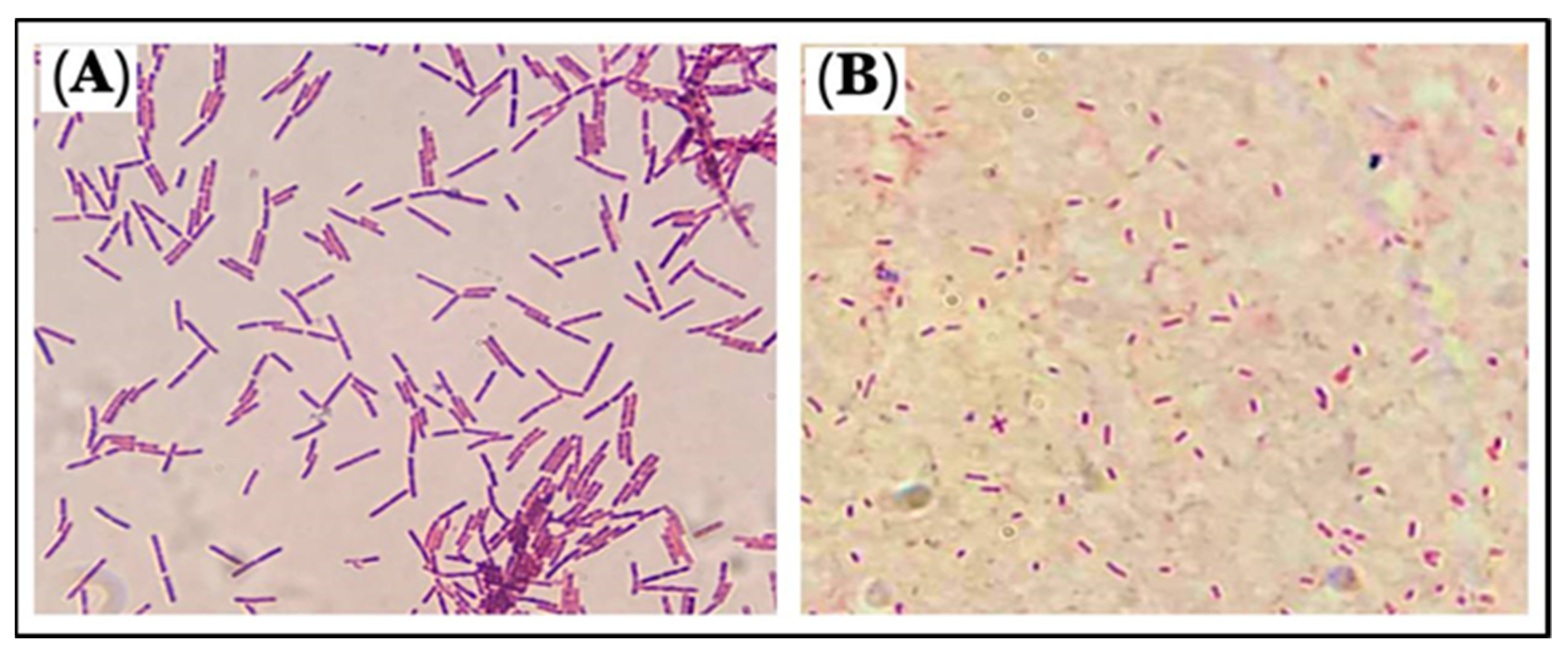
3.4. Microscopic Characterizations
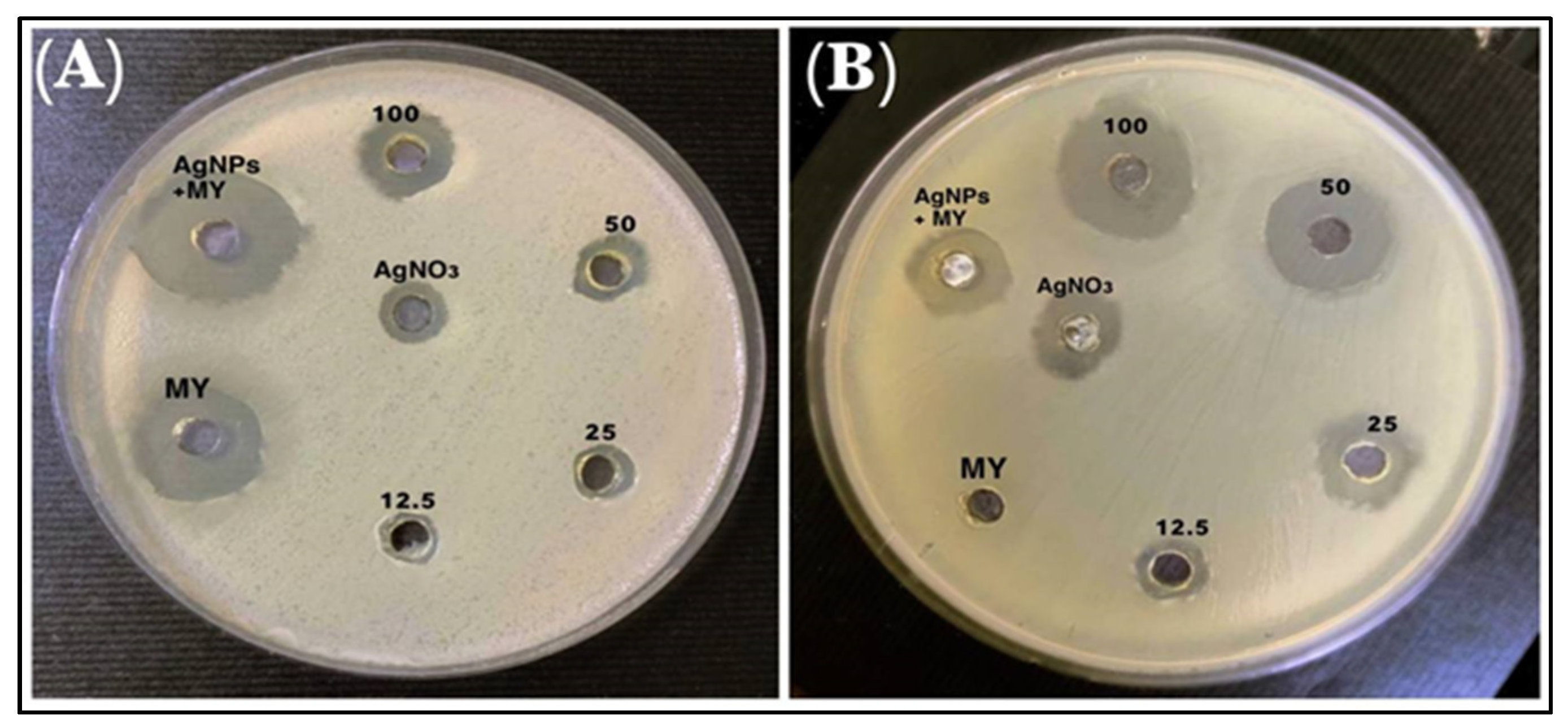
3.5. Antibiotic Susceptibility Tests
3.6. Biofilms Formations
3.7. Measurements of the MIC and MBC of AgNPs and AgNPs–Lincomycin Mixture
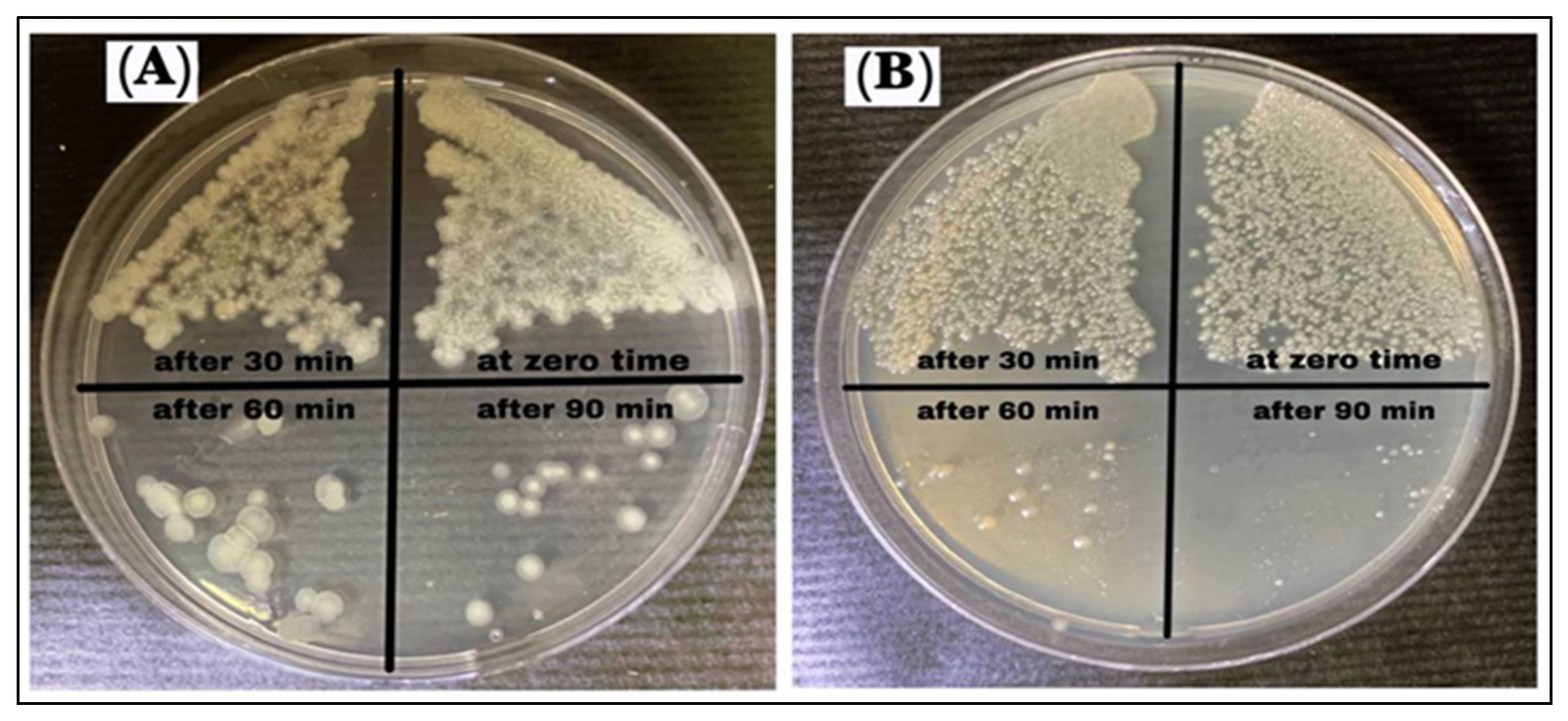
3.8. Growth Curve Test
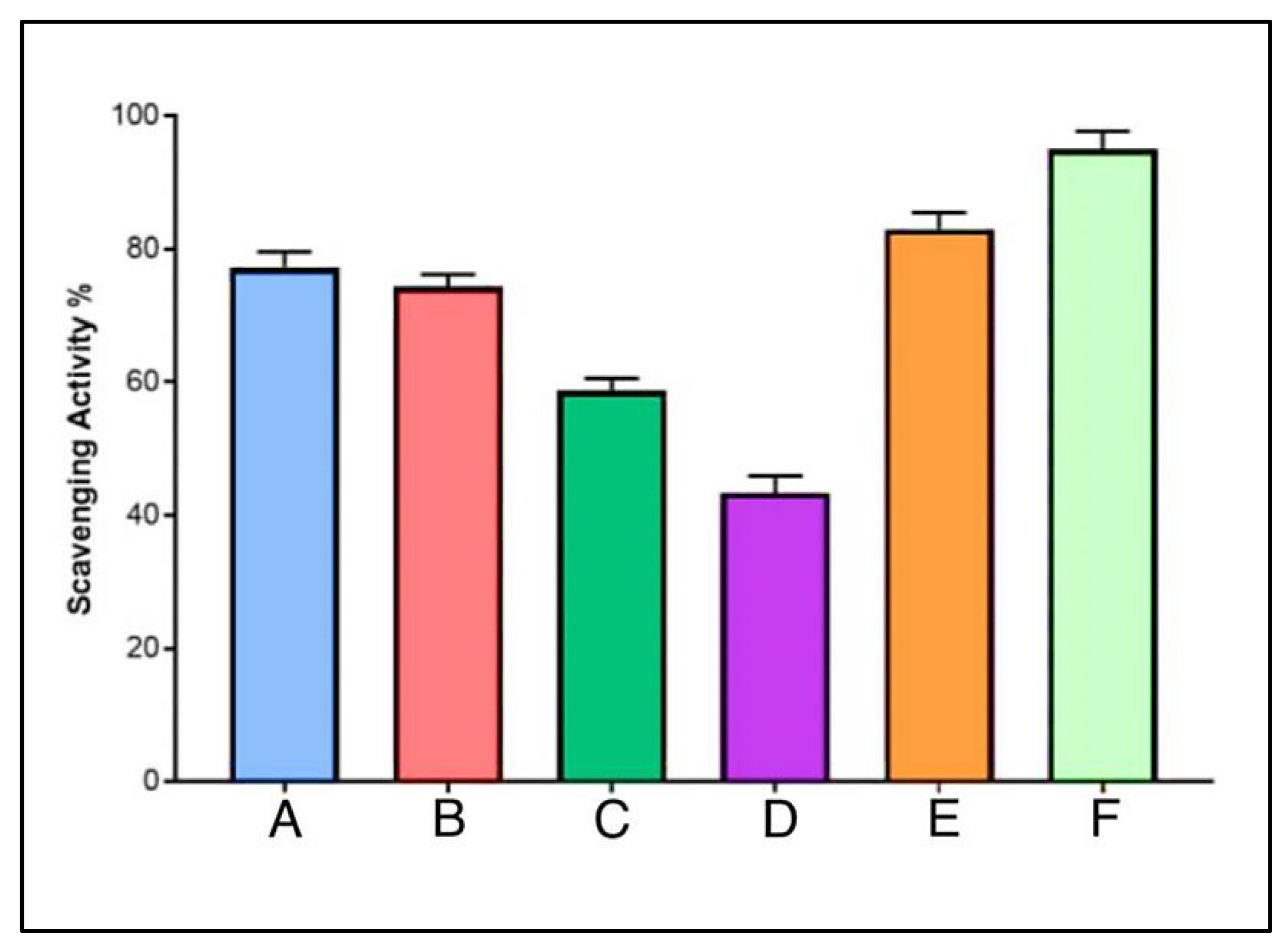
3.9. Antioxidant Potentials
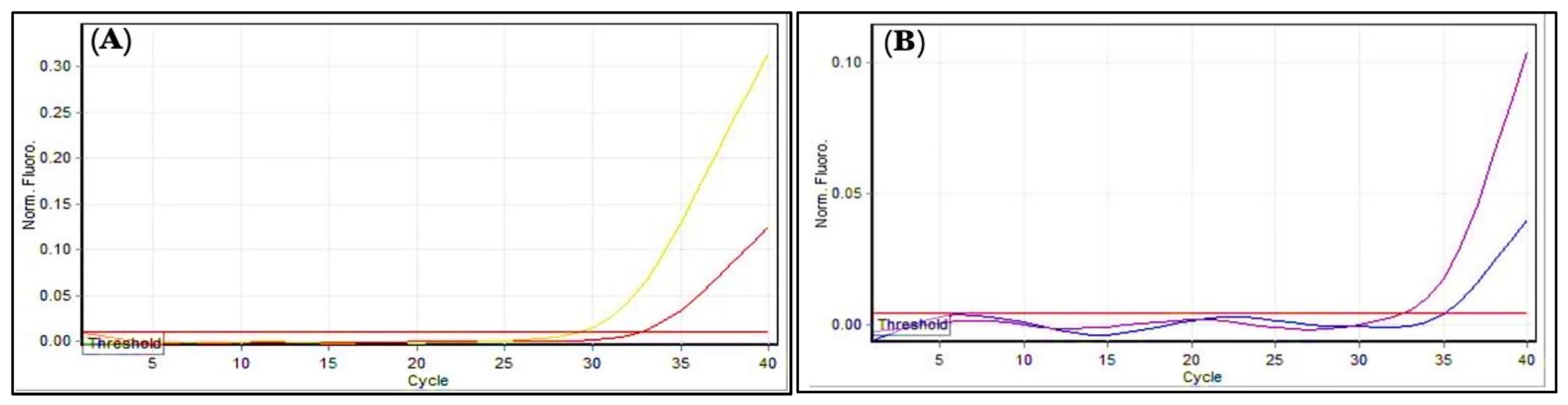
3.10. Genes Expression Levels
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. Global burden of bacterial anti-microbial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Ghazaei, C. Phenotypic and molecular detection of Beta-Lactamase enzyme produced by Bacillus cereus isolated from pasteurized and raw milk. J. Med. Bacteriol. 2019, 8, 1–7. [Google Scholar]
- El-Tarabili, R.M.; Ahmed, E.M.; Alharbi, N.K.; Alharbi, M.A.; AlRokban, A.H.; Naguib, D.; Alhag, S.K.; El Feky, T.M.; Ahmed, A.E.; Mahmoud, A.E. Prevalence, antibiotic profile, virulence determinants, ESBLs, and non-β-lactam encoding genes of MDR Proteus spp. isolated from infected dogs. Front. Genet. 2022, 13, 952689. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.J.; Woo, G.J. Distribution and transferability of tetracycline resistance determinants in Escherichia coli isolated from meat and meat products. Int. J. Food Microbiol. 2011, 145, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Nefedova, E.; Shkil, N.; Luna Vazquez-Gomez, R.; Garibo, D.; Pestryakov, A.; Bogdanchikova, N. AgNPs Targeting the Drug Resistance Problem of Staphylococcus aureus: Susceptibility to Antibiotics and Efflux Effect. Pharmaceutics 2022, 14, 763. [Google Scholar] [CrossRef] [PubMed]
- Ipe, D.S.; Kumar, P.T.S.; Love, R.M.; Hamlet, S.M. Silver nanoparticles at biocompatible dosage synergistically increases bacterial susceptibility to antibiotics. Front. Microbiol. 2020, 11, 1074. [Google Scholar] [CrossRef]
- Ibraheem, D.R.; Hussein, N.N.; Sulaiman, G.M.; Mohammed, H.A.; Khan, R.A.; Al Rugaie, O. Ciprofloxacin-Loaded Silver Nanoparticles as Potent Nano-Antibiotics against Resistant Pathogenic Bacteria. Nanomaterials 2022, 12, 2808. [Google Scholar] [CrossRef]
- Metryka, O.; Wasilkowski, D.; Mrozik, A. Evaluation of the Effects of Ag, Cu, ZnO and TiO2 Nanoparticles on the Expression Level of Oxidative Stress-Related Genes and the Activity of Antioxidant Enzymes in Escherichia coli, Bacillus cereus, and Staphylococcus epidermidis. Int. J. Mol. Sci. 2022, 23, 4966. [Google Scholar] [CrossRef]
- Mofidfar, M.; Kim, E.S.; Larkin, E.L.; Long, L.; Jennings, W.D.; Ahadian, S.; Ghannoum, M.A.; Wnek, G.E. Anti-microbial Activity of Silver Containing Crosslinked Poly(Acrylic Acid) Fibers. Micromachines 2019, 10, 829. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 3000540, Lincomycin. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Lincomycin (accessed on 22 November 2022).
- Finch, R.G.; Greenwood, D.; Whitley, R.J.; Norrby, S.R. Antibiotic and Chemotherapy; Elsevier Health Sciences: Philadelphia, PA, USA, 2010; ISBN 0702047651. [Google Scholar]
- Aronson, J.K. Meyler’s Side Effects of Anti-Microbial Drugs; Elsevier: Amsterdam, The Netherlands, 2009; ISBN 0080932932. [Google Scholar]
- Rezanka, T.; Spizek, J.; Sigler, K. Medicinal use of lincosamides and microbial resistance to them. Anti Infect. Agents Med. Chem. 2007, 6, 133–144. [Google Scholar] [CrossRef]
- Kharitonov, D.S.; Kasach, A.A.; Gibala, A.; Zimowska, M.; Kurilo, I.I.; Wrzesińska, A.; Szyk-Warszyńska, L.; Warszyński, P. Anodic Electrodeposition of Chitosan–AgNP Composites Using In Situ Coordination with Copper Ions. Materials 2021, 14, 2754. [Google Scholar] [CrossRef] [PubMed]
- Buzea, C.; Pacheco, I.I.; Robbie, K. Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases 2007, 2, MR17–MR71. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, R.; Hussein, N.N. Evaluation of the anti-bacterial activity of biosynthesis silver nanoparticles by lactobacillus gasseri bacteria. J. Appl. Sci. Nanotechnol. 2021, 1, 86–95. [Google Scholar] [CrossRef]
- Qureshi, K.A.; Bholay, A.D.; Rai, P.K.; Mohammed, H.A.; Khan, R.A.; Azam, F.; Jaremko, M.; Emwas, A.-H.; Stefanowicz, P.; Waliczek, M. Isolation, characterization, anti-MRSA evaluation, and in-silico multi-target anti-microbial validations of actinomycin X2 and actinomycin D produced by novel Streptomyces smyrnaeus UKAQ_23. Sci. Rep. 2021, 11, 14539. [Google Scholar] [CrossRef]
- Pinzaru, I.; Coricovac, D.; Dehelean, C.; Moacă, E.-A.; Mioc, M.; Baderca, F.; Sizemore, I.; Brittle, S.; Marti, D.; Calina, C.D. Stable PEG-coated silver nanoparticles–A comprehensive toxicological profile. Food Chem. Toxicol. 2018, 111, 546–556. [Google Scholar] [CrossRef]
- Dehghan, D.; Ramandi, M.F.; Taheri, R.A. Investigation of Synergism of Silver Nanoparticle and Erythromycin Inhibition and Detection of Exotoxin-A Gene in Pseudomonas aeruginosa Isolated from Burn Wounds Secretion. Iran. J. Med. Microbiol. 2020, 14, 379–389. [Google Scholar] [CrossRef]
- Hassan, A.; Usman, J.; Kaleem, F.; Omair, M.; Khalid, A.; Iqbal, M. Evaluation of different detection methods of biofilm formation in the clinical isolates. Braz. J. Infect. Dis. 2011, 15, 305–311. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Ali, H.M.; Qureshi, K.A.; Alsharidah, M.; Kandil, Y.I.; Said, R.; Mohammed, S.A.A.; Al-Omar, M.S.; Rugaie, O.A.; Abdellatif, A.A.H. Comparative Phytochemical Profile and Biological Activity of Four Major Medicinal Halophytes from Qassim Flora. Plants 2021, 10, 2208. [Google Scholar] [CrossRef]
- Abdulsada, F.M.; Hussein, N.N.; Sulaiman, G.M.; Al Ali, A.; Alhujaily, M. Evaluation of the Anti-bacterial Properties of Iron Oxide, Polyethylene Glycol, and Gentamicin Conjugated Nanoparticles against Some Multidrug-Resistant Bacteria. J. Funct. Biomater. 2022, 13, 138. [Google Scholar] [CrossRef]
- Chahardehi, A.M.; Ibrahim, D.; Sulaiman, S.F.; Mousavi, L. Time-kill study of ethyl acetate extract of stinging nettle on Bacillus subtilis subsp. spizizenii ATCC CRM-6633 Strain NRS 231. Annu. Res. Rev. Biol. 2015, 6, 33–40. [Google Scholar] [CrossRef]
- Mousa, D.; Abd El Tawab, A.; El Hofy, F.; Maarouf, A. Molecular studies on antibiotic-resistant Bacillus cereus isolated from meat products and humans in Kaliobia, Egypt. Benha Vet. Med. J. 2020, 38, 125–130. [Google Scholar] [CrossRef]
- Khan, U.; Afsana, S.; Kibtia, M.; Hossain, M.; Choudhury, N.; Ahsan, C.R. Presence of BlaCTX-M-15 antibiotic resistance gene in Lactobacillus spp. isolated from Hirschsprung-diseased infants with a stoma. J. Infect. Dev. Ctries. 2019, 13, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Shelenkov, A.; Petrova, L.; Fomina, V.; Zamyatin, M.; Mikhaylova, Y.; Akimkin, V. Multidrug-Resistant Proteus mirabilis Strain with Cointegrate Plasmid. Microorganisms 2020, 8, 1775. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, W.H.; Abdul Hassan, H.M.; Ali, M.A. Molecular and morphological study of the UV radiation on mice sperm formulation. EurAsian J. Biosci. 2020, 14, 4893–4900. [Google Scholar]
- Anandalakshmi, K.; Venugobal, J.; Ramasamy, V. Characterization of silver nanoparticles by green synthesis method using Pedalium murex leaf extract and their anti-bacterial activity. Appl. Nanosci. 2015, 6, 399–408. [Google Scholar] [CrossRef]
- Sudha, A.; Jeyakanthan, J.; Srinivasan, P. Green synthesis of silver nanoparticles using Lippia nodiflora aerial extract and evaluation of their antioxidant, anti-bacterial and cytotoxic effects. Resour. Effic. Technol. 2017, 3, 506–515. [Google Scholar] [CrossRef]
- Dudhagara, P.; Alagiya, J.; Bhagat, C.; Dudhagara, D.; Ghelani, A.; Desai, J.; Patel, R.; Vansia, A.; Nhiem, D.N.; Chen, Y.-Y.; et al. Biogenic Synthesis of Anti-bacterial, Hemo-compatible, and Anti-platelets Lysozyme Functionalized Silver Nanoparticles through the One-Step Process for Therapeutic Applications. Processes 2022, 10, 623. [Google Scholar] [CrossRef]
- XRD Crystallite (Grain) Size Calculator (Scherrer Equation)—InstaNANO. Available online: https://instanano.com/characterization/calculator/xrd/crystallite-size/ (accessed on 23 October 2022).
- Yerragopu, P.S.; Hiregoudar, S.; Nidoni, U.; Ramappa, K.T.; Sreenivas, A.G.; Doddagoudar, S.R. Chemical synthesis of silver nanoparticles using tri-sodium citrate, stability study, and their characterization. Int. Res. J. Pure Appl. Chem. 2020, 21, 37–50. [Google Scholar] [CrossRef]
- Izak-Nau, E.; Huk, A.; Reidy, B.; Uggerud, H.; Vadset, M.; Eiden, S.; Voetz, M.; Himly, M.; Duschl, A.; Dusinska, M. Impact of storage conditions and storage time on silver nanoparticles’ physicochemical properties and implications for their biological effects. RSC Adv. 2015, 5, 84172–84185. [Google Scholar] [CrossRef]
- Du, Q.; Jing, Z.; Qi, H.; Zuo, L.; Zhou, L.; He, H.; Sun, Z. Visual Recognition and Detection of Clindamycin by Au@Ag Core-Shell Nanoparticles. ACS Omega 2021, 6, 14260–14267. [Google Scholar] [CrossRef]
- Fuchs, E.; Raab, C.; Brugger, K.; Ehling-Schulz, M.; Wagner, M.; Stessl, B. Performance Testing of Bacillus cereus Chromogenic Agar Media for Improved Detection in Milk and Other Food Samples. Foods 2022, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Gottenbos, B.; Grijpma, D.W.; van der Mei, H.C.; Feijen, J.; Busscher, H.J. Anti-microbial effects of positively charged surfaces on adhering Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2001, 48, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Salvioni, L.; Galbiati, E.; Collico, V.; Alessio, G.; Avvakumova, S.; Corsi, F.; Tortora, P.; Prosperi, D.; Colombo, M. Negatively charged silver nanoparticles with potent anti-bacterial activity and reduced toxicity for pharmaceutical preparations. Int. J. Nanomed. 2017, 12, 2517. [Google Scholar] [CrossRef]
- Abbaszadegan, A.; Ghahramani, Y.; Gholami, A.; Hemmateenejad, B.; Dorostkar, S.; Nabavizadeh, M.; Sharghi, H. The effect of charge at the surface of silver nanoparticles on anti-microbial activity against Gram-positive and Gram-negative bacteria: A preliminary study. J. Nanomater. 2015, 2015, 720654. [Google Scholar] [CrossRef]
- Morar, M.; Bhullar, K.; Hughes, D.W.; Junop, M.; Wright, G.D. Structure and mechanism of the lincosamide antibiotic adenylyltransferase LinB. Structure 2009, 17, 1649–1659. [Google Scholar] [CrossRef]
- Leclercq, R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef]
- Siddique, M.H.; Aslam, B.; Imran, M.; Ashraf, A.; Nadeem, H.; Hayat, S.; Khurshid, M.; Afzal, M.; Malik, I.R.; Shahzad, M. Effect of silver nanoparticles on biofilm formation and EPS production of multidrug-resistant Klebsiella pneumoniae. BioMed Res. Int. 2020, 2020, 6398165. [Google Scholar] [CrossRef]
- Mikhailova, E.O. Silver nanoparticles: Mechanism of action and probable bio-application. J. Funct. Biomater. 2020, 11, 84. [Google Scholar] [CrossRef]
- Yang, B.; Lei, Z.; Zhao, Y.; Ahmed, S.; Wang, C.; Zhang, S.; Fu, S.; Cao, J.; Qiu, Y. Combination susceptibility testing of common anti-microbials in vitro and the effects of sub-MIC of anti-microbials on Staphylococcus aureus biofilm formation. Front. Microbiol. 2017, 8, 2125. [Google Scholar] [CrossRef]
- Mohsen, E.; El-Borady, O.M.; Mohamed, M.B.; Fahim, I.S. Synthesis and characterization of ciprofloxacin-loaded silver nanoparticles and investigation of their anti-bacterial effect. J. Radiat. Res. Appl. Sci. 2020, 13, 416–425. [Google Scholar] [CrossRef]
- Ferdous, Z.; Nemmar, A. Health Impact of Silver Nanoparticles: A Review of the Biodistribution and Toxicity Following Various Routes of Exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Mohammed, H.A.; Yusuf, M.; Al-Subaiyel, A.; Sulaiman, G.M.; Khan, R.A. SPIONs Conjugate Supported Anticancer Drug Doxorubicin’s Delivery: Current Status, Challenges, and Prospects. Nanomaterials 2022, 12, 3686. [Google Scholar] [CrossRef]
- Khan, B.; Nawaz, M.; Hussain, R.; Price, G.J.; Warsi, M.F.; Waseem, M. Enhanced anti-bacterial activity of size-controlled silver and polyethylene glycol functionalized silver nanoparticles. Chem. Pap. 2021, 75, 743–752. [Google Scholar] [CrossRef]
- de Allori, M.C.G.; Jure, M.A.; Romero, C.; de Castillo, M.E.C. Anti-microbial resistance and production of biofilms in clinical isolates of coagulase-negative Staphylococcus strains. Biol. Pharm. Bull. 2006, 29, 1592–1596. [Google Scholar] [CrossRef]
- Sharaf, S.M.A.; Abbas, H.S.; Ismaeil, T.A.M. Characterization of spirugenic iron oxide nanoparticles and their anti-bacterial activity against multidrug-resistant Helicobacter pylori. Egypt. J. Phycol. 2019, 1, 1–28. [Google Scholar]
- Kashmiri, Z.N.; Mankar, S.A. Free radicals and oxidative stress in bacteria. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 34–40. [Google Scholar]
- Deligiannakis, Y.; Sotiriou, G.A.; Pratsinis, S.E. Antioxidant and antiradical SiO2 nanoparticles covalently functionalized with gallic acid. ACS Appl. Mater. Interfaces 2012, 4, 6609–6617. [Google Scholar] [CrossRef]
- Gowin, E.; Świątek-Kościelna, B.; Mańkowski, P.; Januszkiewicz-Lewandowska, D. The Profile of Microorganisms Responsible for Port-Related Bacteremia in Pediatric Hemato-Oncological Patients. Cancer Control 2020, 27, 107327482090469. [Google Scholar] [CrossRef]
- Fontela, P.S.; O’Donnell, S.; Papenburg, J. Can biomarkers improve the rational use of antibiotics? Curr. Opin. Infect. Dis. 2018, 31, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Coburn, P.S.; Miller, F.C.; Enty, M.A.; Land, C.; LaGrow, A.L.; Mursalin, M.H.; Callegan, M.C. Expression of Bacillus cereus virulence-related genes in an ocular infection-related environment. Microorganisms 2020, 8, 607. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.L. The Molecular Basis for Beta-Lactamase Gene Expression in Bacillus anthracis, Bacillus cereus and Bacillus thuringiensis. Texas Medical Center Dissertations (via ProQuest). AAI3339607. 2008. Available online: https://digitalcommons.library.tmc.edu/dissertations/AAI3339607 (accessed on 22 November 2022).
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step 1 | Step 2 | Step 3 | |
|---|---|---|---|
| Temperature | 25 °C | 42 °C | 85 °C |
| Time | 10 min | 15 min | 5 s |
| Step | Temperatures (°C) | Time (s) | Cycles |
|---|---|---|---|
| Enzyme activation | 94 | 30 | 1 |
| Denaturation | 94 | 5 | 35 |
| Annealing | 52 | 15 | |
| Extension | 72 | 20 |
| Antibiotic | Concentration | Bacillus cereus | Proteus mirabilis | ||||
|---|---|---|---|---|---|---|---|
| µg/Disk | * Antibiotic | * Antibiotic + AgNPs | ** F. I.% | Antibiotic | Antibiotic + AgNPs | F. I.% | |
| AK | 10 | - | - | - | 6 | 24 | 300 |
| AMC-30 | 20/10 | 6 | 12 | 100 | 14 | 18 | 28.5 |
| CTX | 30 | - | - | - | 38 | 38 | - |
| E | 15 | 24 | 28 | 16.7 | - | - | - |
| MY | 15 | 24 | 34 | 41.6 | 6 | 13 | 116.7 |
| P | 10 IU | 6 | 12 | 100 | - | - | - |
| TE | 30 | 18 | 22 | 22.22 | - | - | - |
| STX | 25 | - | - | - | 24 | 24 | - |
| Microorganism | AgNPs, 100 μg/mL | AgNPs, 50 μg/mL | AgNPs, 25 μg/mL | AgNPs, 12.5 μg/mL | Lincomycin, 15 µg/mL | AgNPs–Lincomycin, (100 µg/mL + 15 µg/mL) |
|---|---|---|---|---|---|---|
| B. cereus | 17.50 ± 0.50 | 14.00 ± 1.00 | 11.50 ± 0.5 | 11.00 ± 0.5 | 24.33 ± 0.5 | 35.33 ± 0.50 |
| P. mirabilis | 26.00 ± 0.50 | 25.00 ± 0.50 | 17.33 ± 0.50 | 14.00 ± 0.50 | 6.00 ± 0.00 | 21.66 ± 1.00 |
| Bacteria | Conc. of AgNPs (μg/mL) | Conc. of AgNPs–Lincomycin (μg/mL) | ||
|---|---|---|---|---|
| MIC | MBC | MIC | MBC | |
| B. cereus | 50 | 100 | 12.5 | 25 |
| P. mirabilis | 50 | 100 | 25 | 50 |
| Bacteria | AgNPs (100 μg/mL) | |||
|---|---|---|---|---|
| Zero | 30 min | 60 min | 90 min | |
| B. cereus | ++++ | +++ | ++ | + |
| P. mirabilis | ++++ | +++ | ++ | + |
| Groups | Mean Ct, Bla | Mean Ct, HKG | ΔCt (Mean Ct, Bla—Mean Ct, HKG) | 2−ΔCt | Experimental Group/Control Group | Fold(s) of Gene Expression |
|---|---|---|---|---|---|---|
| Group # 1 (Control) | 29.2 | 22.1 | 7.1000 | 0.0073 | 0.0073/0.0073 | 1.0000 |
| Group # 2 (After AgNPs treatment) | 32.65 | 22.1 | 10.5500 | 0.0007 | 0.0007/0.0073 | 0.0915 |
| Groups | Mean Ct, BlaCTX-M-15 | Mean Ct, HKG | ΔCt (Mean Ct, BlaCTX-M-15—Mean Ct, HKG) | 2−ΔCt | Experimental Group/Control Group | Fold of Gene Expression |
|---|---|---|---|---|---|---|
| Group # 1 (Control) | 32.58 | 18.9 | 13.6800 | 0.00008 | 0.00008/0.00008 | 1.0000 |
| Group # 2 (After AgNPs treatment) | 35.08 | 18.9 | 16.1800 | 0.000013 | 0.000013/0.00008 | 0.1768 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul-Jabbar, A.M.; Hussian, N.N.; Mohammed, H.A.; Aljarbou, A.; Akhtar, N.; Khan, R.A. Combined Anti-Bacterial Actions of Lincomycin and Freshly Prepared Silver Nanoparticles: Overcoming the Resistance to Antibiotics and Enhancement of the Bioactivity. Antibiotics 2022, 11, 1791. https://doi.org/10.3390/antibiotics11121791
Abdul-Jabbar AM, Hussian NN, Mohammed HA, Aljarbou A, Akhtar N, Khan RA. Combined Anti-Bacterial Actions of Lincomycin and Freshly Prepared Silver Nanoparticles: Overcoming the Resistance to Antibiotics and Enhancement of the Bioactivity. Antibiotics. 2022; 11(12):1791. https://doi.org/10.3390/antibiotics11121791
Chicago/Turabian StyleAbdul-Jabbar, Amna M., Nehia N. Hussian, Hamdoon A. Mohammed, Ahmed Aljarbou, Naseem Akhtar, and Riaz A. Khan. 2022. "Combined Anti-Bacterial Actions of Lincomycin and Freshly Prepared Silver Nanoparticles: Overcoming the Resistance to Antibiotics and Enhancement of the Bioactivity" Antibiotics 11, no. 12: 1791. https://doi.org/10.3390/antibiotics11121791
APA StyleAbdul-Jabbar, A. M., Hussian, N. N., Mohammed, H. A., Aljarbou, A., Akhtar, N., & Khan, R. A. (2022). Combined Anti-Bacterial Actions of Lincomycin and Freshly Prepared Silver Nanoparticles: Overcoming the Resistance to Antibiotics and Enhancement of the Bioactivity. Antibiotics, 11(12), 1791. https://doi.org/10.3390/antibiotics11121791