
Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru
 , , , and
, , , and
Abstract
1. Introduction
2. Methods
2.1. Sample Collection and Bacterial Culture
2.2. mcr-1 Gene Screening by PCR
2.3. Whole-Genome Sequencing and Assembly
2.4. Sequence Analysis
3. Results
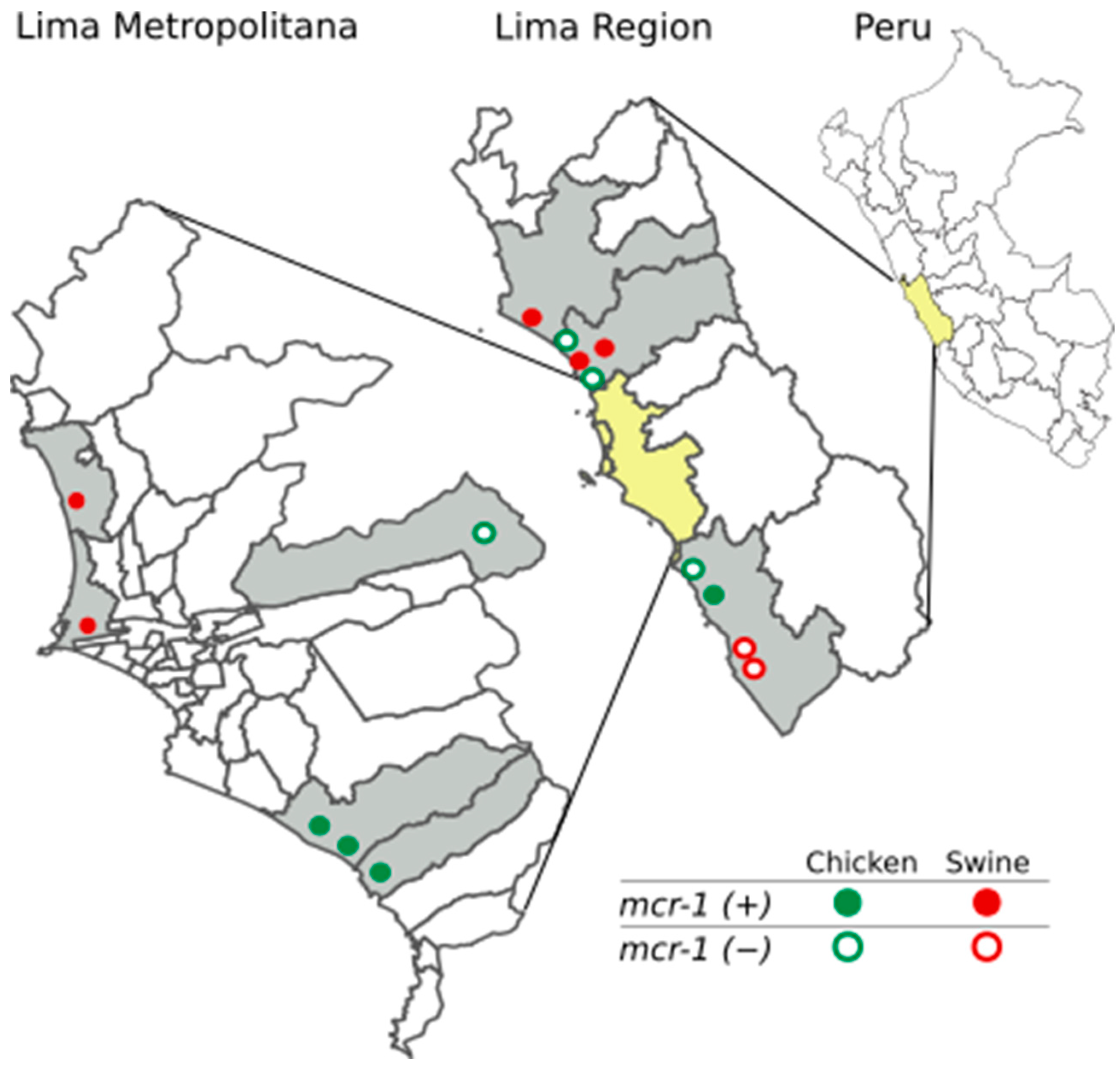
3.1. Prevalence of the mcr-1 Gene in Poultry and Pig Farms
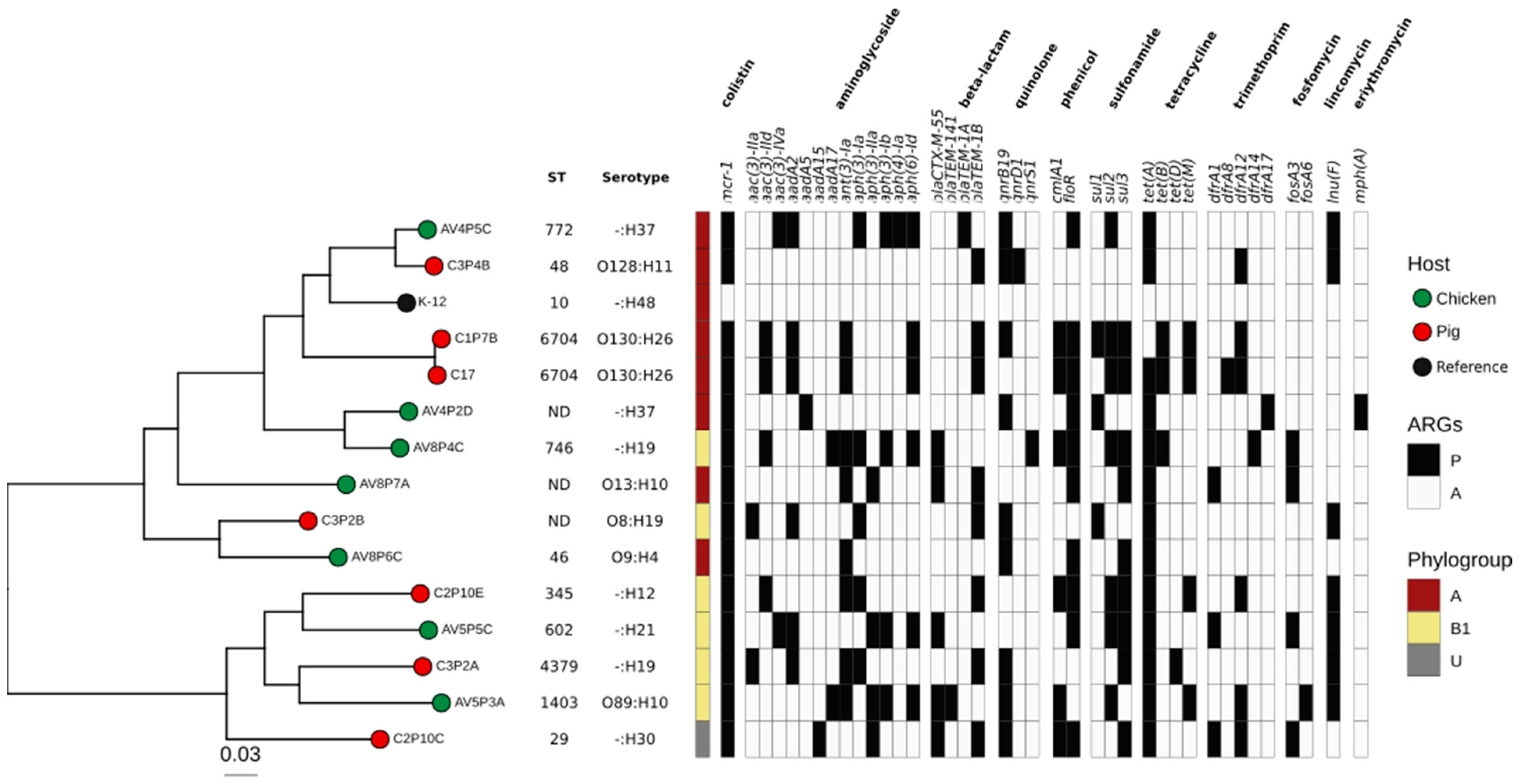
3.2. Genetic Characterization of E. coli Harboring mcr-1 and Resistome
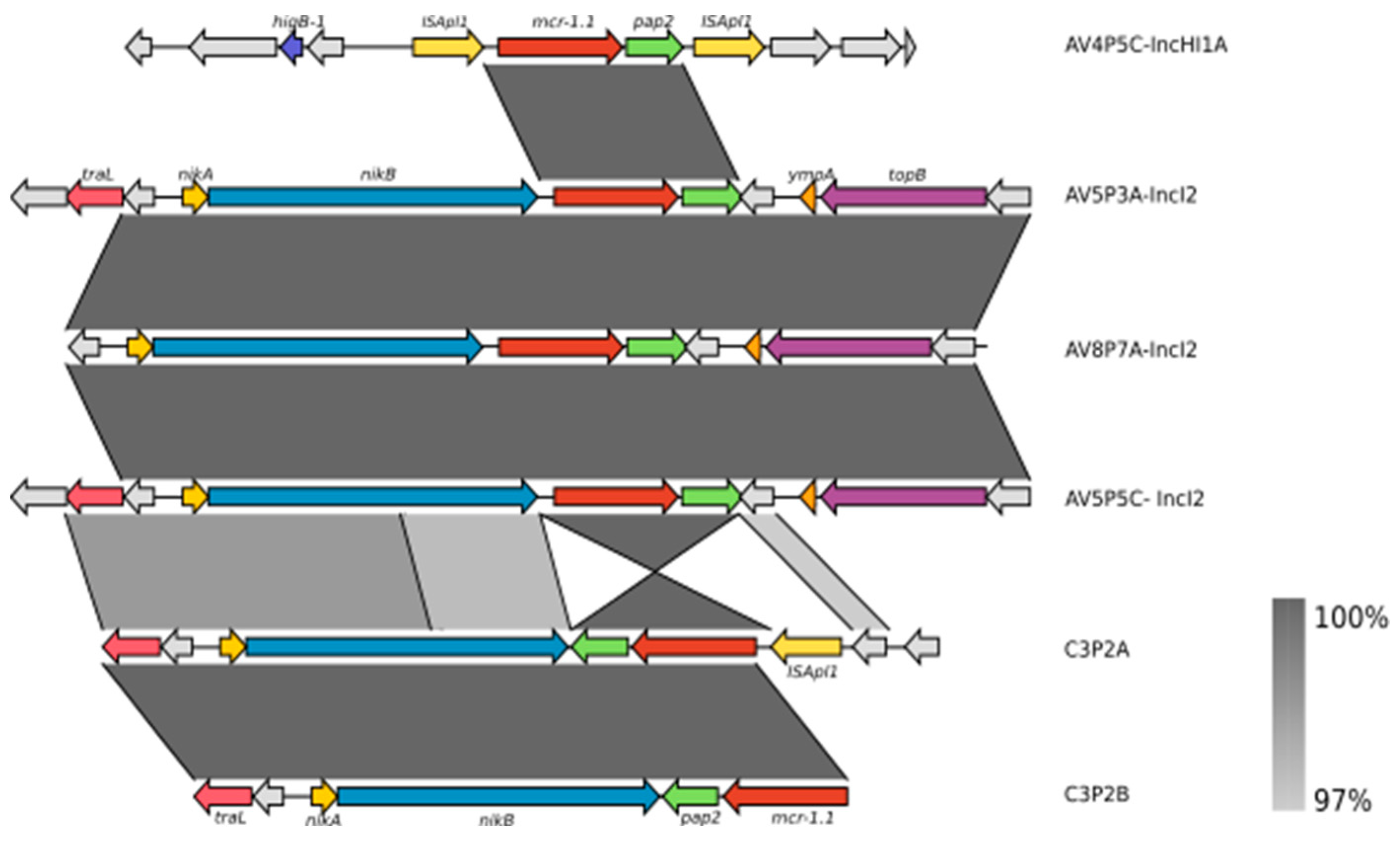
3.3. Characterization of the Genetic Context of the mcr-1 Gene
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sulis, G.; Gandra, S. Access to Antibiotics: Not a Problem in Some LMICs. Lancet Glob. Health 2021, 9, e561–e562. [Google Scholar] [CrossRef] [PubMed]
- WHO Critically Important Antimicrobials for Human Medicine: 6th Revision. Available online: https://www.who.int/publications-detail-redirect/9789241515528 (accessed on 20 October 2022).
- Kempf, I.; Jouy, E.; Chauvin, C. Colistin Use and Colistin Resistance in Bacteria from Animals. Int. J. Antimicrob. Agents 2016, 48, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global Phylogenetic Analysis of Escherichia coli and Plasmids Carrying the mcr-1 Gene Indicates Bacterial Diversity but Plasmid Restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The Global Distribution and Spread of the Mobilized Colistin Resistance Gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6, 4. [Google Scholar] [CrossRef]
- Leekitcharoenphon, P.; Johansson, M.H.K.; Munk, P.; Malorny, B.; Skarżyńska, M.; Wadepohl, K.; Moyano, G.; Hesp, A.; Veldman, K.T.; Bossers, A.; et al. Genomic Evolution of Antimicrobial Resistance in Escherichia coli. Sci. Rep. 2021, 11, 15108. [Google Scholar] [CrossRef]
- Brisola, M.C.; Crecencio, R.B.; Bitner, D.S.; Frigo, A.; Rampazzo, L.; Stefani, L.M.; Faria, G.A. Escherichia coli Used as a Biomarker of Antimicrobial Resistance in Pig Farms of Southern Brazil. Sci. Total Environ. 2019, 647, 362–368. [Google Scholar] [CrossRef]
- Vu Thi Ngoc, B.; Le Viet, T.; Nguyen Thi Tuyet, M.; Nguyen Thi Hong, T.; Nguyen Thi Ngoc, D.; Le Van, D.; Chu Thi, L.; Tran Huy, H.; Penders, J.; Wertheim, H.; et al. Characterization of Genetic Elements Carrying mcr-1 Gene in Escherichia coli from the Community and Hospital Settings in Vietnam. Microbiol. Spectr. 2022, 10, e0135621. [Google Scholar] [CrossRef]
- Yang, H.; Chen, S.; White, D.G.; Zhao, S.; McDermott, P.; Walker, R.; Meng, J. Characterization of Multiple-Antimicrobial-Resistant Escherichia coli Isolates from Diseased Chickens and Swine in China. J. Clin. Microbiol. 2004, 42, 3483–3489. [Google Scholar] [CrossRef]
- Ugarte Silva, R.G.; Olivo López, J.M.; Corso, A.; Pasteran, F.; Albornoz, E.; Sahuanay Blácido, Z.P. Resistencia a Colistín Mediado por el gen mcr-1 Identificado en Cepas de Escherichia coli y Klebsiella pneumoniae: Primeros Reportes en el Perú. In Anales de la Facultad de Medicina; UNMSM: Lima, Peru, 2018; Volume 79, pp. 213–217. [Google Scholar] [CrossRef][Green Version]
- Deshpande, L.M.; Hubler, C.; Davis, A.P.; Castanheira, M. Updated Prevalence of mcr-like Genes among Escherichia coli and Klebsiella pneumoniae in the SENTRY Program and Characterization of mcr-1.11 Variant. Antimicrob. Agents Chemother. 2019, 63, e02450-18. [Google Scholar] [CrossRef] [PubMed]
- Yauri-Condor, K.; Zavaleta Apestegui, M.; Sevilla-Andrade, C.R.; Sara, J.P.; Villoslado Espinoza, C.; Taboada, W.V.; Gonzales-Escalante, E. Extended-Spectrum Beta-Lactamase-Producing Enterobacterales Carrying the mcr-1 Gene in Lima, Peru. Rev. Peru. Med. Exp. Salud Pública 2021, 37, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Escalante, E.; Ruggiero, M.; Cerdeira, L.; Esposito, F.; Fontana, H.; Lincopan, N.; Gutkind, G.; Di Conza, J. Whole-Genome Analysis of a High-Risk Clone of Klebsiella pneumoniae ST147 Carrying Both mcr-1 and blaNDM-1 Genes in Peru. Microb. Drug Resist. 2022, 28, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Salvatierra, G.; Dávila-Barclay, A.; Ayzanoa, B.; Castillo-Vilcahuaman, C.; Huang, M.; Pajuelo, M.J.; Lescano, A.G.; Cabrera, L.; Calderón, M.; et al. Market Chickens as a Source of Antibiotic-Resistant Escherichia coli in a Peri-Urban Community in Lima, Peru. Front. Microbiol. 2021, 12, 635871. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global Trends in Antimicrobial Resistance in Animals in Low- and Middle-Income Countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for Detection of Plasmid-Mediated Colistin Resistance Determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for Surveillance Purposes. Euro Surveill. Bull. Eur. Sur Mal. Transm. Eur. Commun. Dis. Bull. 2018, 23, 17-00672. [Google Scholar] [CrossRef]
- Andrews, S. FastQC. A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 14 July 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinform. Oxf. Engl. 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Waters, N.R.; Abram, F.; Brennan, F.; Holmes, A.; Pritchard, L. Easy Phylotyping of Escherichia coli via the EzClermont Web App and Command-Line Tool. Access Microbiol. 2020, 2, acmi000143. [Google Scholar] [CrossRef]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid Phylogenetic Analysis of Large Samples of Recombinant Bacterial Whole Genome Sequences Using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. Ggtree: An r Package for Visualization and Annotation of Phylogenetic Trees with Their Covariates and Other Associated Data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. PlasmidSPAdes: Assembling Plasmids from Whole Genome Sequencing Data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef]
- Carattoli, A.; Hasman, H. PlasmidFinder and In Silico PMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). In Horizontal Gene Transfer: Methods and Protocols; de la Cruz, F., Ed.; Methods in Molecular Biology; Springer US: New York, NY, USA, 2020; pp. 285–294. ISBN 978-1-4939-9877-7. [Google Scholar]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.-Y. OriTfinder: A Web-Based Tool for the Identification of Origin of Transfers in DNA Sequences of Bacterial Mobile Genetic Elements. Nucleic Acids Res. 2018, 46, W229–W234. [Google Scholar] [CrossRef]
- Schmartz, G.P.; Hartung, A.; Hirsch, P.; Kern, F.; Fehlmann, T.; Müller, R.; Keller, A. PLSDB: Advancing a Comprehensive Database of Bacterial Plasmids. Nucleic Acids Res. 2022, 50, D273–D278. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple Prokaryote Genome Comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Barbieri, N.L.; Pimenta, R.L.; de Melo, D.A.; Nolan, L.K.; de Souza, M.M.S.; Logue, C.M. mcr-1 Identified in Fecal Escherichia coli and Avian Pathogenic E. coli (APEC) From Brazil. Front. Microbiol. 2021, 12, 799. [Google Scholar] [CrossRef]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence Factors, Prevalence and Potential Transmission of Extraintestinal Pathogenic Escherichia coli Isolated from Different Sources: Recent Reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef]
- Snesrud, E.; McGann, P.; Chandler, M. The Birth and Demise of the ISApl1-mcr-1-ISApl1 Composite Transposon: The Vehicle for Transferable Colistin Resistance. mBio 2018, 9, e02381-17. [Google Scholar] [CrossRef] [PubMed]
- El Peruano. Resolucion Directoral-No 0091-2019-MINAGRI-SENASA-DIAIA. Disponen Prohibir la Importación, Comercialización, Fabricación o Elaboración de Productos Veterinarios que Contengan el Principio Activo Colistina (Polimixina E) o Cualquiera de sus Sales y Dictan Diversas Disposiciones. Available online: http://busquedas.elperuano.pe/normaslegales/disponen-prohibir-la-importacion-comercializacion-fabricac-resolucion-directoral-no-0091-2019-minagri-senasa-diaia-1832393-1/ (accessed on 29 April 2022).
- Shen, C.; Zhong, L.-L.; Yang, Y.; Doi, Y.; Paterson, D.L.; Stoesser, N.; Ma, F.; Ahmed, M.A.E.-G.E.-S.; Feng, S.; Huang, S.; et al. Dynamics of mcr-1 Prevalence and mcr-1-Positive Escherichia coli after the Cessation of Colistin Use as a Feed Additive for Animals in China: A Prospective Cross-Sectional and Whole Genome Sequencing-Based Molecular Epidemiological Study. Lancet Microbe 2020, 1, e34–e43. [Google Scholar] [CrossRef] [PubMed]
- Mendes Oliveira, V.R.; Paiva, M.C.; Lima, W.G. Plasmid-Mediated Colistin Resistance in Latin America and Caribbean: A Systematic Review. Travel Med. Infect. Dis. 2019, 31, 101459. [Google Scholar] [CrossRef] [PubMed]
- Duriez, P.; Clermont, O.; Bonacorsi, S.; Bingen, E.; Chaventré, A.; Elion, J.; Picard, B.; Denamur, E. Commensal Escherichia coli Isolates Are Phylogenetically Distributed among Geographically Distinct Human Populations. Microbiology 2001, 147, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Madoshi, B.P.; Kudirkiene, E.; Mtambo, M.M.A.; Muhairwa, A.P.; Lupindu, A.M.; Olsen, J.E. Characterisation of Commensal Escherichia coli Isolated from Apparently Healthy Cattle and Their Attendants in Tanzania. PLoS ONE 2016, 11, e0168160. [Google Scholar] [CrossRef]
- Fuentes-Castillo, D.; Esposito, F.; Cardoso, B.; Dalazen, G.; Moura, Q.; Fuga, B.; Fontana, H.; Cerdeira, L.; Dropa, M.; Rottmann, J.; et al. Genomic Data Reveal International Lineages of Critical Priority Escherichia coli Harbouring Wide Resistome in Andean Condors (Vultur Gryphus Linnaeus, 1758). Mol. Ecol. 2020, 29, 1919–1935. [Google Scholar] [CrossRef]
- Aworh, M.K.; Abiodun-Adewusi, O.; Mba, N.; Helwigh, B.; Hendriksen, R.S. Prevalence and Risk Factors for Faecal Carriage of Multidrug Resistant Escherichia coli among Slaughterhouse Workers. Sci. Rep. 2021, 11, 13362. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Monte, D.F.; Mem, A.; Fernandes, M.R.; Cerdeira, L.; Esposito, F.; Galvão, J.A.; Franco, B.D.G.M.; Lincopan, N.; Landgraf, M. Chicken Meat as a Reservoir of Colistin-Resistant Escherichia coli Strains Carrying mcr-1 Genes in South America. Antimicrob. Agents Chemother. 2017, 61, e02718-16. [Google Scholar] [CrossRef]
- Faccone, D.; Moredo, F.A.; Giacoboni, G.I.; Albornoz, E.; Alarcón, L.; Nievas, V.F.; Corso, A. Multidrug-Resistant Escherichia coli Harbouring mcr-1 and blaCTX-M Genes Isolated from Swine in Argentina. J. Glob. Antimicrob. Resist. 2019, 18, 160–162. [Google Scholar] [CrossRef]
- Rumi, M.V.; Mas, J.; Elena, A.; Cerdeira, L.; Muñoz, M.E.; Lincopan, N.; Gentilini, É.R.; Di Conza, J.; Gutkind, G. Co-Occurrence of Clinically Relevant β-Lactamases and MCR-1 Encoding Genes in Escherichia coli from Companion Animals in Argentina. Vet. Microbiol. 2019, 230, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, P.; Yang, X.; Wang, Z.; Fanning, S.; Wang, J.; Du, P.; Bai, L. Identification of a Novel Hybrid Plasmid Coproducing MCR-1 and MCR-3 Variant from an Escherichia coli Strain. J. Antimicrob. Chemother. 2019, 74, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Du, P.; Zhang, P.; Li, Y.; Yang, X.; Wang, Z.; Wang, J.; Bai, L. Comprehensive Genomic Investigation of Coevolution of Mcr Genes in Escherichia coli Strains via Nanopore Sequencing. Glob. Chall. 2021, 5, 2000014. [Google Scholar] [CrossRef] [PubMed]
- Lentz, S.A.M.; Dalmolin, T.V.; Barth, A.L.; Martins, A.F. mcr-1 Gene in Latin America: How Is It Disseminated Among Humans, Animals, and the Environment? Front. Public Health 2021, 9, 648940. [Google Scholar] [CrossRef]
- Papa-Ezdra, R.; Grill Diaz, F.; Vieytes, M.; García-Fulgueiras, V.; Caiata, L.; Ávila, P.; Brasesco, M.; Christophersen, I.; Cordeiro, N.F.; Algorta, G.; et al. First Three Escherichia coli Isolates Harbouring mcr-1 in Uruguay. J. Glob. Antimicrob. Resist. 2020, 20, 187–190. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Class | Antibiotics | Number of mcr-1 + E. coli (%) | Gene Name (n) |
|---|---|---|---|
| Aminoglycoside | gentamicin | 8 (57) | aac(3)-IIa (2), aac(3)-IId (4), aac(3)-IVa (1), aac(3)-VIa (1) |
| hygromycin B | 1 (7) | aph(4)-Ia (1) | |
| kanamycin | 11 (79) | aph(3’)-Ia (5), aph(3’)-IIa (4), aph(6)-Id (6) | |
| streptomycin | 13 (93) | aadA2 (6), aadA5 (1), aadA15 (1), aadA17 (2), ant(3’’)-Ia (8), aph(3’’)-Ib (4) | |
| Beta-lactam | ampicillin | 10 (71) | blaTEM-1A (1), blaTEM-1B (8), blaTEM-141 (1) |
| ceftriaxone | 5 (36) | blaCTX-M-55 (5) | |
| Quinolone | ciprofloxacin | 10 (71) | qnrB19 (9), qnrD1 (1), qnrS2 (1) |
| Folate pathway antagonist | trimethoprim | 10 (71) | dfrA1 (3), dfrA8 (1), dfrA12 (6), dfrA14 (1), dfrA17 (1) |
| Fosfomycin | fosfomycin | 5 (36) | fosA3 (4), fosA6 (1) |
| Glycylcycline | tetracycline | 14 (100) | tet(A) (13), tet(B) (3), tet(D) (1), tet(M) (4) |
| lincomycin | 7 (50) | Inu(F) (7) | |
| Macrolide | erythromycin | 1 (7) | mph(A) (1) |
| Phenicol | chloramphenicol | 11 (79) | cmlA1 (6), floR (10) |
| Sulphonamide | sulfamethoxazole | 13 (93) | sul1 (3), sul2 (7), sul3 (9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carhuaricra, D.; Duran Gonzales, C.G.; Rodríguez Cueva, C.L.; Ignacion León, Y.; Silvestre Espejo, T.; Marcelo Monge, G.; Rosadio Alcántara, R.H.; Lincopan, N.; Espinoza, L.L.; Maturrano Hernández, L. Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru. Antibiotics 2022, 11, 1781. https://doi.org/10.3390/antibiotics11121781
Carhuaricra D, Duran Gonzales CG, Rodríguez Cueva CL, Ignacion León Y, Silvestre Espejo T, Marcelo Monge G, Rosadio Alcántara RH, Lincopan N, Espinoza LL, Maturrano Hernández L. Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru. Antibiotics. 2022; 11(12):1781. https://doi.org/10.3390/antibiotics11121781
Chicago/Turabian StyleCarhuaricra, Dennis, Carla G. Duran Gonzales, Carmen L. Rodríguez Cueva, Yennifer Ignacion León, Thalia Silvestre Espejo, Geraldine Marcelo Monge, Raúl H. Rosadio Alcántara, Nilton Lincopan, Luis Luna Espinoza, and Lenin Maturrano Hernández. 2022. "Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru" Antibiotics 11, no. 12: 1781. https://doi.org/10.3390/antibiotics11121781
APA StyleCarhuaricra, D., Duran Gonzales, C. G., Rodríguez Cueva, C. L., Ignacion León, Y., Silvestre Espejo, T., Marcelo Monge, G., Rosadio Alcántara, R. H., Lincopan, N., Espinoza, L. L., & Maturrano Hernández, L. (2022). Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru. Antibiotics, 11(12), 1781. https://doi.org/10.3390/antibiotics11121781