
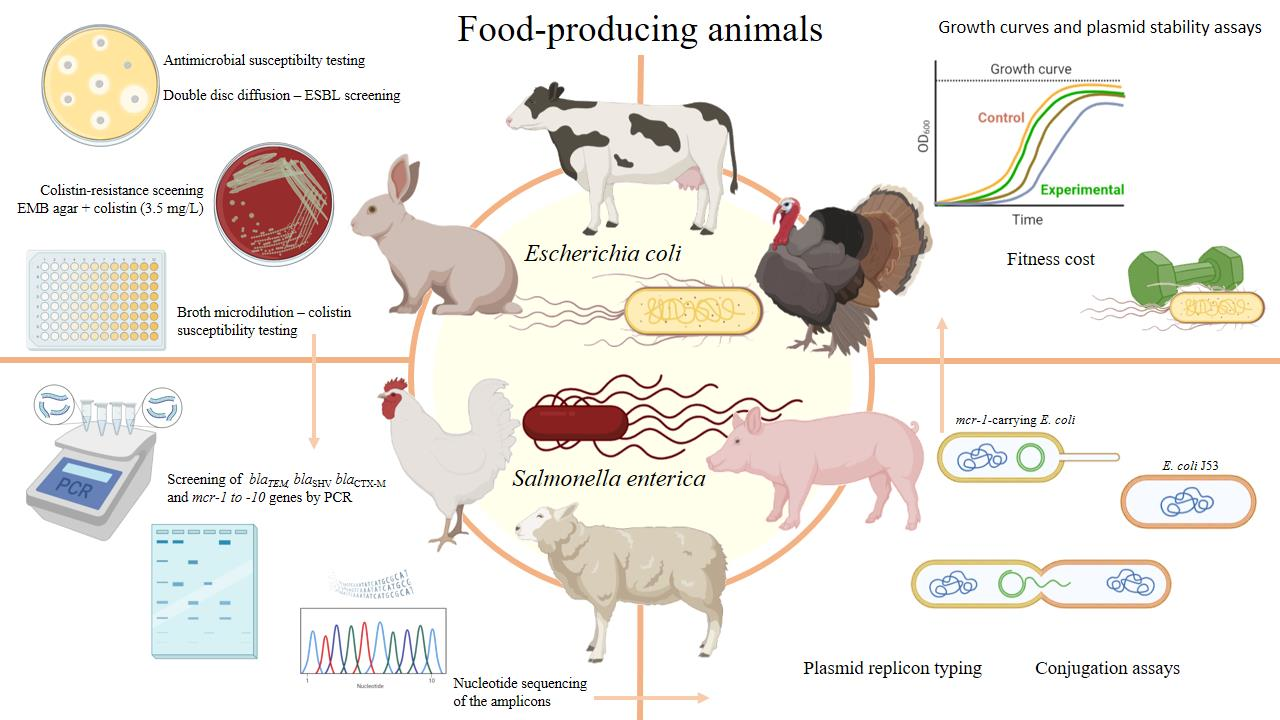
Occurrence and Biological Cost of mcr-1-Carrying Plasmids Co-harbouring Beta-Lactamase Resistance Genes in Zoonotic Pathogens from Intensive Animal Production

 ,
, 
 , and
, and 
Abstract

1. Introduction
2. Results
2.1. Origin of Bacterial Isolates and Antimicrobial Susceptibility Test
2.2. Screening of mcr and ESBL Genes
2.3. Conjugation Experiments
2.4. Growth Rates and Plasmid Stability
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Detection of Antimicrobial Resistance Genes
4.4. Conjugation Assay and Plasmid Replicon Typing
4.5. Determination of Growth Rates
4.6. Plasmid Stability Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/antimicrobial-resistance/publications/global-action-plan/en/ (accessed on 10 October 2019).
- CDC. Antibiotic Resistance Threats in the United States. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 10 December 2019).
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.; Fernandes, L.; Matias, M.; Mateus, A.; Silveira, E.; Domingues, S.; Pomba, C.; Da Silva, G.J. Longitudinal Study Detects the Co-Carriage of ESBL and mcr-1 and -4 Genes in Escherichia coli Strains in a Portuguese Farrow-to-Finish Swine Herd. Animals 2022, 12, 2209. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Theriault, W.; Letellier, A. Colistin in Pig Production: Chemistry, Mechanism of Antibacterial Action, Microbial Resistance Emergence, and One Health Perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef] [PubMed]
- EMA. Updated Advice on the Use of Colistin Products in Animals within the European Union: Development of Resistance and Possible Impact on Human and Animal Health—EMA/CVMP/CHMP/231573/2016; EMA: Amsterdam, The Netherlands, 2016. [Google Scholar]
- European Medicines Agency. Categorisation of Antibiotics in the European Union; EMA/CVMP/CHMP/682198/2017; European Medicines Agency: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Rawat, D.; Nair, D. Extended-spectrum beta-lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef] [PubMed]
- WHO. Critically Important Antimicrobials for Human Medicine—3rd Rev. Available online: https://apps.who.int/iris/bitstream/handle/10665/77376/;jsessionid=0C947CA333F9F82CF7708F537AE6FB85?sequence=1 (accessed on 10 October 2019).
- Lima, T.; Domingues, S.; Da Silva, G.J. Plasmid-Mediated Colistin Resistance in Salmonella enterica: A Review. Microorganisms 2019, 7, 55. [Google Scholar] [CrossRef]
- Zafer, M.M.; El-Mahallawy, H.A.; Abdulhak, A.; Amin, M.A.; Al-Agamy, M.H.; Radwan, H.H. Emergence of colistin resistance in multidrug-resistant Klebsiella pneumoniae and Escherichia coli strains isolated from cancer patients. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 40. [Google Scholar] [CrossRef]
- Pragasam, A.K.; Shankar, C.; Veeraraghavan, B.; Biswas, I.; Nabarro, L.E.; Inbanathan, F.Y.; George, B.; Verghese, S. Molecular Mechanisms of Colistin Resistance in Klebsiella pneumoniae Causing Bacteremia from India-A First Report. Front. Microbiol. 2016, 7, 2135. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet. Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Hadjadj, L.; Baron, S.A.; Olaitan, A.O.; Morand, S.; Rolain, J.M. Co-occurrence of Variants of mcr-3 and mcr-8 Genes in a Klebsiella pneumoniae Isolate From Laos. Front. Microbiol. 2019, 10, 2720. [Google Scholar] [CrossRef]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-producing Enterobacterales in pigs in Portugal: Evidence of direct links with antibiotic selective pressure. Int. J. Antimicrob. Agents 2019, 55, 105802. [Google Scholar] [CrossRef]
- Manageiro, V.; Clemente, L.; Romao, R.; Silva, C.; Vieira, L.; Ferreira, E.; Canica, M. IncX4 plasmid carrying the new mcr-1.9 gene variant in a CTX-M-8-producing Escherichia coli isolate recovered from swine. Front. Microbiol. 2019, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I. The biological cost of mutational antibiotic resistance: Any practical conclusions? Curr. Opin. Microbiol. 2006, 9, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, B.; Thomson, N.R.; Thomas, C.M.; Brooks, K.; Sanders, M.; Delsol, A.A.; Roe, J.M.; Bennett, P.M.; Enne, V.I. Fitness of Escherichia coli strains carrying expressed and partially silent IncN and IncP1 plasmids. BMC Microbiol. 2012, 12, 53. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef]
- Lima, T.; Domingues, S.; Da Silva, G.J. Manure as a Potential Hotspot for Antibiotic Resistance Dissemination by Horizontal Gene Transfer Events. Vet. Sci. 2020, 7, 110. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.H. Monitoring colistin resistance in food animals, an urgent threat. Expert Rev. Anti-Infect. Ther. 2018, 16, 443–446. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Cao, J.; Bi, Y.; Lv, N.; Liu, F.; Liang, S.; Shi, Y.; Jiao, X.; Gao, G.F.; et al. Antibiotic resistance gene reservoir in live poultry markets. J. Infect. 2019, 78, 445–453. [Google Scholar] [CrossRef]
- Clemente, L.; Manageiro, V.; Correia, I.; Amaro, A.; Albuquerque, T.; Themudo, P.; Ferreira, E.; Canica, M. Revealing mcr-1-positive ESBL-producing Escherichia coli strains among Enterobacteriaceae from food-producing animals (bovine, swine and poultry) and meat (bovine and swine), Portugal, 2010–2015. Int. J. Food Microbiol. 2019, 296, 37–42. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef]
- Figueiredo, R.; Card, R.M.; Nunez, J.; Pomba, C.; Mendonca, N.; Anjum, M.F.; Da Silva, G.J. Detection of an mcr-1-encoding plasmid mediating colistin resistance in Salmonella enterica from retail meat in Portugal. J. Antimicrob. Chemother. 2016, 71, 2338–2340. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.; Cristino, L.; Peixe, L.; Antunes, P. MCR-1 in multidrug-resistant and copper-tolerant clinically relevant Salmonella 1,4,[5],12:i:-and S. Rissen clones in Portugal, 2011 to 2015. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. = Eur. Commun. Dis. Bull. 2016, 21, 30270. [Google Scholar] [CrossRef] [PubMed]
- Band, V.I.; Weiss, D.S. Heteroresistance: A cause of unexplained antibiotic treatment failure? PLoS Pathog. 2019, 15, e1007726. [Google Scholar] [CrossRef] [PubMed]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Kieffer, N.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. High Rate of MCR-1-Producing Escherichia coli and Klebsiella pneumoniae among Pigs, Portugal. Emerg. Infect. Dis. 2017, 23, 2023–2029. [Google Scholar] [CrossRef]
- Freitas-Silva, J.; Inacio, A.S.; Mourao, J.; Antunes, P.; Mendes, A.; de Carvalho, A.P.; Vasconcelos, V.; Peixe, L.; da Costa, P.M. Occurrence of mcr-1 in Escherichia coli from rabbits of intensive farming. Vet. Microbiol. 2018, 227, 78–81. [Google Scholar] [CrossRef]
- Agnoletti, F.; Brunetta, R.; Bano, L.; Drigo, I.; Mazzolini, E. Longitudinal study on antimicrobial consumption and resistance in rabbit farming. Int. J. Antimicrob. Agents 2018, 51, 197–205. [Google Scholar] [CrossRef]
- Wang, X.; Zhai, Z.; Zhao, X.; Zhang, H.; Jiang, H.; Wang, X.; Wang, H.; Chang, W. Occurrence and characteristics of Escherichia coli mcr-1-like in rabbits in Shandong, China. Vet. Med. Sci. 2021, 7, 219–225. [Google Scholar] [CrossRef]
- Argudin, M.A.; Deplano, A.; Meghraoui, A.; Dodemont, M.; Heinrichs, A.; Denis, O.; Nonhoff, C.; Roisin, S. Bacteria from Animals as a Pool of Antimicrobial Resistance Genes. Antibiotics 2017, 6, 12. [Google Scholar] [CrossRef]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef]
- Tong, H.; Liu, J.; Yao, X.; Jia, H.; Wei, J.; Shao, D.; Liu, K.; Qiu, Y.; Ma, Z.; Li, B. High carriage rate of mcr-1 and antimicrobial resistance profiles of mcr-1-positive Escherichia coli isolates in swine faecal samples collected from eighteen provinces in China. Vet. Microbiol. 2018, 225, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Lopez Navas, A.; Mackay, D.; et al. Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ke, B.; Zhao, X.; Guo, Y.; Wang, W.; Wang, X.; Zhu, H. Antimicrobial Resistance Profile of mcr-1 Positive Clinical Isolates of Escherichia coli in China From 2013 to 2016. Front. Microbiol. 2018, 9, 2514. [Google Scholar] [CrossRef]
- Burow, E.; Rostalski, A.; Harlizius, J.; Gangl, A.; Simoneit, C.; Grobbel, M.; Kollas, C.; Tenhagen, B.A.; Kasbohrer, A. Antibiotic resistance in Escherichia coli from pigs from birth to slaughter and its association with antibiotic treatment. Prev. Vet. Med. 2019, 165, 52–62. [Google Scholar] [CrossRef]
- Figueiredo, R.; Henriques, A.; Sereno, R.; Mendonca, N.; da Silva, G.J. Antimicrobial resistance and extended-spectrum beta-lactamases of Salmonella enterica serotypes isolated from livestock and processed food in Portugal: An update. Foodborne Pathog. Dis. 2015, 12, 110–117. [Google Scholar] [CrossRef]
- Schulz, J.; Kemper, N.; Hartung, J.; Janusch, F.; Mohring, S.A.I.; Hamscher, G. Analysis of fluoroquinolones in dusts from intensive livestock farming and the co-occurrence of fluoroquinolone-resistant Escherichia coli. Sci. Rep. 2019, 9, 5117. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yu, L.; Chen, X.; Zhi, C.; Yao, X.; Liu, Y.; Wu, S.; Guo, Z.; Yi, L.; Zeng, Z.; et al. High Prevalence of Colistin Resistance and mcr-1 Gene in Escherichia coli Isolated from Food Animals in China. Front. Microbiol. 2017, 8, 562. [Google Scholar] [CrossRef]
- Li, X.P.; Sun, R.Y.; Song, J.Q.; Fang, L.X.; Zhang, R.M.; Lian, X.L.; Liao, X.P.; Liu, Y.H.; Lin, J.; Sun, J. Within-host heterogeneity and flexibility of mcr-1 transmission in chicken gut. Int. J. Antimicrob. Agents 2020, 55, 105806. [Google Scholar] [CrossRef]
- Brauer, A.; Telling, K.; Laht, M.; Kalmus, P.; Lutsar, I.; Remm, M.; Kisand, V.; Tenson, T. Plasmid with Colistin Resistance Gene mcr-1 in Extended-Spectrum-beta-Lactamase-Producing Escherichia coli Strains Isolated from Pig Slurry in Estonia. Antimicrob. Agents Chemother. 2016, 60, 6933–6936. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, C.; Shen, D.; Liu, J.; Ma, Z.; Yang, B.; Ling, W.; Waigi, M.G. Elimination of the risks of colistin resistance gene (mcr-1) in livestock manure during composting. Environ. Int. 2019, 126, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. MBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Kyung-Hyo, D.; Jae-Won, B.; Wan-Kyu, L. Antimicrobial Resistance Profiles of Escherichia coli from Diarrheic Weaned Piglets after the Ban on Antibiotic Growth Promoters in Feed. Antibiotics 2020, 9, 755. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, M.; Spiller, O.B.; Andrey, D.O.; Hinchliffe, P.; Li, H.; MacLean, C.; Niumsup, P.; Powell, L.; Pritchard, M.; et al. Balancing mcr-1 expression and bacterial survival is a delicate equilibrium between essential cellular defence mechanisms. Nat. Commun. 2017, 8, 2054. [Google Scholar] [CrossRef]
- Ma, K.; Feng, Y.; Zong, Z. Fitness cost of a mcr-1-carrying IncHI2 plasmid. PLoS ONE 2018, 13, e0209706. [Google Scholar] [CrossRef]
- Wu, J.; Dong, X.; Zhang, L.; Lin, Y.; Yang, K. Reversing Antibiotic Resistance Caused by Mobile Resistance Genes of High Fitness Cost. mSphere 2021, 6, e0035621. [Google Scholar] [CrossRef]
- Li, W.; Liu, Z.; Yin, W.; Yang, L.; Qiao, L.; Song, S.; Ling, Z.; Zheng, R.; Wu, C.; Wang, Y.; et al. MCR Expression Conferring Varied Fitness Costs on Host Bacteria and Affecting Bacteria Virulence. Antibiotics 2021, 10, 872. [Google Scholar] [CrossRef]
- Shen, C.; Zhong, L.L.; Yang, Y.; Doi, Y.; Paterson, D.L.; Stoesser, N.; Ma, F.; El-Sayed Ahmed, M.A.E.; Feng, S.; Huang, S.; et al. Dynamics of mcr-1 prevalence and mcr-1-positive Escherichia coli after the cessation of colistin use as a feed additive for animals in China: A prospective cross-sectional and whole genome sequencing-based molecular epidemiological study. Lancet Microbe 2020, 1, e34–e43. [Google Scholar] [CrossRef]
- Wu, R.; Yi, L.X.; Yu, L.F.; Wang, J.; Liu, Y.; Chen, X.; Lv, L.; Yang, J.; Liu, J.H. Fitness Advantage of mcr-1-Bearing IncI2 and IncX4 Plasmids in Vitro. Front. Microbiol. 2018, 9, 331. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.X. Antigenic formulae of the Salmonella serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–166. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 9.0. 2019. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_9.0_Breakpoint_Tables.pdf (accessed on 4 October 2022).
- Nordmann, P.; Jayol, A.; Poirel, L. A Universal Culture Medium for Screening Polymyxin-Resistant Gram-Negative Isolates. J. Clin. Microbiol. 2016, 54, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, N.; Leitao, J.; Manageiro, V.; Ferreira, E.; Canica, M. Spread of extended-spectrum beta-lactamase CTX-M-producing Escherichia coli clinical isolates in community and nosocomial environments in Portugal. Antimicrob. Agents Chemother. 2007, 51, 1946–1955. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. Eur. Commun. Dis. Bull. 2018, 23, 17-00672. [Google Scholar] [CrossRef]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a Novel mcr-6 to mcr-9 Multiplex PCR and Assessment of mcr-1 to mcr-9 Occurrence in Colistin-Resistant Salmonella enterica Isolates From Environment, Feed, Animals and Food (2011-2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Bryksin, A.V.; Matsumura, I. Rational design of a plasmid origin that replicates efficiently in both gram-positive and gram-negative bacteria. PLoS ONE 2010, 5, e13244. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | E. coli (n = 98) | S. enterica (n = 142) | |
|---|---|---|---|
| Poultry | S. Typhimurium | 19 | |
| S. Enteritidis | 14 | ||
| S. Anatum | 11 | ||
| S. Infantis | 11 | ||
| S. Havana | 10 | ||
| S. 4,12:i:- | 9 | ||
| S. Brandenburg | 7 | ||
| S. Typhimurium -like | 6 | ||
| S. Cerro | 4 | ||
| S. Indiana | 4 | ||
| S. Lexington | 4 | ||
| S. Bredeney | 3 | ||
| S. Rissen | 3 | ||
| S. Virchow | 3 | ||
| S. Kentucky | 2 | ||
| S. Mbandaka | 2 | ||
| S. Tennessee | 2 | ||
| S. 9,46:-:- | 1 | ||
| S. 3,15:z10: | 1 | ||
| S. Salamae 4(5),12:b- | 1 | ||
| S. Agona | 1 | ||
| S. Bardo | 1 | ||
| S. Berta | 1 | ||
| S. Hadar | 1 | ||
| S. Kottbus | 1 | ||
| S. Llandoff | 1 | ||
| S. Newport | 1 | ||
| S. Ohio | 1 | ||
| S. Seftenberg | 1 | ||
| S. Schwarzengrund | 1 | ||
| Not typed | 6 | ||
| 77 | 133 | ||
| Swine | S. Derby | 2 | |
| S. Typhimurium | 1 | ||
| Not typed | 4 | ||
| 16 | 7 | ||
| Rabbit | 3 | - | - |
| Sheep | 2 | - | - |
| Animal feed | - | Not typed | 1 |
| Bovine | - | S. Anatum | 1 |
| E. coli (n = 98) | S. enterica (n = 142) | |
|---|---|---|
| Boots swabs | 58 | |
| Carcasses/fresh meat | 42 | 20 |
| Ready to cook meat | 18 | |
| Organs | 40 | 2 |
| Rectal swabs | 6 | |
| Faeces | 6 | 24 |
| Incubator/bed swabs | 2 | 10 |
| Neck skin | 5 | |
| Abscess | 2 | |
| Frozen meat | 2 | |
| Eggs | 2 | |
| Animal feed | 1 |
| Isolate | Source of Isolates | Date of Isolation | Growth in Agar a (h) | Resistance Genes | Colistin MIC b (mg/L) | Antimicrobial Resistance c | Conjugation Frequency (Cells per Recipient) | mcr-1 Plasmid Typing |
|---|---|---|---|---|---|---|---|---|
| E. coli 162 | Chicken organs | 12/2017 | 24 | mcr-1 | 8 | AMC; AML; TE; CIP | 10−2 | P |
| E. coli 166 | Fowl carcasse | 01/2018 | 24 | mcr-1; blaTEM-1 | 16 | AMC; AML; TE; CIP | 10−4 | HI2; P |
| E. coli 170 | Turkey carcasse | 10/2017 | 24 | mcr-1; blaTEM-1 | 8 | AMC; AML; TE; CIP | 10−2 | P |
| E. coli 176 | Rabbit organs | 11/2017 | 24 | mcr-1 | 16 | AML; TE; CIP | 10−6 | I1/Iγ |
| E. coli 177 | Rabbit organs | 11/2017 | 24 | mcr-1 | 32 | AML; TE; CIP | 10−7 | HI2 |
| E. coli 182 | Chicken organs | 11/2017 | No growth | blaTEM-1; blaCTX-M-1 | N.A. | ATM; CTX; AML; TE; CIP | n.a. | n.a. |
| E. coli 185 | Swine organs | 11/2017 | 24 | mcr-1 | 16 | AML; TE; CIP | 10−7 | HI2 |
| E. coli 186 | Swine organs | 11/2017 | 48 | mcr-1; blaTEM-1; blaCTX-M-15 | 16 | ATM; CTX; AML; TE | 10−5 | HI2 |
| E. coli 189 | Swine organs | 11/2017 | 24 | mcr-1; blaCTX-M-1 | 32 | ATM; CTX; AML; TE; CIP | 10−4 | F (FIB) |
| E. coli 206 | Turkey organs | 11/2017 | 48 | blaTEM-1; blaOXY-2 | N.A. | AMC, AML; TE; CIP; CEX | n.a. | n.a. |
| E. coli 212 | Swine rectal swab | 12/2017 | 24 | mcr-1 | 16 | AML; TE; CIP | 10−3 | n.d. |
| E. coli 216 | Swine rectal swab | 12/2017 | 24 | mcr-1 | 16 | AMC; AML; TE; CIP | 10−4 | N |
| E. coli 219 | Piglet faeces | 12/2017 | 24 | mcr-1 | 16 | AML; TE | 10−2 | N |
| E. coli 220 | Piglet faeces | 12/2017 | 24 | mcr-1 | 8 | AML; TE | 10−2 | HI1 |
| E. coli 221 | Swine rectal swab | 12/2017 | 24 | mcr-1 | 16 | AML; TE; CIP | 10−3 | HI1 |
| E. coli 222 | Swine rectal swab | 12/2017 | 48 | mcr-1 | 8 | AML; TE | 10−2 | n.d. |
| E. coli 226 | Poultry carcass | 01/2018 | 24 | mcr-1 | 16 | AML; TE; CIP | 10−3 | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, T.; Loureiro, D.; Henriques, A.; Ramos, F.; Pomba, C.; Domingues, S.; da Silva, G.J. Occurrence and Biological Cost of mcr-1-Carrying Plasmids Co-harbouring Beta-Lactamase Resistance Genes in Zoonotic Pathogens from Intensive Animal Production. Antibiotics 2022, 11, 1356. https://doi.org/10.3390/antibiotics11101356
Lima T, Loureiro D, Henriques A, Ramos F, Pomba C, Domingues S, da Silva GJ. Occurrence and Biological Cost of mcr-1-Carrying Plasmids Co-harbouring Beta-Lactamase Resistance Genes in Zoonotic Pathogens from Intensive Animal Production. Antibiotics. 2022; 11(10):1356. https://doi.org/10.3390/antibiotics11101356
Chicago/Turabian StyleLima, Tiago, Dina Loureiro, Ana Henriques, Fernando Ramos, Constança Pomba, Sara Domingues, and Gabriela Jorge da Silva. 2022. "Occurrence and Biological Cost of mcr-1-Carrying Plasmids Co-harbouring Beta-Lactamase Resistance Genes in Zoonotic Pathogens from Intensive Animal Production" Antibiotics 11, no. 10: 1356. https://doi.org/10.3390/antibiotics11101356
APA StyleLima, T., Loureiro, D., Henriques, A., Ramos, F., Pomba, C., Domingues, S., & da Silva, G. J. (2022). Occurrence and Biological Cost of mcr-1-Carrying Plasmids Co-harbouring Beta-Lactamase Resistance Genes in Zoonotic Pathogens from Intensive Animal Production. Antibiotics, 11(10), 1356. https://doi.org/10.3390/antibiotics11101356