
Sesuvium portulacastrum-Mediated Removal of Nitrogen and Phosphorus Affected by Sulfadiazine in Aquaculture Wastewater


 ,
,  and
and
Abstract
:1. Introduction
2. Results
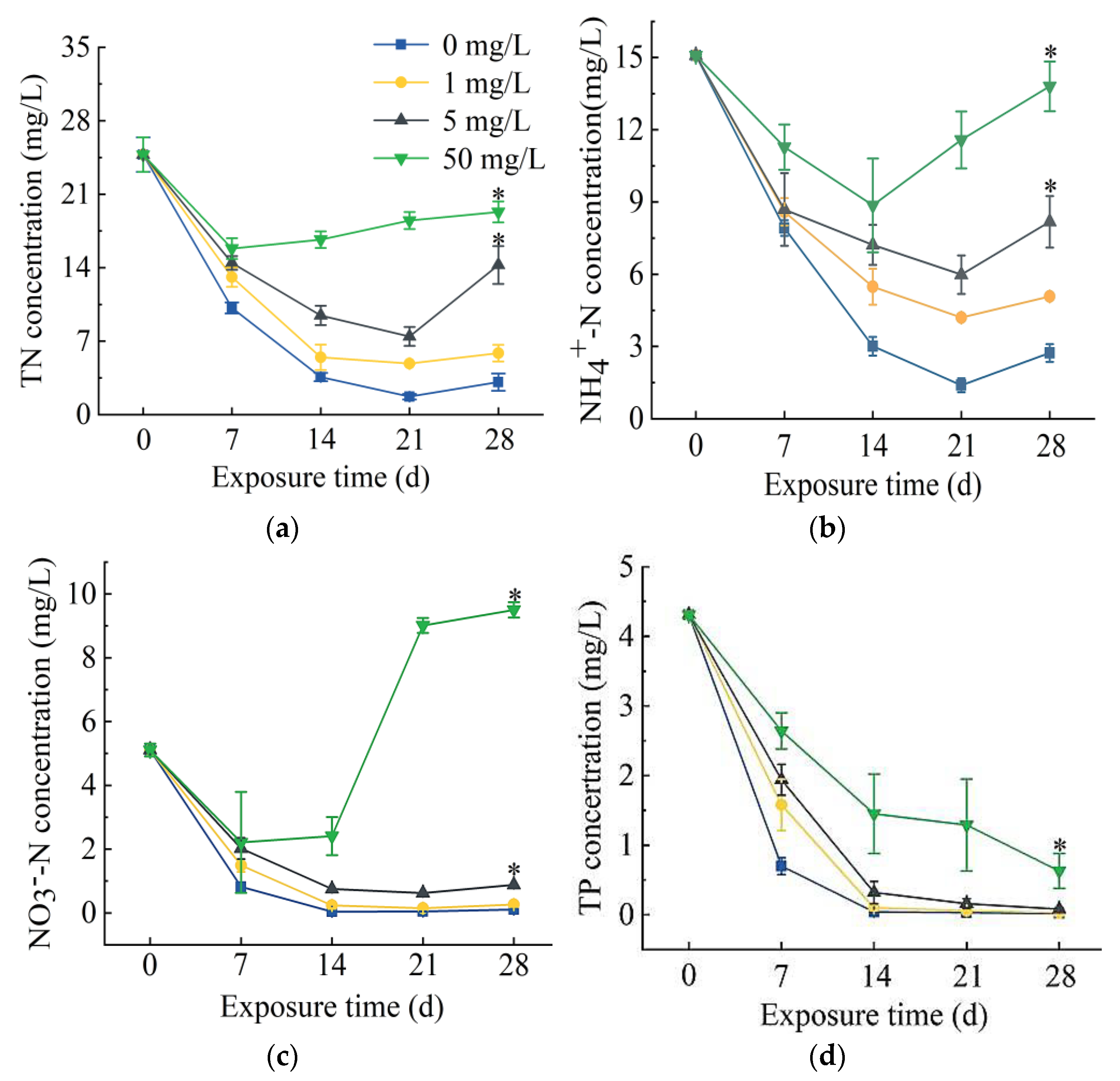
2.1. Effects of Different Concentrations of SD on S. portulacastrum-Mediated Removal of N and P
2.2. Effects of SD on Plant Growth and Physiological Responses
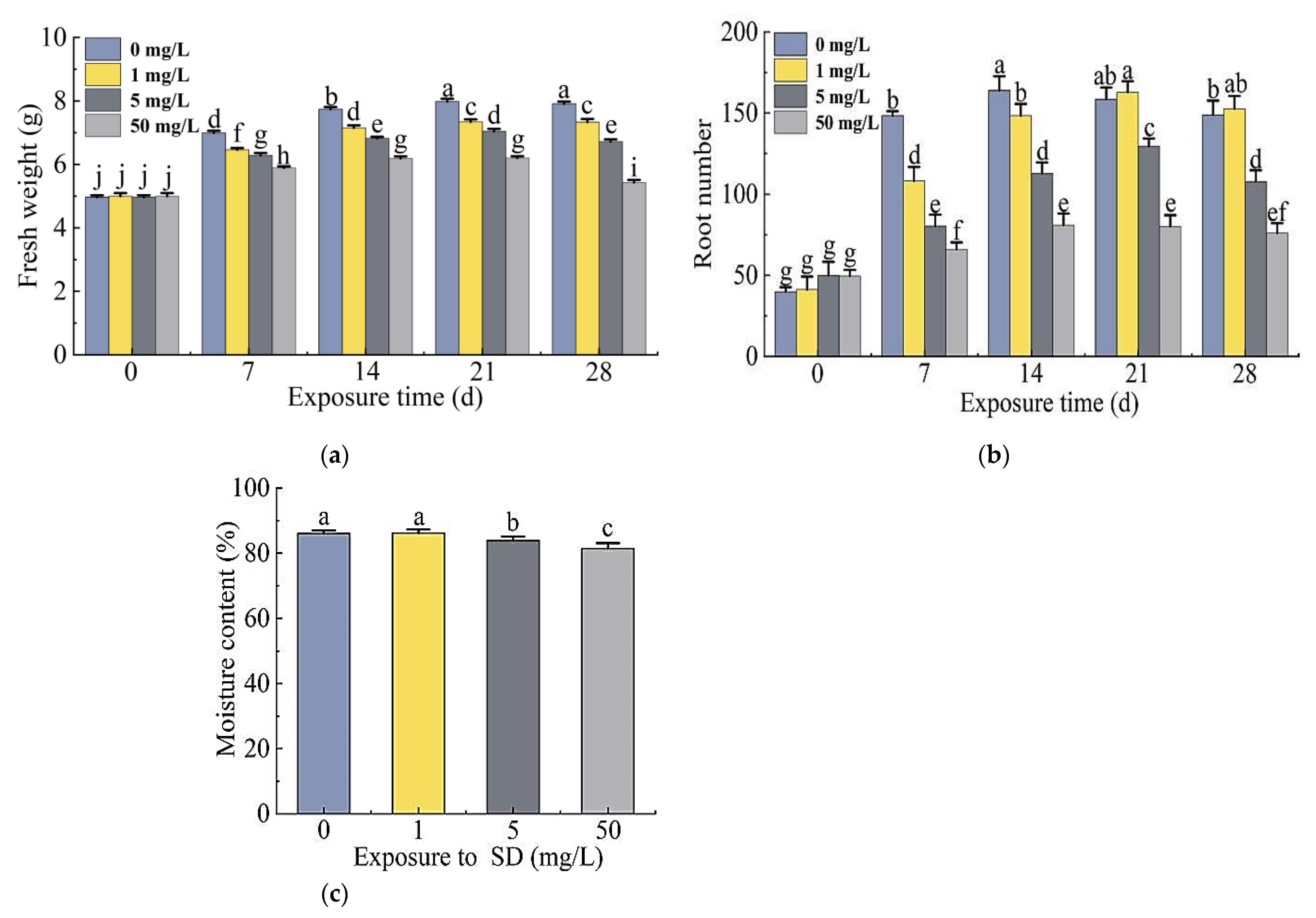
2.2.1. Growth Responses of S. portulacastrum
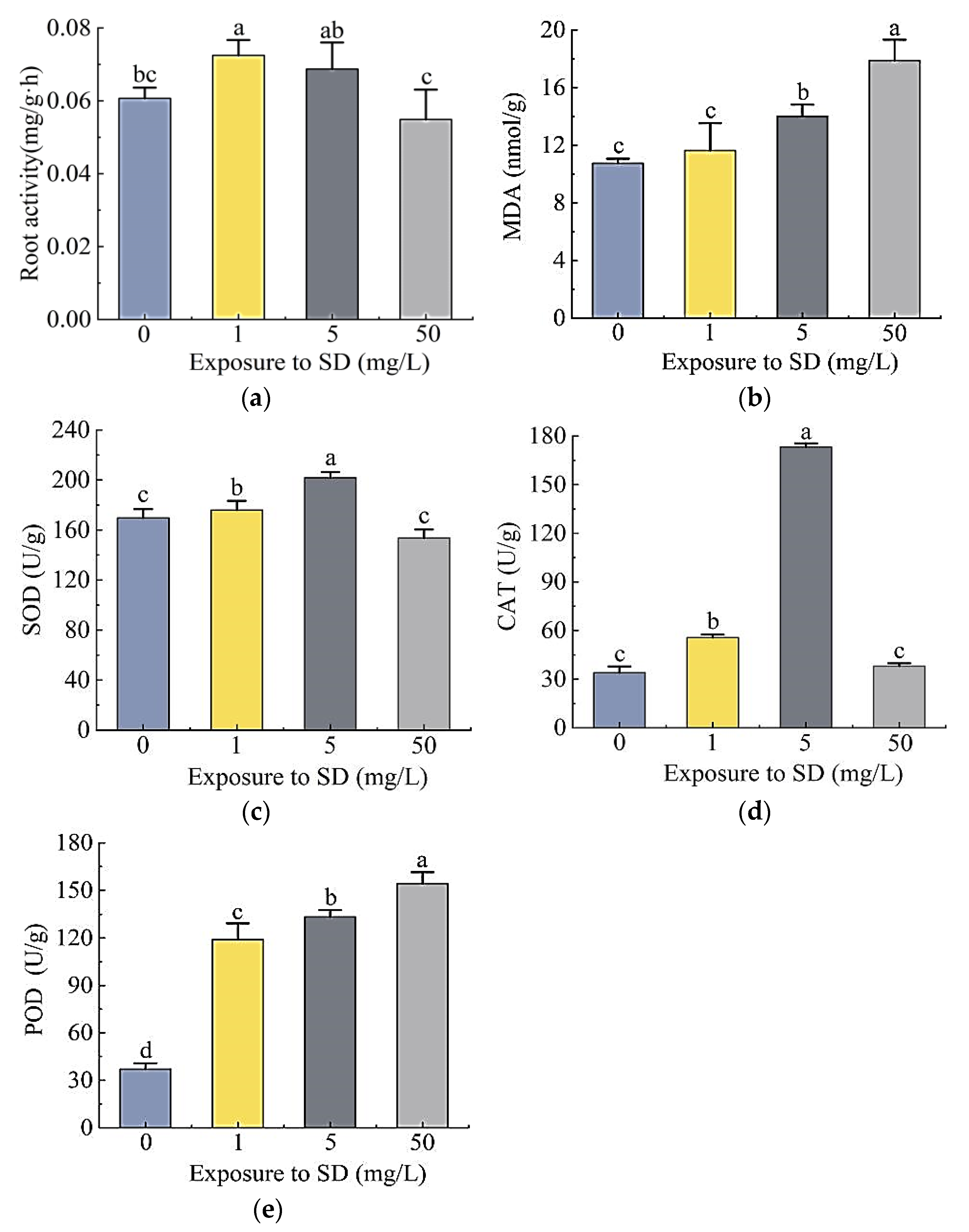
2.2.2. Physiological Responses of S. portulacastrum to SD Stress
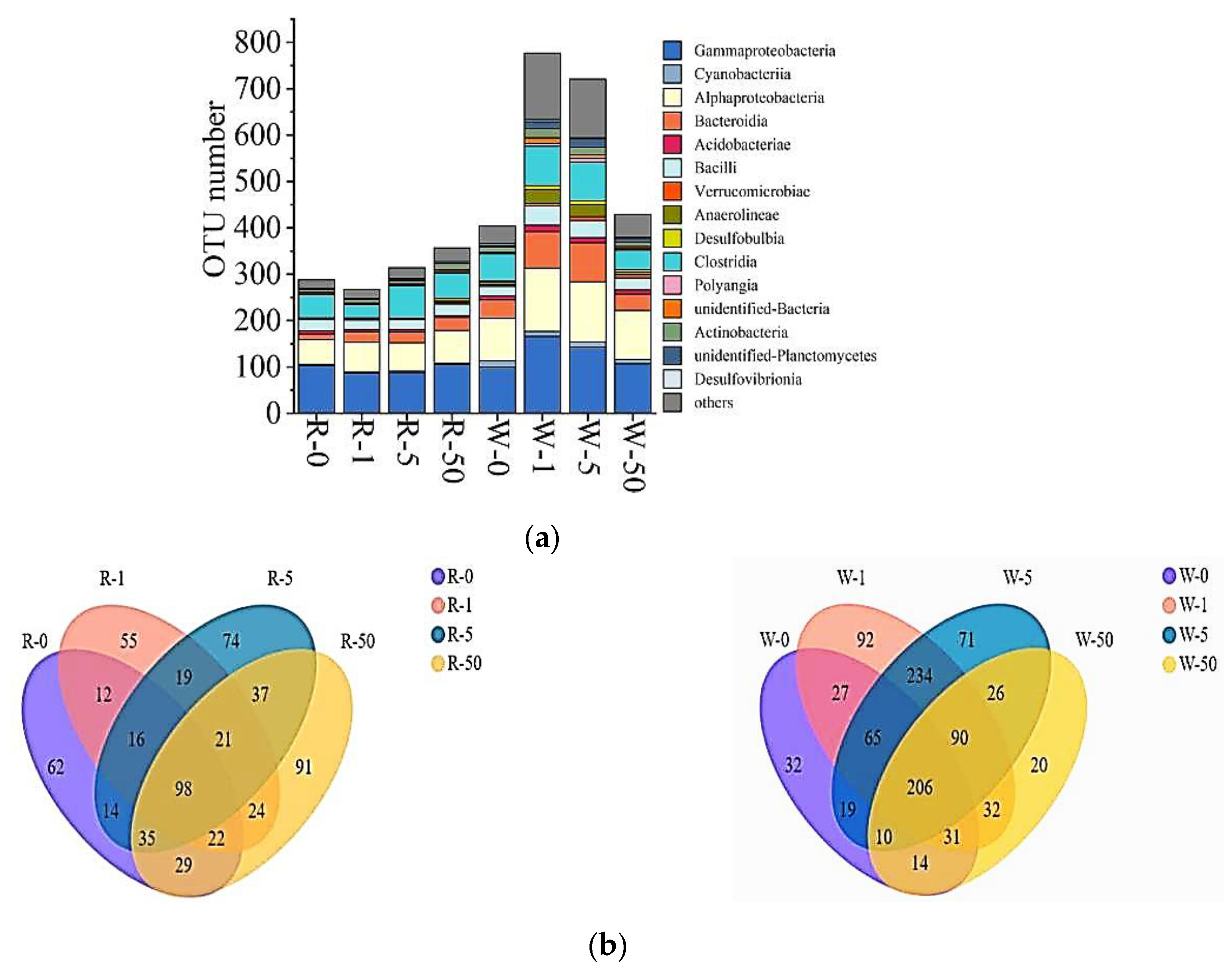
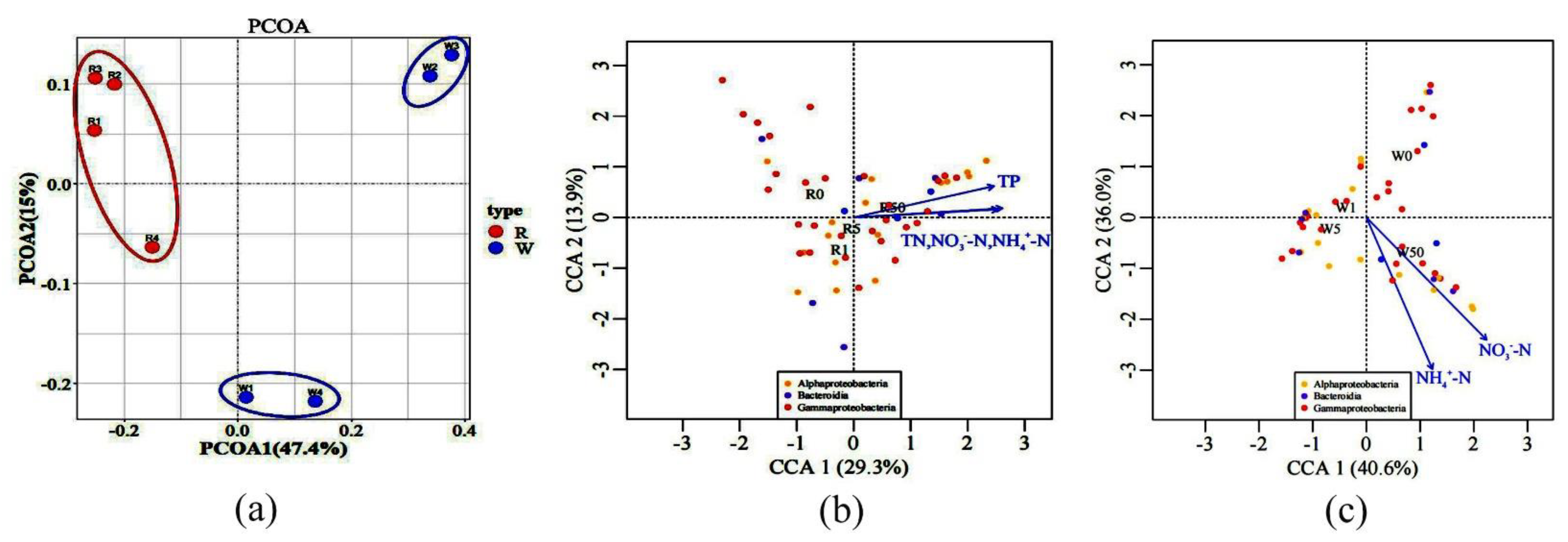
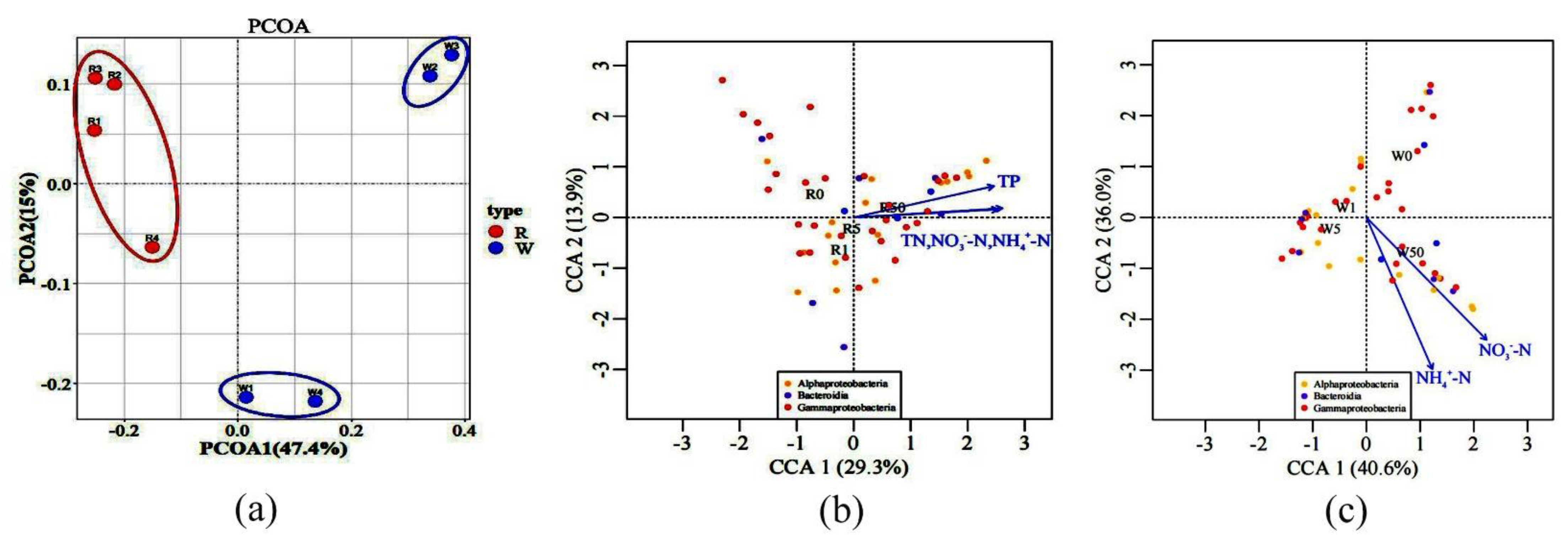
2.3. The Impact of SD on Microbial Communities
2.4. The Influence of SD on Microbial Compositions and Metabolic Activities
3. Discussion
3.1. SD Effects on Plant Growth and Physiological Activities
3.2. SD Effects on Microbial Communities
3.3. SD Effects on Plant–Microbe Interactions
4. Materials and Methods
4.1. Experimental Materials
4.1.1. Preparation of Different Concentrations of SD in Aquaculture Wastewater
4.1.2. Preparation of Plant Material
4.2. Experimental Method
4.2.1. Collection of Nutrient Salt Samples and Analysis Methods
4.2.2. Determination of Antioxidant Enzymes and Malondialdehyde in Plant Leaves and Root Activity
4.2.3. Collection of Water Samples and High-Throughput Sequencing
4.3. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018: Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Gao, F.; Li, C.; Yang, Z.; Zeng, G.; Feng, L.; Liu, J.; Liu, M.; Cai, H. Continuous microalgae cultivation in aquaculture wastewater by a membrane photobioreactor for biomass production and nutrients removal. Ecol. Eng. 2016, 92, 55–61. [Google Scholar] [CrossRef]
- Song, D.; Xu, J.; Fu, Y.; Xu, L.; Shan, B. Polysulfone/sulfonated polysulfone alloy membranes with an improved performance in processing mariculture wastewater. Chem. Eng. J. 2016, 304, 882–889. [Google Scholar] [CrossRef]
- Van Rijn, J. Waste treatment in recirculating aquaculture systems. Aquac. Eng. 2013, 53, 49–56. [Google Scholar] [CrossRef]
- Barbu, M.; Ceangă, E.; Caraman, S. Water quality modeling and control in recirculating aquaculture systems. Urban Agric. 2018, 2, 64. [Google Scholar]
- Ting, W.; Tan, I.; Salleh, S.; Wahab, N. Application of water hyacinth (eichhornia crassipes) for phytoremediation of ammoniacal nitrogen: A review. J. Water Process Eng. 2018, 22, 239–249. [Google Scholar] [CrossRef]
- Hu, H.; Zhou, Q.; Li, X.; Lou, W.; Du, C.; Teng, Q.; Zhang, D.; Liu, H.; Zhong, Y.; Yang, C. Phytoremediation of anaerobically digested swine wastewater contaminated by oxytetracycline via lemna aequinoctialis: Nutrient removal, growth characteristics and degradation pathways. Bioresour. Technol. 2019, 291, 121853. [Google Scholar] [CrossRef]
- Glick, B.R.; Karaturovic, D.M.; Newell, P.C. A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol. 1995, 41, 533–536. [Google Scholar] [CrossRef]
- Chen, H.; Liu, S.; Xu, X.; Liu, S.; Zhou, G.; Sun, K.; Zhao, J.; Ying, G. Antibiotics in typical marine aquaculture farms surrounding hailing island, south china: Occurrence, bioaccumulation and human dietary exposure. Mar. Pollut. Bull. 2015, 90, 181–187. [Google Scholar] [CrossRef]
- Liu, X.; Steele, J.C.; Meng, X. Usage, residue, and human health risk of antibiotics in chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef]
- Nguyen Dang Giang, C.; Sebesvari, Z.; Renaud, F.; Rosendahl, I.; Hoang Minh, Q.; Amelung, W. Occurrence and dissipation of the antibiotics sulfamethoxazole, sulfadiazine, trimethoprim, and enrofloxacin in the mekong delta, vietnam. PLoS ONE 2015, 10, e0131855. [Google Scholar]
- Han, Q.; Zhao, S.; Zhang, X.; Wang, X.; Song, C.; Wang, S. Distribution, combined pollution and risk assessment of antibiotics in typical marine aquaculture farms surrounding the Yellow Sea, North China. Environ. Int. 2020, 138, 105551. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Kang, Y.; Zhang, R.; Han, M.; Zeng, W.; Wang, Y.; Yu, K.; Yang, Y. Occurrence, source, and the fate of antibiotics in mariculture ponds near the Maowei Sea, South China: Storm caused the increase of antibiotics usage. Sci. Total Environ. 2021, 752, 141882. [Google Scholar] [CrossRef]
- Thuy, H.T.T.; Loan, T.T.C. Antibiotic contaminants in coastal wetlands from vietnamese shrimp farming. Environ. Sci. Pollut. Res. 2011, 18, 835–841. [Google Scholar] [CrossRef]
- Peng, Y.; Gao, F.; Yang, H.; Wu, H.; Li, C.; Lu, M.; Yang, Z. Simultaneous Removal of Nutrient And Sulfonamides From Marine Aquaculture Wastewater By Concentrated And Attached Cultivation of Chlorella Vulgaris In An Algal Biofilm Membrane Photobioreactor (Bf-Mpbr). Sci. Total Environ. 2020, 725, 138524. [Google Scholar] [CrossRef]
- Chen, J.; Xu, H.; Sun, Y.; Huang, L.; Zhang, P.; Zou, C.; Yu, B.; Zhu, G.; Zhao, C. Interspecific Differences In Growth Response And Tolerance To The Antibiotic Sulfadiazine In Ten Clonal Wetland Plants In South China. Sci. Total Environ. 2016, 543, 197–205. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Gor, B.K.; Desai, N.S.; Nikam, T.D.; Suprasanna, P. Sesuvium portulacastrum, a plant for drought, salt stress, sand fixation, food and phytoremediation: A review. Agron. Sustain. Dev. 2013, 33, 329–348. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Lin, Y.; Yang, Y.; Shen, Q.; Huang, J.; Wang, S.; Zhu, X.; Li, Z. Tolerance and bioaccumulation of cd and cu in sesuvium portulacastrum. Ecotoxicol. Environ. Saf. 2018, 147, 306–312. [Google Scholar] [CrossRef]
- Boxman, S.E.; Nystrom, M.; Capodice, J.C.; Ergas, S.J.; Main, K.L.; Trotz, M.A. Effect of support medium, hydraulic loading rate and plant density on water quality and growth of halophytes in marine aquaponic systems. Aquac. Res. 2017, 48, 2463–2477. [Google Scholar] [CrossRef]
- Liu, X.; Pu, X.; Luo, D.; Lu, J.; Liu, Z. Model assessment of nutrient removal via planting sesuvium portulacastrum in floating beds in eutrophic marine waters: The case of aquaculture areas of dongshan bay. Acta Oceanol. Sin. 2019, 38, 91–100. [Google Scholar] [CrossRef]
- Polomski, R.F.; Taylor, M.D.; Bielenberg, D.G.; Bridges, W.C.; Klaine, S.J.; Whitwell, T. Nitrogen and phosphorus remediation by three floating aquatic macrophytes in greenhouse-based laboratory-scale subsurface constructed wetlands. Water Air Soil Pollut. 2009, 197, 223–232. [Google Scholar] [CrossRef]
- Körner, S.; Vermaat, J. The relative importance of Lemna gibba L., bacteria and algae for the nitrogen and phosphorus removal in duckweed-covered domestic wastewater. Water Res. 1998, 32, 3651–3661. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, H.; Xiao, W.; Lu, J.; Lu, D.; Chen, X.; Zheng, X.; Jeppesen, E.; Zhang, W.; Wang, L. Ecotoxicological effects of sulfonamide on and its removal by the submerged plant Vallisneria natans (Lour.) hara. Water Res. 2020, 170, 115354. [Google Scholar] [CrossRef] [PubMed]
- Michelini, L.; Reichel, R.; Werner, W.; Ghisi, R.; Thiele-Bruhn, S. Sulfadiazine uptake and effects on Salix fragilis L. and Zea mays L. plants. Water Air Soil Pollut. 2012, 223, 5243–5257. [Google Scholar] [CrossRef]
- Michelini, L.; La Rocca, N.; Rascio, N.; Ghisi, R. Structural and functional alterations induced by two sulfonamide antibiotics on barley plants. Plant Physiol. Biochem. 2013, 67, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fuego, D.; Keunen, E.; Cuypers, A.; Bertrand, A.; González, A. Mycorrhization protects betula pubescens ehr. From metal-induced oxidative stress increasing its tolerance to grow in an industrial polluted soil. J. Hazard. Mater. 2017, 336, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Spengler, A.; Wanninger, L.; Pflugmacher, S. Oxidative stress mediated toxicity of TiO2 nanoparticles after a concentration and time dependent exposure of the aquatic macrophyte hydrilla verticillata. Aquat. Toxicol. 2017, 190, 32–39. [Google Scholar] [CrossRef]
- Zhao, S.; He, L.; Lu, Y.; Duo, L. The impact of modified nano-carbon black on the earthworm eisenia fetida under turfgrass growing conditions: Assessment of survival, biomass, and antioxidant enzymatic activities. J. Hazard. Mater. 2017, 338, 218–223. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M.; Słaba, M.; Mazur, J. Effect of nickel on antioxidative enzyme activities, proline and chlorophyll contents in wheat shoots. Biol. Plant. 2006, 50, 653–659. [Google Scholar] [CrossRef]
- Deng, X.; Gao, K.; Sun, J. Physiological and biochemical responses of Synechococcus sp. pcc7942 to irgarol 1051 and diuron. Aquat. Toxicol. 2012, 122, 113–119. [Google Scholar]
- Tang, C.; Liu, Y.; Zeng, G.; Li, X.; Xu, W.; Li, C.; Yuan, X. Effects of exogenous spermidine on antioxidant system responses of Typha latifolia L. under CD2+ stress. J. Integr. Plant Biol. 2005, 47, 428–434. [Google Scholar] [CrossRef]
- Qian, H.; Chen, W.; Sheng, G.; Xu, X.; Liu, W.; Fu, Z. Effects of glufosinate on antioxidant enzymes, subcellular structure, and gene expression in the unicellular green alga chlorella vulgaris. Aquat. Toxicol. 2008, 88, 301–307. [Google Scholar] [CrossRef]
- Singh, R.; Upadhyay, A.; Chandra, P.; Singh, D. Sodium chloride incites reactive oxygen species in green algae chlorococcum humicola and chlorella vulgaris: Implication on lipid synthesis, mineral nutrients and antioxidant system. Bioresour. Technol. 2018, 270, 489–497. [Google Scholar] [CrossRef]
- Xu, Q. Pollution Characteristics of Typical Contaminates and Physiological Effects of Antibiotics in the Soil-Vegtable System. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2019. [Google Scholar]
- Liu, X.; Lv, Y.; Xu, K.; Xiao, X.; Xi, B.; Lu, S. Response of ginger growth to a tetracycline-contaminated environment and residues of antibiotic and antibiotic resistance genes. Chemosphere 2018, 201, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Van Donk, E.; Gulati, R.D.; Iedema, A.; Meulemans, J.T. Macrophyte-related shifts in the nitrogen and phosphorus contents of the different trophic levels in a biomanipulated shallow lake. In Nutrient Dynamics and Retention in Land/Water Ecotones of Lowland, Temperate Lakes and Rivers; Springer: Dordrecht, The Netherlands, 1993; pp. 19–26. [Google Scholar]
- Barko, J.W.; James, W.F. Effects of submerged aquatic macrophytes on nutrient dynamics, sedimentation, and resuspension. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 197–214. [Google Scholar]
- Lin, H.; Jin, D.; Freitag, T.; Sun, W.; Yu, Q.; Fu, J.; Ma, J. A compositional shift in the soil microbiome induced by tetracycline, sulfamonomethoxine and ciprofloxacin entering a plant-soil system. Environ. Pollut. 2016, 212, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Kristiansson, E.; Fick, J.; Janzon, A.; Grabic, R.; Rutgersson, C.; Weijdegård, B.; Söderström, H.; Larsson, D.J. Pyrosequencing of antibiotic-contaminated river sediments reveals high levels of resistance and gene transfer elements. PLoS ONE 2011, 6, e17038. [Google Scholar] [CrossRef]
- Novo, A.; André, S.; Viana, P.; Nunes, O.C.; Manaia, C.M. Antibiotic resistance, antimicrobial residues and bacterial community composition in urban wastewater. Water Res. 2013, 47, 1875–1887. [Google Scholar] [CrossRef] [Green Version]
- Hovanec, T.A.; Taylor, L.T.; Blakis, A.; Delong, E.F. Nitrospira-like bacteria associated with nitrite oxidation in freshwater aquaria. Appl. Environ. Microbiol. 1998, 64, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Nold, S.C.; Zhou, J.; Devol, A.H.; Tiedje, J.M. Pacific northwest marine sediments contain ammonia-oxidizing bacteria in the β subdivision of the proteobacteria. Appl. Environ. Microbiol. 2000, 66, 4532–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chon, K.; Kim, Y.; Chang, N.; Cho, J. Evaluating wastewater stabilizing constructed wetland, through diversity and abundance of the nitrite reductase gene nirs, with regard to nitrogen control. Desalination 2010, 264, 201–205. [Google Scholar] [CrossRef]
- Sharma, V.K.; Johnson, N.; Cizmas, L.; Mcdonald, T.J.; Kim, H. A review of the influence of treatment strategies on antibiotic resistant bacteria and antibiotic resistance genes. Chemosphere 2016, 150, 702–714. [Google Scholar] [CrossRef]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Li, B.; Zou, R.; Dai, Y.; Xie, S.; Yuan, B. Biodegradation of antibiotic ciprofloxacin: Pathways, influential factors, and bacterial community structure. Environ. Sci. Pollut. Res. 2016, 23, 7911–7918. [Google Scholar] [CrossRef]
- Deng, Z.; Zhao, L.; Kong, Z.; Yang, W.; Lindström, K.; Wang, E.; Wei, G. Diversity of endophytic bacteria within nodules of the sphaerophysa salsula in different regions of loess plateau in China. FEMS Microbiol. Ecol. 2011, 76, 463–475. [Google Scholar] [CrossRef]
- Rahman, M.M.; Flory, E.; Koyro, H.; Abideen, Z.; Schikora, A.; Suarez, C.; Schnell, S.; Cardinale, M. Consistent associations with beneficial bacteria in the seed endosphere of barley (Hordeum vulgare L.). Syst. Appl. Microbiol. 2018, 41, 386–398. [Google Scholar] [CrossRef]
- Santos-Medellín, C.; Edwards, J.; Liechty, Z.; Nguyen, B.; Sundaresan, V. Drought stress results in a compartment-specific restructuring of the rice root-associated microbiomes. mBio 2017, 8, e00764-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, N.; Yong-Xin, L.; Zhang, X.; Hu, B.; Qin, Y.; Xu, H.; Wang, H.; Guo, X.; Qian, J. Root microbiota shift in rice correlates with resident time in the field and developmental stage. Sci. China Life Sci. 2018, 61, 613–621. [Google Scholar] [CrossRef]
- Tan, Y. Rhizosphere Microbial Diversity, Ecological Function and Plant-Microbial Interaction Mechanism of Salt-Tolerant Ecological Floating Bed Halophyte Sesuvium Portulacastrum. Master’s Thesis, Xiamen University, Xiamen, China, 2019. [Google Scholar]
- Walters, W.A.; Jin, Z.; Youngblut, N.; Wallace, J.G.; Sutter, J.; Zhang, W.; González-Peña, A.; Peiffer, J.; Koren, O.; Shi, Q. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 7368–7373. [Google Scholar] [CrossRef] [Green Version]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Snyder, W.C. Nutrition of strawberry plant under controlled conditions. (a) Effects of deficiencies of boron and certain other elements, (b) susceptibility to injury from sodium salts. Proc. Am. Soc. Hortic. Sci. 1933, 30, 288–294. [Google Scholar]
- China, E. Methods for Monitoring and Analysis of Water and Wastewater; Chinese Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. Vsearch: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Edgar, R.C. Uparse: Highly accurate otu sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rrna sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample/Index | ACE | Shannon | Chao 1 | |
|---|---|---|---|---|
| R | R-0 | 340.07 | 1.56 | 332.38 |
| R-1 | 304.30 | 1.62 | 297.69 | |
| R-5 | 370.86 | 1.24 | 357.56 | |
| R-50 | 421.08 | 1.57 | 416.38 | |
| W | W-0 | 465.31 | 1.95 | 441.67 |
| W-1 | 842.95 | 3.73 | 846.00 | |
| W-5 | 739.91 | 4.02 | 727.62 | |
| W-50 | 516.71 | 2.99 | 519.02 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Wang, D.; He, W.; Liu, H.; Chen, J.; Wei, X.; Mu, J. Sesuvium portulacastrum-Mediated Removal of Nitrogen and Phosphorus Affected by Sulfadiazine in Aquaculture Wastewater. Antibiotics 2022, 11, 68. https://doi.org/10.3390/antibiotics11010068
Zhang C, Wang D, He W, Liu H, Chen J, Wei X, Mu J. Sesuvium portulacastrum-Mediated Removal of Nitrogen and Phosphorus Affected by Sulfadiazine in Aquaculture Wastewater. Antibiotics. 2022; 11(1):68. https://doi.org/10.3390/antibiotics11010068
Chicago/Turabian StyleZhang, Chaoyue, Dan Wang, Weihong He, Hong Liu, Jianjun Chen, Xiangying Wei, and Jingli Mu. 2022. "Sesuvium portulacastrum-Mediated Removal of Nitrogen and Phosphorus Affected by Sulfadiazine in Aquaculture Wastewater" Antibiotics 11, no. 1: 68. https://doi.org/10.3390/antibiotics11010068
APA StyleZhang, C., Wang, D., He, W., Liu, H., Chen, J., Wei, X., & Mu, J. (2022). Sesuvium portulacastrum-Mediated Removal of Nitrogen and Phosphorus Affected by Sulfadiazine in Aquaculture Wastewater. Antibiotics, 11(1), 68. https://doi.org/10.3390/antibiotics11010068