The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia


Abstract
:1. Introduction
2. Results
2.1. Prevalence of ESBL-Producing E. coli
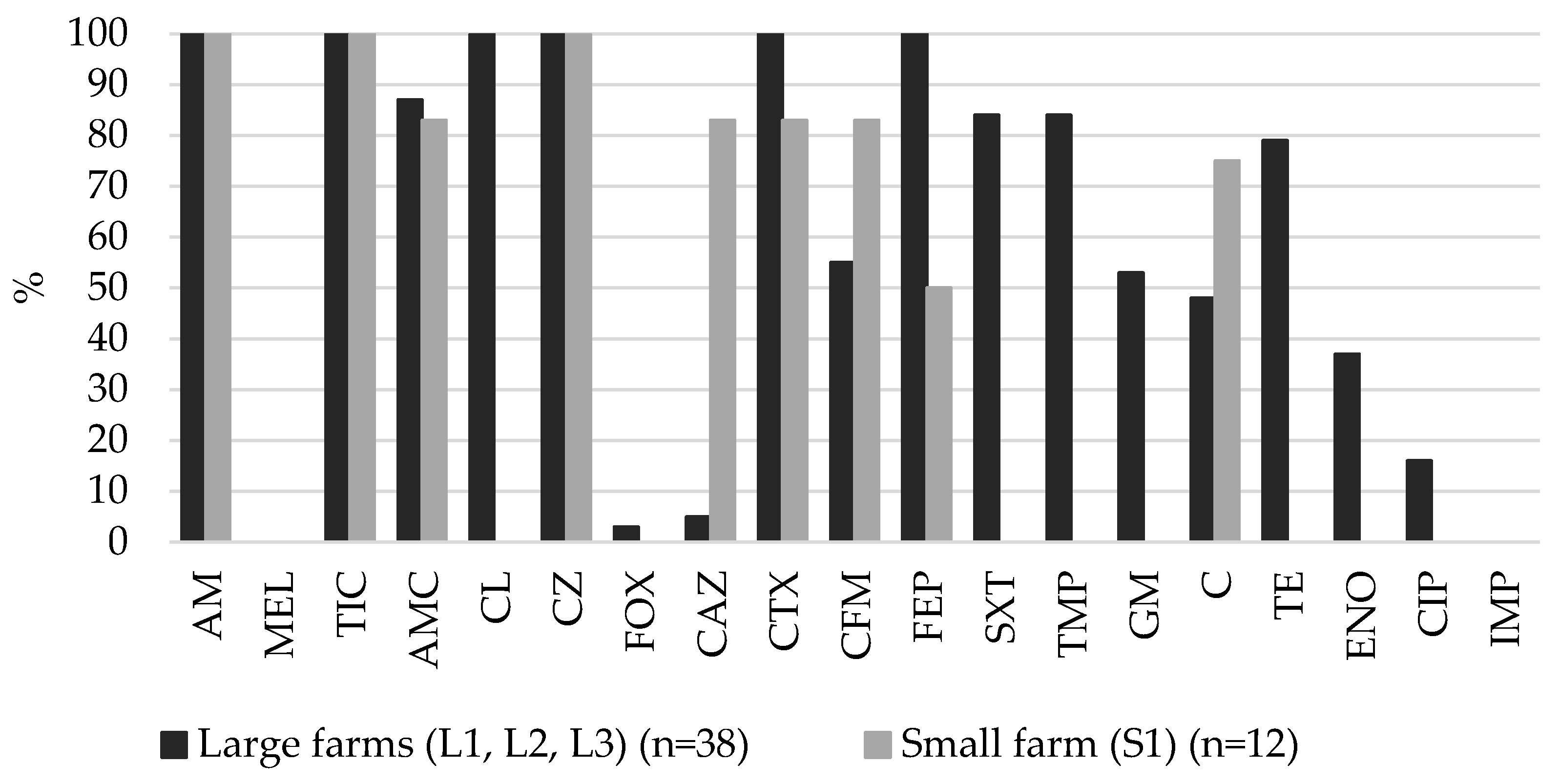
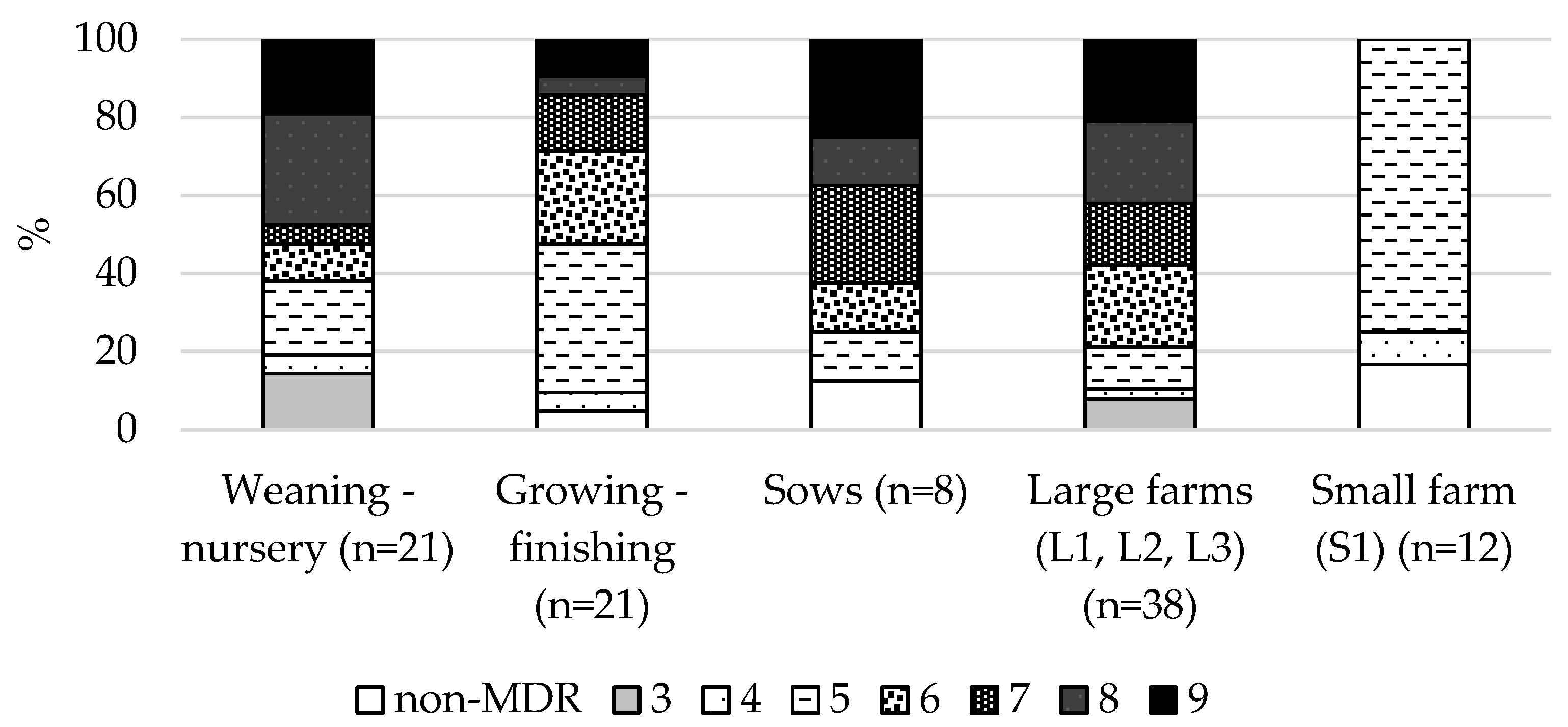
2.2. Resistance of ESBL-Producing E. coli Isolates Tested
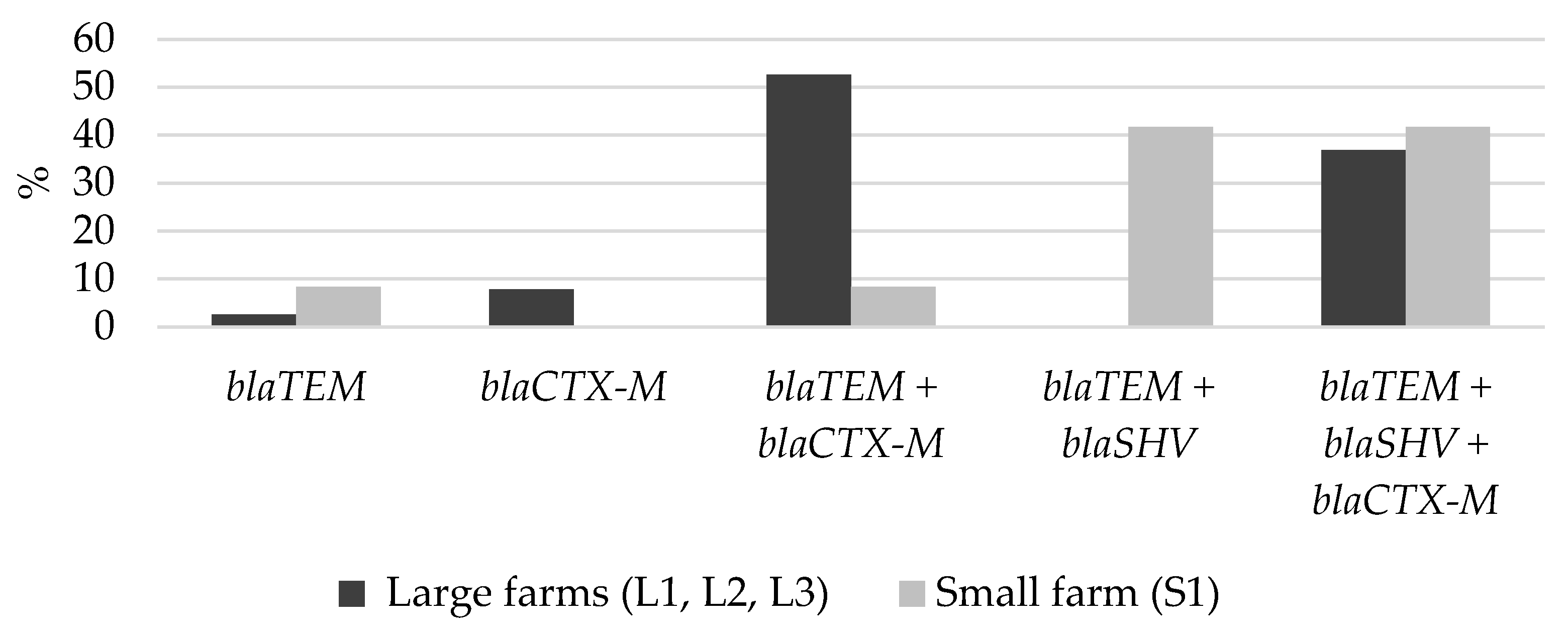
2.3. Distribution of Beta-Lactamase Gene(s) in ESBL-Producing E. coli
2.4. Associations of Beta-Lactamase Gene(s) to Phenotypic Antimicrobial
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Characterization of Farms
4.2. Screening of Extended-Spectrum-Beta-Lactamase-Producing E. coli
4.3. Phenotypic Confirmatory Test for ESBLs in E. coli
4.4. Antimicrobial Susceptibility Testing
4.5. Identification of Beta-Lactamase Genes in the Fecal ESBL-Producing E. coli
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EDCD; EFSA; EMA. ECDC/EFSA/EMA Second Joint Report on the Integrated Analysis of the Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-producing Animals. EFSA J. 2017, 15, e04872. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, D.G.D.; Dewulf, J.; Piñeiro, C.; Edwards, S.; Kyriazakis, I. A Critical Reflection on Intensive Pork Production with an Emphasis on Animal Health and Welfare. J. Anim. Sci. 2020, 98 (Suppl. 1), S15–S26. [Google Scholar] [CrossRef] [PubMed]
- EFSA; ECDC. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2017/2018. EFSA J. 2020, 18, e06007. [Google Scholar] [CrossRef] [Green Version]
- ESVAC. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2018; European Medicines Agency: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Silva, N.; Carvalho, I.; Currie, C.; Sousa, M.; Igrejas, G.; Poeta, P. Extended-Spectrum-β-Lactamase and Carbapenemase-Producing Enterobacteriaceae in Food-Producing Animals in Europe: An impact on public health? In Antibiotic Drug Resistance; Capelo-Martinez, J.-L., Igrejas, G., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 261–273. [Google Scholar] [CrossRef]
- Carvalho, I.; Cunha, R.; Martins, C.; Martínez-Álvarez, S.; Safia Chenouf, N.; Pimenta, P.; Pereira, A.R.; Ramos, S.; Sadi, M.; Martins, Â.; et al. Antimicrobial Resistance Genes and Diversity of Clones among Faecal ESBL-Producing Escherichia coli Isolated from Healthy and Sick Dogs Living in Portugal. Antibiotics 2021, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Silva, V.; Dapkevicius, M.d.L.E.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, 2239. [Google Scholar] [CrossRef] [PubMed]
- WHO. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug Resistant Bacterial Infections, Including Tuberculosis; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef] [PubMed]
- De Koster, S.; Ringenier, M.; Lammens, C.; Stegeman, A.; Tobias, T.; Velkers, F.; Vernooij, H.; Kluytmans-van den Bergh, M.; Kluytmans, J.; Dewulf, J.; et al. ESBL-Producing, Carbapenem- and Ciprofloxacin-Resistant Escherichia coli in Belgian and Dutch Broiler and Pig Farms: A Cross-Sectional and Cross-Border Study. Antibiotics 2021, 10, 945. [Google Scholar] [CrossRef] [PubMed]
- van der Fels-Klerx, H.J.; Puister-Jansen, L.F.; van Asselt, E.D.; Burgers, S.L.G.E. Farm Factors Associated with the Use of Antibiotics in Pig Production1. J. Anim. Sci. 2011, 89, 1922–1929. [Google Scholar] [CrossRef] [Green Version]
- Ström, G.; Halje, M.; Karlsson, D.; Jiwakanon, J.; Pringle, M.; Fernström, L.-L.; Magnusson, U. Antimicrobial Use and Antimicrobial Susceptibility in Escherichia coli on Small- and Medium-Scale Pig Farms in North-Eastern Thailand. Antimicrob. Resist. Infect. Control 2017, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Dohmen, W.; Bonten, M.J.M.; Bos, M.E.H.; van Marm, S.; Scharringa, J.; Wagenaar, J.A.; Heederik, D.J.J. Carriage of Extended-Spectrum β-Lactamases in Pig Farmers Is Associated with Occurrence in Pigs. Clin. Microbiol. Infect. 2015, 21, 917–923. [Google Scholar] [CrossRef] [Green Version]
- Moor, J.; Aebi, S.; Rickli, S.; Mostacci, N.; Overesch, G.; Oppliger, A.; Hilty, M. Dynamics of Extended-Spectrum Cephalosporin-Resistant Escherichia coli in Pig Farms: A Longitudinal Study. Int. J. Antimicrob. Agents 2021, 48, 106382. [Google Scholar] [CrossRef]
- Schmid, A.; Hörmansdorfer, S.; Messelhäusser, U.; Käsbohrer, A.; Sauter-Louis, C.; Mansfeld, R. Prevalence of Extended-Spectrum β-Lactamase-Producing Escherichia coli on Bavarian Dairy and Beef Cattle Farms. Appl. Environ. Microbiol. 2013, 79, 3027–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmithausen, R.M.; Schulze-Geisthoevel, S.V.; Stemmer, F.; El-Jade, M.; Reif, M.; Hack, S.; Meilaender, A.; Montabauer, G.; Fimmers, R.; Parcina, M.; et al. Analysis of Transmission of MRSA and ESBL-E among Pigs and Farm Personnel. PLoS ONE 2015, 10, e0138173. [Google Scholar] [CrossRef]
- Dohmen, W.; Dorado-García, A.; Bonten, M.J.M.; Wagenaar, J.A.; Mevius, D.; Heederik, D.J.J. Risk Factors for ESBL-Producing Escherichia coli on Pig Farms: A Longitudinal Study in the Context of Reduced Use of Antimicrobials. PLoS ONE 2017, 12, e0174094. [Google Scholar] [CrossRef]
- Gay, N.; Leclaire, A.; Laval, M.; Miltgen, G.; Jégo, M.; Stéphane, R.; Jaubert, J.; Belmonte, O.; Cardinale, E. Risk Factors of Extended-Spectrum β-Lactamase Producing Enterobacteriaceae Occurrence in Farms in Reunion, Madagascar and Mayotte Islands, 2016–2017. Vet. Sci. 2018, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Reyns, T.; De Boever, S.; Schauvliege, S.; Gasthuys, F.; Meissonnier, G.; Oswald, I.; De Backer, P.; Croubels, S. Influence of Administration Route on the Biotransformation of Amoxicillin in the Pig. J. Vet. Pharmacol. Ther. 2009, 32, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Burch, D.G.S.; Sperling, D. Amoxicillin-Current Use in Swine Medicine. J. Vet. Pharmacol. Ther. 2018, 41, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-Producing Enterobacterales in Pigs in Portugal: Evidence of Direct Links with Antibiotic Selective Pressure. Int. J. Antimicrob. Agents 2020, 55, 105802. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Mukherjee, M. Incidence and Risk of Co-Transmission of Plasmid-Mediated Quinolone Resistance and Extended-Spectrum β-Lactamase Genes in Fluoroquinolone-Resistant Uropathogenic Escherichia coli: A First Study from Kolkata, India. J. Glob. Antimicrob. Resist. 2018, 14, 217–223. [Google Scholar] [CrossRef]
- Livermore, D.M.; Day, M.; Cleary, P.; Hopkins, K.L.; Toleman, M.A.; Wareham, D.W.; Wiuff, C.; Doumith, M.; Woodford, N. OXA-1 β-Lactamase and Non-Susceptibility to Penicillin/β-Lactamase Inhibitor Combinations among ESBL-Producing Escherichia coli. J. Antimicrob. Chemother. 2019, 74, 326–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, D.; Nair, D. Extended-Spectrum β-Lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A Review of SHV Extended-Spectrum β-Lactamases: Neglected Yet Ubiquitous. Front. Microbiol. 2016, 7, 1374. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, R. Growing Group of Extended-Spectrum β-Lactamases: The CTX-M Enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeynudin, A.; Pritsch, M.; Schubert, S.; Messerer, M.; Liegl, G.; Hoelscher, M.; Belachew, T.; Wieser, A. Prevalence and Antibiotic Susceptibility Pattern of CTX-M Type Extended-Spectrum β-Lactamases among Clinical Isolates of Gram-Negative Bacilli in Jimma, Ethiopia. BMC Infect. Dis. 2018, 18, 524. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Graells, C.; Berbers, B.; Verhaegen, B.; Vanneste, K.; Marchal, K.; Roosens, N.H.C.; Botteldoorn, N.; De Keersmaecker, S.C.J. First Detection of a Plasmid Located Carbapenem Resistant BlaVIM-1 Gene in E. coli Isolated from Meat Products at Retail in Belgium in 2015. Int. J. Food Microbiol. 2020, 324, 108624. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.F. Beta-lactam Antibiotics. In Antimicrobial Therapy in Veterinary Medicine; Giguère, S., Prescott, J.F., Dowling, P.M., Eds.; John Wiley & Sons: Amsterdam, The Netherlands, 2013; pp. 133–152. [Google Scholar] [CrossRef]
- Fuchs, F.; Hamprecht, A. Results from a Prospective In Vitro Study on the Mecillinam (Amdinocillin) Susceptibility of Enterobacterales. Antimicrob. Agents Chemother. 2019, 63, e02402-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EMA; CVMP; CHMP. Categorisation of Antibiotics in the European Union; EMA: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Hölzel, C.S.; Müller, C.; Harms, K.S.; Mikolajewski, S.; Schäfer, S.; Schwaiger, K.; Bauer, J. Heavy Metals in Liquid Pig Manure in Light of Bacterial Antimicrobial Resistance. Environ. Res. 2012, 113, 21–27. [Google Scholar] [CrossRef]
- Ciesinski, L.; Guenther, S.; Pieper, R.; Kalisch, M.; Bednorz, C.; Wieler, L.H. High Dietary Zinc Feeding Promotes Persistence of Multi-Resistant E. coli in the Swine Gut. PLoS ONE 2018, 13, e0191660. [Google Scholar] [CrossRef]
- Akwar, H.T.; Poppe, C.; Wilson, J.; Reid-Smith, R.J.; Dyck, M.; Waddington, J.; Shang, D.; McEwen, S.A. Prevalence and Patterns of Antimicrobial Resistance of Fecal Escherichia coli among Pigs on 47 Farrow-to-Finish Farms with Different in-Feed Medication Policies in Ontario and British Columbia. Can. J. Vet. Res. = Rev. Can. Rech. Vet. 2008, 72, 195–201. [Google Scholar]
- De Lucia, A.; Card, R.M.; Duggett, N.; Smith, R.P.; Davies, R.; Cawthraw, S.A.; Anjum, M.F.; Rambaldi, M.; Ostanello, F.; Martelli, F. Reduction in Antimicrobial Resistance Prevalence in Escherichia coli from a Pig Farm Following Withdrawal of Group Antimicrobial Treatment. Vet. Microbiol. 2021, 258, 109125. [Google Scholar] [CrossRef]
- Jensen, V.F.; Emborg, H.-D.; Aarestrup, F.M. Indications and Patterns of Therapeutic Use of Antimicrobial Agents in the Danish Pig Production from 2002 to 2008. J. Vet. Pharmacol. Ther. 2012, 35, 33–46. [Google Scholar] [CrossRef]
- Lekagul, A.; Tangcharoensathien, V.; Mills, A.; Rushton, J.; Yeung, S. How Antibiotics Are Used in Pig Farming: A Mixed-Methods Study of Pig Farmers, Feed Mills and Veterinarians in Thailand. BMJ Glob. Health 2020, 5, e001918. [Google Scholar] [CrossRef] [Green Version]
- Barilli, E.; Vismarra, A.; Villa, Z.; Bonilauri, P.; Bacci, C. ESβL E. coli Isolated in Pig’s Chain: Genetic Analysis Associated to the Phenotype and Biofilm Synthesis Evaluation. Int. J. Food Microbiol. 2019, 289, 162–167. [Google Scholar] [CrossRef]
- Biasino, W.; De Zutter, L.; Garcia-Graells, C.; Uyttendaele, M.; Botteldoorn, N.; Gowda, T.; Van Damme, I. Quantification, Distribution and Diversity of ESBL/AmpC-Producing Escherichia coli on Freshly Slaughtered Pig Carcasses. Int. J. Food Microbiol. 2018, 281, 32–35. [Google Scholar] [CrossRef]
- Von Salviati, C.; Friese, A.; Roschanski, N.; Laube, H.; Guerra, B.; Käsbohrer, A.; Kreienbrock, L.; Roesler, U. Extended-Spectrum Beta-Lactamases (ESBL)/AmpC Beta-Lactamases-Producing Escherichia coli in German Fattening Pig Farms: A Longitudinal Study. Berl. Munch. Tierarztl. Wochenschr. 2014, 127, 412–419. [Google Scholar] [CrossRef]
- Peng, S.; Herrero-Fresno, A.; Olsen, J.E.; Dalsgaard, A. Influence of Zinc on CTX-M-1 β-Lactamase Expression in Escherichia coli. J. Glob. Antimicrob. Resist. 2020, 22, 613–619. [Google Scholar] [CrossRef]
- Cantón, R.; Morosini, M.I.; Martin, O.; de la Maza, S.; de la Pedrosa, G.E.G. IRT and CMT β-Lactamases and Inhibitor Resistance. Clin. Microbiol. Infect. 2008, 14, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Bradford, P.A. Interplay between β-Lactamases and New β-Lactamase Inhibitors. Nat. Rev. Microbiol. 2019, 17, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Naziri, Z.; Derakhshandeh, A.; Soltani Borchaloee, A.; Poormaleknia, M.; Azimzadeh, N. Treatment Failure in Urinary Tract Infections: A Warning Witness for Virulent Multi-Drug Resistant ESBL- Producing Escherichia coli. Infect. Drug Resist. 2020, 13, 1839–1850. [Google Scholar] [CrossRef]
- Livermore, D.M.; Mushtaq, S.; Nguyen, T.; Warner, M. Strategies to Overcome Extended-Spectrum β-Lactamases (ESBLs) and AmpC β-Lactamases in Shigellae. Int. J. Antimicrob. Agents 2011, 37, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum β-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedenic, B.; Vranes, J.; Suto, S.; Zagar, Z. Bactericidal Activity of Oral β-Lactam Antibiotics in Plasma and Urine versus Isogenic Escherichia coli Strains Producing Broad- and Extended-Spectrum β-Lactamases. Int. J. Antimicrob. Agents 2005, 25, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Ozgumus, O.B.; Tosun, I.; Aydin, F.; Kilic, A.O. Horizontol Dissemination of TEM- and SHV-Typr Beta-Lactamase Genes-Carrying Resistance Plasmids amongst Clonical Isolates of Enterobacteriaceae. Braz. J. Microbiol. 2008, 39, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Balkhed, Å.Ö.; Tärnberg, M.; Monstein, H.-J.; Hällgren, A.; Hanberger, H.; Nilsson, L.E. High Frequency of Co-Resistance in CTX-M-Producing Escherichia coli to Non-Beta-Lactam Antibiotics, with the Exceptions of Amikacin, Nitrofurantoin, Colistin, Tigecycline, and Fosfomycin, in a County of Sweden. Scand. J. Infect. Dis. 2013, 45, 271–278. [Google Scholar] [CrossRef]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef] [Green Version]
- Salah, F.D.; Soubeiga, S.T.; Ouattara, A.K.; Sadji, A.Y.; Metuor-Dabire, A.; Obiri-Yeboah, D.; Banla-Kere, A.; Karou, S.; Simpore, J. Distribution of Quinolone Resistance Gene (Qnr) in ESBL-Producing Escherichia coli and Klebsiella spp. in Lomé, Togo. Antimicrob. Resist. Infect. Control 2019, 8, 104. [Google Scholar] [CrossRef]
- Kim, J.O.; Yoo, I.Y.; Yu, J.K.; Kwon, J.A.; Kim, S.Y.; Park, Y.-J. Predominance and Clonal Spread of CTX-M-15 in Cefotaxime-Resistant Klebsiella Pneumoniae in Korea and Their Association with Plasmid-Mediated Quinolone Resistance Determinants. J. Infect. Chemother. 2021, 27, 1186–1192. [Google Scholar] [CrossRef]
- Lalak, A.; Wasyl, D.; Zając, M.; Skarżyńska, M.; Hoszowski, A.; Samcik, I.; Woźniakowski, G.; Szulowski, K. Mechanisms of Cephalosporin Resistance in Indicator Escherichia coli Isolated from Food Animals. Vet. Microbiol. 2016, 194, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Blanc, D.S.; Poncet, F.; Grandbastien, B.; Greub, G.; Senn, L.; Nordmann, P. Evaluation of the Performance of Rapid Tests for Screening Carriers of Acquired ESBL-Producing Enterobacterales and Their Impact on Turnaround Time. J. Hosp. Infect. 2021, 108, 19–24. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance; Version 2.0. 2017. Available online: https://www.eucast.org/resistance_mechanisms/ (accessed on 10 September 2021).
- CLSI. Performance Standards for Antimicrobial & Antifungal Susceptibility Testing. Clinical and Laboratory Standards Institute, Wayne, USA, M100 ED28:2018. 2018. Available online: https://clsi.org/about/about-clsi/about-clsi-antimicrobial-and-antifungal-susceptibility-testing-resources/ (accessed on 10 September 2021).
- EUCAST. EUCAST Disk Diffusion Method for Antimicrobial Susceptibility Testing, Version 6.0. 2017. Available online: https://kaldur.landspitali.is/focal/gaedahandbaekur/gnhsykla.nsf/5e27f2e5a88c898e00256500003c98c2/795d21eac4c71fdf00257ac30056d723/$FILE/Manual_v_6.0_EUCAST_Disk_Test_final.pdf (accessed on 10 September 2021).
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 9.0. 2019. Available online: http://www.Eucast.org (accessed on 14 June 2019).
- Jahanbakhsh, S.; Smith, M.G.; Kohan-Ghadr, H.-R.; Letellier, A.; Abraham, S.; Trott, D.J.; Fairbrother, J.M. Dynamics of Extended-Spectrum Cephalosporin Resistance in Pathogenic Escherichia coli Isolated from Diseased Pigs in Quebec, Canada. Int. J. Antimicrob. Agents 2016, 48, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Kaftandzieva, A.; Trajkovska-Dokic, E.; Panovski, N. Prevalence and Molecular Characterization of Extended Spectrum Beta-Lactamases (ESBLs) Producing Escherichia coli and Klebsiella Pneumoniae. Prilozi 2011, 32, 129–141. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Site | Positive | |
|---|---|---|
| Presence of ESBL-producing E. coli on large and small pig farms, (%) | ||
| Large farms (n = 465) | L1 (n = 115) | 74 (64.3) c |
| L2 (n = 127) | 38 (29.9) b | |
| L3 (n = 121) | 13 (10.7) a | |
| L4 (n = 102) | 0 | |
| Small farms (n = 150) | S1 (n = 40) | 19 (47.5) c |
| S3 (n = 41) | 0 | |
| S2 (n = 69) | 0 | |
| Presence of ESBL-producing E. coli in pigs of different age, (%) | ||
| Weeks of age (n = 538) | 4 (n = 110) | 31 (28.2) b |
| 6 (n = 151) | 53 (35.1) b | |
| 8 (n = 105) | 11 (10.5) a | |
| 12 (n = 87) | 10 (11.5) a | |
| 20 (n = 85) | 7 (8.2) a | |
| Sows (n = 77) | 32 (41.6) b | |
| Patterns of Resistance to Beta-lactams | Number of Beta-lactamase Gene(s) | ||||||
|---|---|---|---|---|---|---|---|
| blaCTX-M | blaSHV | blaTEM | blaTEM+ blaSHV | blaTEM+ blaCTX-M | blaTEM+ blaSHV+ blaCTX-M | Total | |
| Large farms (L1, L2, L3) | |||||||
| AM-TIC-AMC-CL-CZ-CAZ-CTX-CFM-FEP | 0 | 0 | 0 | 0 | 1 | 1 | 2 |
| AM-TIC-AMC-CL-CZ-CTX-CFM-FEP | 1 | 0 | 1 | 0 | 6 | 9 | 17 |
| AM-TIC-AMC-CL-CZ-CTX-FEP | 1 | 0 | 0 | 0 | 8 | 4 | 13 |
| AM-TIC-AMC-CL-CZ-FOX-CTX-CFM-FEP | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| AM-TIC-CL-CZ-CTX-CFM-FEP | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| AM-TIC-CL-CZ-CTX-FEP | 0 | 0 | 0 | 0 | 4 | 0 | 4 |
| Small farm (S1) | |||||||
| AM-TIC-AMC-CZ-CAZ-CTX-CFM | 0 | 0 | 0 | 1 | 0 | 3 | 4 |
| AM-TIC-AMC-CZ-CAZ-CTX-CFM-FEP | 0 | 0 | 0 | 4 | 1 | 1 | 6 |
| AM-TIC-CZ | 0 | 0 | 1 | 0 | 0 | 1 | 2 |
| Total | 3 | 0 | 2 | 5 | 21 | 19 | 50 |
| Selected Beta-Lactamase Gene | Antibiotic | OR | 95% CI | p-Value |
|---|---|---|---|---|
| blaSHV | AMC | 6.79 | 1.39–66.10 | 0.0083 |
| CL | 0.12 | 0.03–0.41 | 0.0001 | |
| CAZ | 8.38 | 2.47–37.17 | 0.0001 | |
| CFM | 4.36 | 1.70–12.00 | 0.0009 | |
| FEP | 0.15 | 0.01–0.79 | 0.0124 | |
| SXT | 0.30 | 0.12–0.77 | 0.0067 | |
| TMP | 0.30 | 0.12–0.77 | 0.0067 | |
| blaCTX-M | CL | 34.86 | 6.63–357.80 | 0.0001 |
| CTX | 6.77 | 0.45–101.68 | 0.0931 | |
| CAZ | 0.08 | 0.01–0.33 | 0.0001 | |
| SXT | 15.03 | 3.02–148.12 | 0.0001 | |
| TMP | 15.03 | 3.02–148.12 | 0.0001 | |
| GM | inf | 2.90–inf | 0.0003 | |
| TE | 12.11 | 2.45–118.82 | 0.0003 | |
| ENO | inf | 1.46–inf | 0.0093 |
| Farm | Total Number of Sows | Time to Weaning (Days) | Diarrhea/Critical Periods | Feed Additives | Antibiotics Used | Immunisation against E. coli b | ||
|---|---|---|---|---|---|---|---|---|
| Penicillin/Board-Spectrum Penicillins | Cephalo-Sporins | Non-Beta Lactams | ||||||
| L1 | 2100 | 28 | common/ 1–3 days; 4–6 weeks; 7–8 weeks | ZnO–150 mg kg−1 | AMX | CFT | CS, ENO, MAR, OT, SXT a | yes |
| L2 | 700 | 33 | common/ 2–3 weeks; 5–6 weeks | ZnO–150 mg kg−1; during weaning–zinc and electrolyte agent c | P, AMX | no | GM, OT, TMP + S, | yes |
| L3 | 1700 | 28 | sporadically | ZnO–150 mg kg−1; starter phase (2–3 weeks after weaning)–ZnO 2500 mg kg−1; water acidification d | no | CFT, CFQ | CS, ENO, GM, OT, TMP + S, | yes |
| L4 | 1000 | 28 | sporadically | ZnO–150 mg kg−1; | P, AMC | no | OT TMP + S, | no |
| S1 | 15 | 28 | common/ 5–6 weeks | - | P, AMX | no | ENO, OT, TMP + S | yes |
| S2 | 40 | 28–42 | rare | - | no | no | TY | no |
| S3 | 20 | 40 | rare | - | no | no | TM | no |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gāliņa, D.; Balins, A.; Valdovska, A. The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia. Antibiotics 2021, 10, 1099. https://doi.org/10.3390/antibiotics10091099
Gāliņa D, Balins A, Valdovska A. The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia. Antibiotics. 2021; 10(9):1099. https://doi.org/10.3390/antibiotics10091099
Chicago/Turabian StyleGāliņa, Daiga, Andris Balins, and Anda Valdovska. 2021. "The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia" Antibiotics 10, no. 9: 1099. https://doi.org/10.3390/antibiotics10091099
APA StyleGāliņa, D., Balins, A., & Valdovska, A. (2021). The Prevalence and Characterization of Fecal Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Isolated from Pigs on Farms of Different Sizes in Latvia. Antibiotics, 10(9), 1099. https://doi.org/10.3390/antibiotics10091099