
Antimicrobial Peptides and Their Applications in Biomedical Sector



Abstract
1. Introduction
2. Factors Affecting the Functioning of AMPs
2.1. Secondary Structure
2.2. Charge
2.3. Hydrophobicity
2.4. Amphipathicity
2.5. Hydrophobic Moment
2.6. Polar Angle
2.7. Peptide Length
3. Functions of AMPs
3.1. Disruption of Bacteria
3.2. Antifungal Activity
3.3. Antiviral Activity
3.4. Inhibition of Cancer Cell Growth
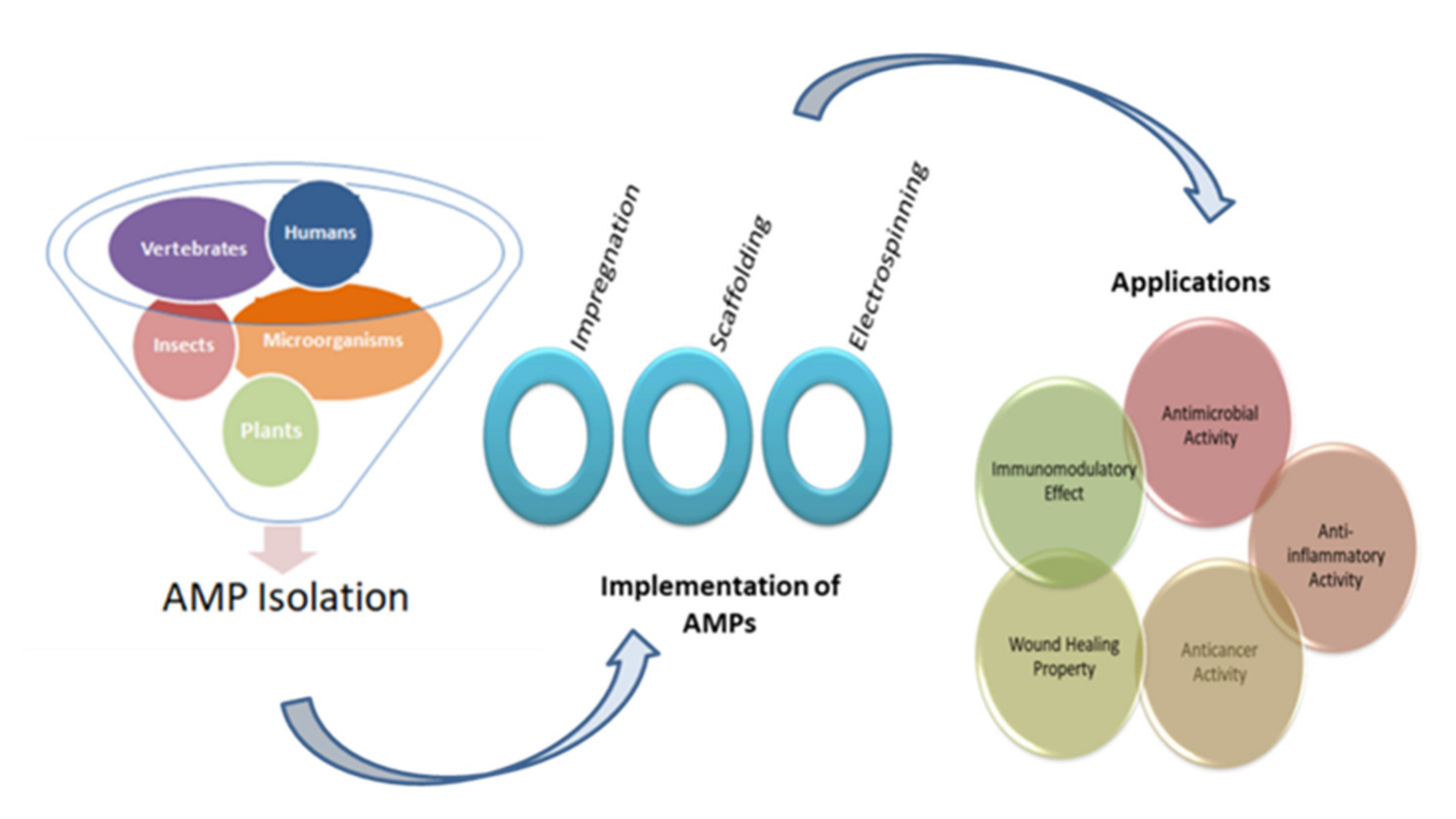
3.5. Immunomodulatory Effect
3.6. Anti-Inflammatory Effect
3.7. Wound Healing
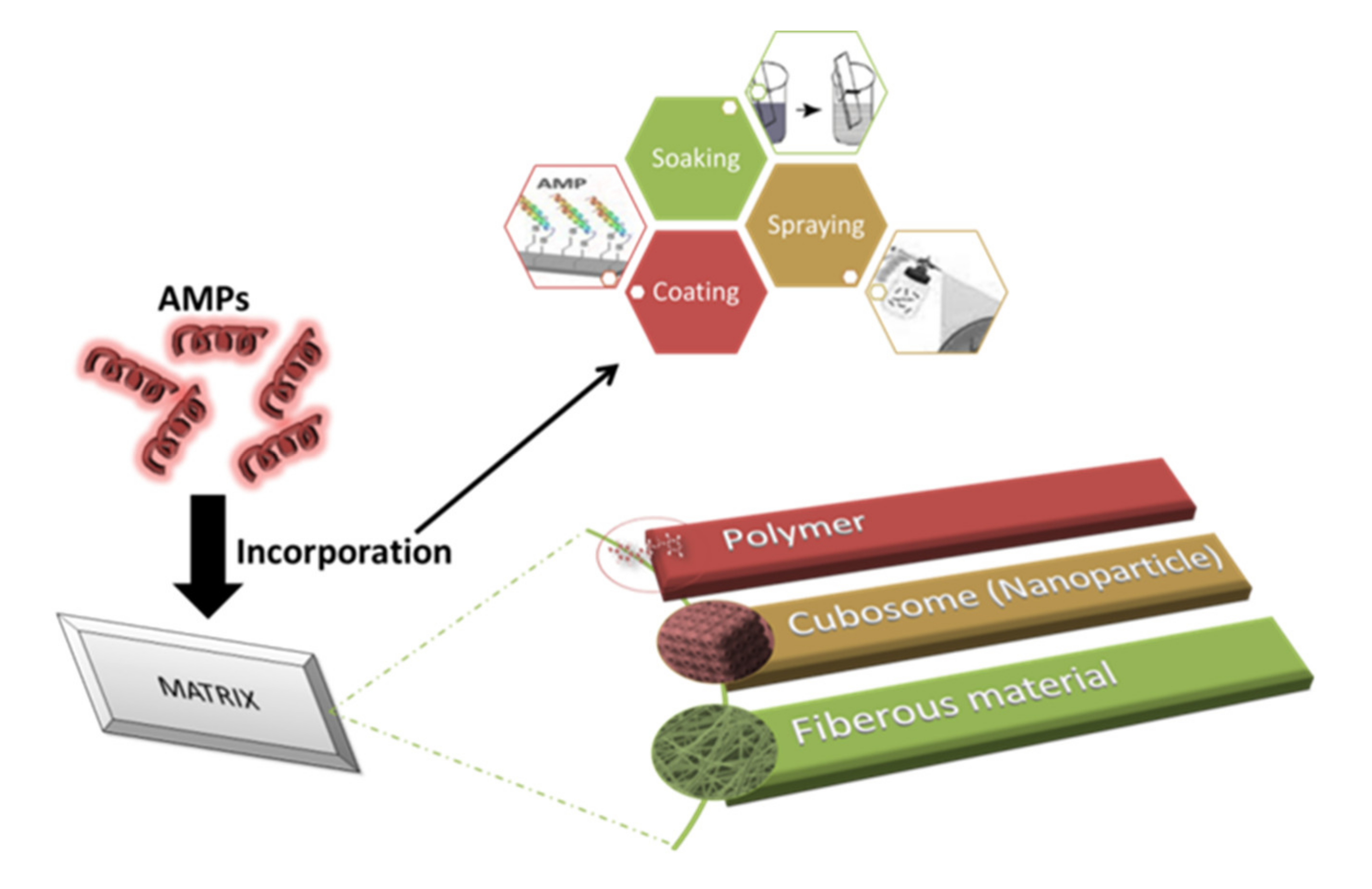
4. Implementation Techniques
4.1. Impregnation of AMPs
4.2. Scaffolding
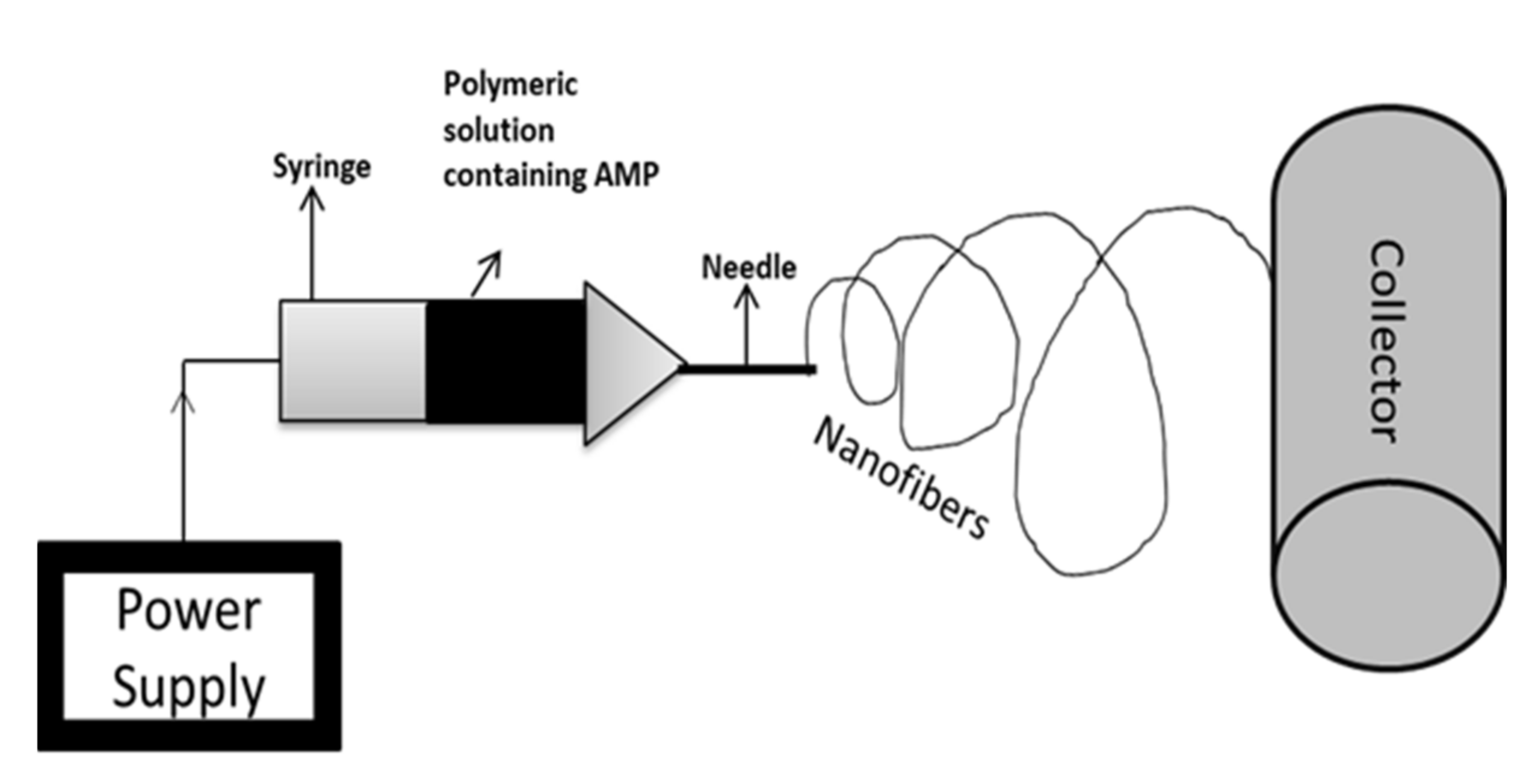
4.3. Electrospinning
5. Applications in Biomedical Sector
5.1. Vertebrate-Derived Antimicrobial Peptides
5.2. Insect Derived AMPs
5.3. Plant Derived AMPs
5.4. Microorganism Derived AMPs
6. Smart and Intelligent Delivery of AMPs
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, A.; Gupta, R.; Singh, R.L. Microbes and environment. In Principles and Applications of Environmental Biotechnology for a Sustainable Future; Springer: Singapore, 2017; pp. 43–84. [Google Scholar]
- Leekha, S.; Terrell, C.L.; Edson, R.S. General principles of antimicrobial therapy. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2011; Volume 86, pp. 156–167. [Google Scholar]
- Grace, J.L.; Huang, J.X.; Cheah, S.E.; Truong, N.P.; Cooper, M.A.; Li, J.; Davis, T.P.; Quinn, J.F.; Velkov, T.; Whittaker, M.R. Antibacterial low molecular weight cationic polymers: Dissecting the contribution of hydrophobicity, chain length and charge to activity. RSC Adv. 2016, 6, 15469–15477. [Google Scholar] [CrossRef]
- Ahmed, M.S. Synthesis of Antimicrobial Polymers to Overcome Antimicrobial Resistance. Ph.D. Thesis, Florida International University, Miami, FL, USA, 12 June 2017. [Google Scholar]
- Willing, B.P.; Russell, S.L.; Finlay, B.B. Shifting the balance: Antibiotic effects on host–microbiota mutualism. Nat. Rev. Microbiol. 2011, 9, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, e24122. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Mondal, N.R.; Jagdale, D.M. AntimicrobialPeptides A Review on its Types, Mechanism of Action, Synthesis and Therapeutic Applications. Int. J. For. Pharm. Res. Sch. 2016, 5, 72–81. [Google Scholar]
- Hultmark, D. Drosophila immunity: Paths and patterns. Curr. Opin. Immunol. 2003, 15, 12–19. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Almarwani, B.; Phambu, N.; Hamada, Y.Z.; Sunda-Meya, A. Interactions of an Anionic Antimicrobial Peptide with Zinc (II): Application to Bacterial Mimetic Membranes. Langmuir 2020, 36, 14554–14562. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural determinants of host defense peptides for antimicrobial activity and target cell selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. DRAMP 2.0, an updated data repository of antimicrobial peptides. Sci. Data 2019, 6, 1–10. [Google Scholar] [CrossRef]
- Reddy, K.; Yedery, R.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect Immunity. Purification and Properties of Three Inducible Bactericidal Proteins from Hemolymph of Immunized Pupae of Hyalophora cecropia. Eur. J. Biochem. 2005, 106, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, J.G. Phagocytin: A bactericidal substance from polymorphonuclear leucocytes. J. Exp. Med. 1956, 103, 589–611. [Google Scholar] [CrossRef] [PubMed]
- De Caleya, R.F.; Gonzalez-Pascual, B.; Garcia-Olmedo, F.; Carbonero, P. Susceptibility of phytopathogenic bacteria to wheat purothionins in vitro. Appl. Microbiol. 1972, 23, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.D.; Dubos, R.J. The Isolation of Bactericidal Substances from Cultures of Bacillus Brevis. J. Biol. Chem. 1941, 141, 155–162. [Google Scholar] [CrossRef]
- Ohtsuka, Y.; Inagaki, H. In silico identification and functional validation of linear cationic α-helical antimicrobial peptides in the ascidian Ciona intestinalis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.; Egorov, T. Plant antimicrobial peptides. Plant. Signal. Pept. 2012, 16, 107–133. [Google Scholar] [CrossRef]
- Thomas, S.; Karnik, S.; Barai, R.; Jayaraman, V.K.; Idicula-Thomas, S. CAMP: A useful resource for research on antimicrobial peptides. Nucleic Acids Res. 2009, 38, D774–D780. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M. Antibiotics set to flood Florida’s troubled orange orchards. Nature 2019, 567, 302–303. [Google Scholar] [CrossRef]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Meng, S.; Xu, H.; Wang, F. Research advances of antimicrobial peptides and applications in food industry and agriculture. Curr. Protein Pept. Sci. 2010, 11, 264–273. [Google Scholar] [CrossRef]
- Peschel, A.; Sahl, H.G. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat. Rev. Microbiol. 2006, 4, 529–536. [Google Scholar] [CrossRef]
- Yang, L.; Gordon, V.; Mishra, A.; Som, A.; Purdy, K.R.; Davis, M.A.; Tew, G.N.; Wong, G.C.L. Synthetic Antimicrobial Oligomers Induce a Composition-Dependent Topological Transition in Membranes. J. Am. Chem. Soc. 2007, 129, 12141–12147. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhou, F.; Su, H.; Zhang, Y. Structural design of microbicidal cationic oligomers and their synergistic interaction with azoles against Candida albicans. Sci. Rep. 2019, 9, 1–11. [Google Scholar]
- Palermo, E.F.; Kuroda, K. Structural determinants of antimicrobial activity in polymers which mimic host defense peptides. Appl. Microbiol. Biotechnol. 2010, 87, 1605–1615. [Google Scholar] [CrossRef]
- Li, P.; Li, X.; Saravanan, R.; Li, C.M.; Leong, S.S.J. Antimicrobial macromolecules: Synthesis methods and future applications. RSC Adv. 2012, 2, 4031–4044. [Google Scholar] [CrossRef]
- Waghu, F.H.; Gopi, L.; Barai, R.S.; Ramteke, P.; Nizami, B.; Idicula-Thomas, S. CAMP: Collection of sequences and structures of antimicrobial peptides. Nucleic Acids Res. 2013, 42, D1154–D1158. [Google Scholar] [CrossRef]
- Lee, T.-H.; Hall, K.N.; Aguilar, M.-I. Antimicrobial Peptide Structure and Mechanism of Action: A Focus on the Role of Membrane Structure. Curr. Top. Med. Chem. 2015, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Schmidtchen, A.; Pasupuleti, M.; Malmsten, M. Effect of hydrophobic modifications in antimicrobial peptides. Adv. Colloid Interface Sci. 2014, 205, 265–274. [Google Scholar] [CrossRef]
- McKay, M.J.; Afrose, F.; Koeppe II, R.E.; Greathouse, D.V. Helix formation and stability in membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2018, 1860, 2108–2117. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jackman, J.A.; Cho, N.J. Comparing the membrane-interaction profiles of two antiviral peptides: Insights into structure–function relationship. Langmuir 2019, 35, 9934–9943. [Google Scholar] [CrossRef]
- Sánchez-Vásquez, L.; Silva-Sanchez, J.; Jiménez-Vargas, J.M.; Rodriguez-Romero, A.; Muñoz-Garay, C.; Rodriguez, M.D.C.; Gurrola, G.B.; Possani, L.D. Enhanced antimicrobial activity of novel synthetic peptides derived from vejovine and hadrurin. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.F.; Pires, Á.S.; Franco, O.L. CS-AMPPred: An updated SVM model for antimicrobial activity prediction in cysteine-stabilized peptides. PLoS ONE 2012, 7, e51444. [Google Scholar] [CrossRef]
- Dathe, M.; Nikolenko, H.; Meyer, J.; Beyermann, M.; Bienert, M. Optimization of the antimicrobial activity of magainin peptides by modification of charge. FEBS Lett. 2001, 501, 146–150. [Google Scholar] [CrossRef]
- Hall, K.; Lee, T.H.; Aguilar, M.I. The role of electrostatic interactions in the membrane binding of melittin. J. Mol. Recognit. 2011, 24, 108–118. [Google Scholar] [CrossRef]
- Sengupta, D.; Leontiadou, H.; Mark, A.; Marrink, S.-J. Toroidal pores formed by antimicrobial peptides show significant disorder. Biochim. et Biophys. Acta (BBA) Biomembr. 2008, 1778, 2308–2317. [Google Scholar] [CrossRef] [PubMed]
- Laverty, G.; Gorman, S.P.; Gilmore, B.F. The Potential of Antimicrobial Peptides as Biocides. Int. J. Mol. Sci. 2011, 12, 6566–6596. [Google Scholar] [CrossRef] [PubMed]
- Sowa-Jasiłek, A.; Zdybicka-Barabas, A.; Stączek, S.; Pawlikowska-Pawlęga, B.; Grygorczuk-Płaneta, K.; Skrzypiec, K.; Gruszecki, W.I.; Mak, P.; Cytryńska, M. Antifungal Activity of Anionic Defense Peptides: Insight into the Action of Galleria mellonella Anionic Peptide 2. Int. J. Mol. Sci. 2020, 21, 1912. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, D.; Langel, K.; Eriste, E.; Langel, U. Cell-Penetrating Peptides: Design, Synthesis, and Applications. ACS Nano 2014, 8, 1972–1994. [Google Scholar] [CrossRef]
- Futaki, S.; Ohashi, W.; Suzuki, T.; Niwa, M.; Tanaka, S.; Ueda, K.; Harashima, H.; Sugiura, Y. Stearylated Arginine-Rich Peptides: A New Class of Transfection Systems. Bioconjugate Chem. 2001, 12, 1005–1011. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.; Macdonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef]
- Brogden, N.K.; Brogden, K.A. Will new generations of modified antimicrobial peptides improve their potential as pharmaceuticals? Int. J. Antimicrob. Agents 2011, 38, 217–225. [Google Scholar] [CrossRef]
- Rathinakumar, R.; Walkenhorst, W.F.; Wimley, W.C. Broad-Spectrum Antimicrobial Peptides by Rational Combinatorial Design and High-Throughput Screening: The Importance of Interfacial Activity. J. Am. Chem. Soc. 2009, 131, 7609–7617. [Google Scholar] [CrossRef]
- Narayana, J.L.; Mishra, B.; Lushnikova, T.; Wu, Q.; Chhonker, Y.S.; Zhang, Y.; Zarena, D.; Salnikov, E.S.; Dang, X.; Wang, F.; et al. Two distinct amphipathic peptide antibiotics with systemic efficacy. Proc. Natl. Acad. Sci. USA 2020, 117, 19446–19454. [Google Scholar] [CrossRef]
- Eisenberg, D.; Weiss, R.M.; Terwilliger, T.; Wilcox, W. Hydrophobic moments and protein structure. Faraday Symp. Chem. Soc. 1982, 17, 109–120. [Google Scholar] [CrossRef]
- Porto, W.F.; do Vale Ferreira, K.C.; Ribeiro, S.M.; Franco, O.L. Sense the Moment: A highly sensitive antimicrobial activity predictor based on hydrophobic moment. bioRxiv 2020. [Google Scholar] [CrossRef]
- Han, H.M.; Gopal, R.; Park, Y. Design and membrane-disruption mechanism of charge-enriched AMPs exhibiting cell selectivity, high-salt resistance, and anti-biofilm properties. Amino Acids 2015, 48, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Ebenhan, T.; Gheysens, O.; Kruger, H.G.; Zeevaart, J.R.; Sathekge, M.M. Antimicrobial Peptides: Their Role as Infection-Selective Tracers for Molecular Imaging. BioMed Res. Int. 2014, 2014, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, N.; Matsuzaki, K. Polar Angle as a Determinant of Amphipathic α-Helix-Lipid Interactions: A Model Peptide Study. Biophys. J. 2000, 79, 2075–2083. [Google Scholar] [CrossRef]
- Marchand, A.; Augenstreich, J.; Loiseau, C.; Verdon, J.; Lecomte, S.; Berjeaud, J.M. Effect of amino acid substitution in the staphylococcal peptides warnericin RK and PSMα on their anti-Legionella and hemolytic activities. Mol. Cell. Biochem. 2015, 405, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Juba, M.L.; Porter, D.K.; Williams, E.H.; Rodriguez, C.A.; Barksdale, S.M.; Bishop, B.M. Helical cationic antimicrobial peptide length and its impact on membrane disruption. Biochim. et Biophys. Acta (BBA) Biomembr. 2015, 1848, 1081–1091. [Google Scholar] [CrossRef]
- Epand, R.; Vogel, H.J. Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta (BBA) Biomembr. 1999, 1462, 11–28. [Google Scholar] [CrossRef]
- Podda, E.; Benincasa, M.; Pacor, S.; Micali, F.; Mattiuzzo, M.; Gennaro, R.; Scocchi, M. Dual mode of action of Bac7, a proline-rich antibacterial peptide. Biochim. Biophys. Acta (BBA) Gen. Subj. 2006, 1760, 1732–1740. [Google Scholar] [CrossRef]
- Elhag, O.; Zhou, D.; Song, Q.; Soomro, A.A.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Screening, Expression, Purification and Functional Characterization of Novel Antimicrobial Peptide Genes from Hermetia illucens (L.). PLoS ONE 2017, 12, e0169582. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the treatment of multidrug-resistant pathogens: A critical review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet. Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Nation, R.L.; Li, J.; Cars, O.; Couet, W.; Dudley, M.N.; Kaye, K.S.; Mouton, J.W.; Paterson, D.; Tam, V.H.; Theuretzbacher, U.; et al. Framework for optimisation of the clinical use of colistin and polymyxin B: The Prato polymyxin consensus. Lancet Infect. Dis. 2015, 15, 225–234. [Google Scholar] [CrossRef]
- Hirsch, R.; Wiesner, J.; Marker, A.; Pfeifer, Y.; Bauer, A.; Hammann, E.P.; Vilcinskas, A. Profiling antimicrobial peptides from the medical maggot Lucilia sericata as potential antibiotics for MDR Gram-negative bacteria. J. Antimicrob. Chemother. 2018, 74, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Kerenga, B.K.; McKenna, J.; Harvey, P.J.; Quimbar, P.; Garcia-Ceron, D.; Lay, F.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K.; et al. Salt-Tolerant Antifungal and Antibacterial Activities of the Corn Defensin ZmD32. Front. Microbiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Shurko, J.F.; Galega, R.S.; Li, C.; Lee, G.C. Evaluation of LL-37 antimicrobial peptide derivatives alone and in combination with vancomycin against S. aureus. J. Antibiot. 2018, 71, 971–974. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 1–2. [Google Scholar]
- Fenner, A. Antimicrobial peptide derived from moths can eradicate UPEC biofilms and could offer a novel therapeutic option. Nat. Rev. Urol. 2020, 17, 191. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Kwok, S.H.; Humble, J.L.; Liang, Y.; Tang, S.W.; Tang, K.H.; Tse, M.K.; Lei, J.H.; Ramalingam, R.; Koohi-Moghadam, M.; et al. BING, a novel antimicrobial peptide isolated from Japanese medaka plasma, targets bacterial envelope stress response by suppressing cpxR expression. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Zaet, A.; Dartevelle, P.; Daouad, F.; Ehlinger, C.; Quilès, F.; Francius, G.; Boehler, C.; Bergthold, C.; Frisch, B.; Prévost, G.; et al. D-Cateslytin, a new antimicrobial peptide with therapeutic potential. Sci. Rep. 2017, 7, 15199. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.; Ribeiro, S.M.; Matos, C.O.; Pires, Á.S.; Fensterseifer, I.C.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In silico optimization of a guava antimicrobial peptide enables combinatorial exploration for peptide design. Nat. Commun. 2018, 9, 1–2. [Google Scholar] [CrossRef]
- Ma, B.; Fang, C.; Lu, L.; Wang, M.; Xue, X.; Zhou, Y.; Li, M.; Hu, Y.; Luo, X.; Hou, Z. The antimicrobial peptide thanatin disrupts the bacterial outer membrane and inactivates the NDM-1 metallo-β-lactamase. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef]
- Knappe, D.; Piantavigna, S.; Hansen, A.; Mechler, A.; Binas, A.; Nolte, O.; Martin, L.L.; Hoffmann, R. Oncocin (VDKPPYLPRPRPPRRIYNR-NH2): A novel antibacterial peptide optimized against gram-negative human pathogens. J. Med. Chem. 2010, 53, 5240–5247. [Google Scholar] [CrossRef]
- De Lucca, A.J.; Walsh, T.J. Antifungal peptides: Novel therapeutic compounds against emerging pathogens. Antimicrob. Agents Chemother. 1999, 43, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Contursi, P.; Vollaro, A.; Fusco, S.; Notomista, E.; Catania, M.R. Antifungal and anti-biofilm activity of the first cryptic antimicrobial peptide from an archaeal protein against Candida spp. clinical isolates. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Utkina, L.L.; Andreev, Y.A.; Rogozhin, E.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Oparin, P.B.; Vassilevski, A.A.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. Genes encoding 4-Cys antimicrobial peptides in wheat T riticum kiharae Dorof. et Migush.: Multimodular structural organization, instraspecific variability, distribution and role in defence. FEBS J. 2013, 280, 3594–3608. [Google Scholar] [CrossRef]
- Ochiai, A.; Ogawa, K.; Fukuda, M.; Ohori, M.; Kanaoka, T.; Tanaka, T.; Taniguchi, M.; Sagehashi, Y. Rice Defensin OsAFP1 is a New Drug Candidate against Human Pathogenic Fungi. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Luo, X.-L.; Li, J.-X.; Huang, H.-R.; Duan, J.-L.; Dai, R.-X.; Tao, R.-J.; Yang, L.; Hou, J.-Y.; Jia, X.-M.; Xu, J.-F. LL37 Inhibits Aspergillus fumigatus Infection via Directly Binding to the Fungus and Preventing Excessive Inflammation. Front. Immunol. 2019, 10, 283. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento Dias, J.; de Souza Silva, C.; de Araújo, A.R.; Souza, J.M.; Júnior, P.H.; Cabral, W.F.; da Silva, M.D.; Eaton, P.; de Almeida, J.R.; Nicola, A.M.; et al. Mechanisms of action of antimicrobial peptides ToAP2 and NDBP-5.7 against Candida albicans planktonic and biofilm cells. Sci. Rep. 2020, 10, 1–4. [Google Scholar]
- Velivelli, S.L.S.; Czymmek, K.J.; Li, H.; Shaw, J.B.; Buchko, G.W.; Shah, D.M. Antifungal symbiotic peptide NCR044 exhibits unique structure and multifaceted mechanisms of action that confer plant protection. Proc. Natl. Acad. Sci. USA 2020, 117, 16043–16054. [Google Scholar] [CrossRef]
- Nakamura, I.; Yoshimura, S.; Masaki, T.; Takase, S.; Ohsumi, K.; Hashimoto, M.; Furukawa, S.; Fujie, A. ASP2397: A novel antifungal agent produced by Acremonium persicinum MF-347833. J. Antibiot. 2016, 70, 45–51. [Google Scholar] [CrossRef]
- Colombo, M.; Masiero, S.; Rosa, S.; Caporali, E.; Toffolatti, S.L.; Mizzotti, C.; Tadini, L.; Rossi, F.; Pellegrino, S.; Musetti, R.; et al. NoPv1: A synthetic antimicrobial peptide aptamer targeting the causal agents of grapevine downy mildew and potato late blight. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.R.; Gross, T.; Becker, A.; Vilcinskas, A.; Rahnamaeian, M. The selective antifungal activity of Drosophila melanogaster metchnikowin reflects the species-dependent inhibition of succinate–coenzyme Q reductase. Sci. Rep. 2017, 7, 1–9. [Google Scholar]
- De Azevedo, P.O.; Mendonça, C.M.; Moreno, A.C.; Bueno, A.V.; de Almeida, S.R.; Seibert, L.; Converti, A.; Watanabe, I.S.; Gierus, M.; de Souza Oliveira, R.P. Antibacterial and antifungal activity of crude and freeze-dried bacteriocin-like inhibitory substance produced by Pediococcus pentosaceus. Sci. Rep. 2020, 10, 1–4. [Google Scholar]
- Gong, Z.; Karlsson, A.J. Translocation of cell-penetrating peptides into Candida fungal pathogens. Protein Sci. 2017, 26, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sun, J.; Xia, S.; Tian, X.; Cheserek, M.J.; Le, G. Mechanism of antifungal activity of antimicrobial peptide APP, a cell-penetrating peptide derivative, against Candida albicans: Intracellular DNA binding and cell cycle arrest. Appl. Microbiol. Biotechnol. 2016, 100, 3245–3253. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jia, F.; Dang, W.; Zhao, Y.; Zhu, R.; Sun, M.; Qiu, S.; An, X.; Ma, Z.; Zhu, Y.; et al. Antifungal effect and action mechanism of antimicrobial peptide polybia-CP. J. Pept. Sci. 2015, 22, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E. Mechanisms of viral emergence. Vet. Res. 2010, 41, 38. [Google Scholar] [CrossRef]
- Castel, G.; Chtéoui, M.; Heyd, B.; Tordo, N. Phage Display of Combinatorial Peptide Libraries: Application to Antiviral Research. Molecules 2011, 16, 3499–3518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Jiang, X.; Qiao, J.; Wang, Z.; Tong, A.; Yang, J.; Yang, S.; Yang, L. Antimicrobial peptide DP7 with potential activity against SARS coronavirus infections. Signal Transduct. Target. Ther. 2021, 6, 1–3. [Google Scholar]
- Zhang, Z.; Zhang, H.; Peng, T.; Li, D.; Xu, J. Melittin suppresses cathepsin S-induced invasion and angiogenesis via blocking of the VEGF-A/VEGFR-2/MEK1/ERK1/2 pathway in human hepatocellular carcinoma. Oncol. Lett. 2016, 11, 610–618. [Google Scholar] [CrossRef][Green Version]
- Elnagdy, S.; AlKhazindar, M. The Potential of Antimicrobial Peptides as an Antiviral Therapy against COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 780–782. [Google Scholar] [CrossRef]
- Wiens, M.E.; Smith, J.G. α-Defensin HD5 Inhibits Human Papillomavirus 16 Infection via Capsid Stabilization and Redirection to the Lysosome. mBio 2017, 8, e02304-16. [Google Scholar] [CrossRef]
- Mohan, K.V.; Rao, S.S.; Atreya, C.D. Antiviral activity of selected antimicrobial peptides against vaccinia virus. Antivir. Res. 2010, 86, 306–311. [Google Scholar] [CrossRef]
- Nigro, E.; Colavita, I.; Sarnataro, D.; Scudiero, O.; Zambrano, G.; Granata, V.; Daniele, A.; Carotenuto, A.; Galdiero, S.; Folliero, V.; et al. An ancestral host defence peptide within human β-defensin 3 recapitulates the antibacterial and antiviral activity of the full-length molecule. Sci. Rep. 2015, 5, 18450. [Google Scholar] [CrossRef]
- Methatham, T.; Boonchuen, P.; Jaree, P.; Tassanakajon, A.; Somboonwiwat, K. Antiviral action of the antimicrobial peptide ALFPm3 from Penaeus monodon against white spot syndrome virus. Dev. Comp. Immunol. 2017, 69, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.M.; Oliveira, M.D.; Dias, R.S.; Nacif-Marçal, L.; Feio, R.N.; Ferreira, S.O.; Oliveira, L.L.; Silva, C.C.; Paula, S.O. The antimicrobial peptide HS-1 inhibits dengue virus infection. Virology 2018, 514, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Novoa, B.; Romero, A.; Álvarez, Á.L.; Moreira, R.; Pereiro, P.; Costa, M.M.; Dios, S.; Estepa, A.; Parra, F.; Figueras, A. Antiviral activity of myticin C peptide from mussel: An ancient defense against herpesviruses. J. Virol. 2016, 90, 7692–7702. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, J.; Zhang, K.; Chu, H.; Liu, D.; Poon, V.K.-M.; Chan, C.C.-S.; Leung, H.-C.; Fai, N.; Lin, Y.-P.; et al. A novel peptide with potent and broad-spectrum antiviral activities against multiple respiratory viruses. Sci. Rep. 2016, 6, 22008. [Google Scholar] [CrossRef] [PubMed]
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer 2012, 12, 237–251. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, Y.; Gao, S.; Cui, Y.; He, D.; Wang, L.; Chen, Y. Comparison on effect of hydrophobicity on the antibacterial and antifungal activities of α-helical antimicrobial peptides. Sci. China Chem. 2013, 56, 1307–1314. [Google Scholar] [CrossRef]
- Hu, C.; Chen, X.; Huang, Y.; Chen, Y. Co-administration of iRGD with peptide HPRP-A1 to improve anticancer activity and membrane penetrability. Sci. Rep. 2018, 8, 2274. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.E.F.; Najas, J.Z.G.; Magalhães, L.G.; Bobey, A.F.; Mendonça, J.N.; Lopes, N.P.; Leme, F.; Teixeira, S.P.; Trovó, M.; Andricopulo, A.D.; et al. Inhibition of Breast Cancer Cell Migration by Cyclotides Isolated from Pombalia calceolaria. J. Nat. Prod. 2018, 81, 1203–1208. [Google Scholar] [CrossRef]
- Rai, D.; Qian, S. Interaction of the Antimicrobial Peptide Aurein 1.2 and Charged Lipid Bilayer. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, X.; Li, Y.; Lan, X.; Leung, P.; Li, J.; Li, G.; Xie, M.; Han, Y.; Lin, X. Composite Membranes of Recombinant Silkworm Antimicrobial Peptide and Poly (L-lactic Acid) (PLLA) for biomedical application. Sci. Rep. 2016, 6, 31149. [Google Scholar] [CrossRef]
- Baindara, P.; Gautam, A.; Raghava, G.; Korpole, S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci. Rep. 2017, 7, srep46541. [Google Scholar] [CrossRef]
- Shi, D.; Hou, X.; Wang, L.; Gao, Y.; Wu, D.; Xi, X.; Zhou, M.; Kwok, H.F.; Duan, J.; Chen, T.; et al. Two Novel Dermaseptin-Like Antimicrobial Peptides with Anticancer Activities from the Skin Secretion of Pachymedusa dacnicolor. Toxins 2016, 8, 144. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, I.-W.; Kim, S.-H.; Kim, M.-A.; Yun, E.-Y.; Nam, S.-H.; Ahn, M.-Y.; Kang, D.; Hwang, J.S. Anticancer Activity of the Antimicrobial Peptide Scolopendrasin VII Derived from the Centipede, Scolopendra subspinipes mutilans. J. Microbiol. Biotechnol. 2015, 25, 1275–1280. [Google Scholar] [CrossRef]
- Li, C.; Liu, H.; Yang, Y.; Xu, X.; Lv, T.; Zhang, H.; Liu, K.; Zhang, S.; Chen, Y. N-myristoylation of Antimicrobial Peptide CM4 Enhances Its Anticancer Activity by Interacting with Cell Membrane and Targeting Mitochondria in Breast Cancer Cells. Front. Pharmacol. 2018, 9, 1297. [Google Scholar] [CrossRef]
- Chu, H.-L.; Yip, B.-S.; Chen, K.-H.; Yu, H.-Y.; Chih, Y.-H.; Cheng, H.-T.; Chou, Y.-T.; Cheng, J.-W. Novel Antimicrobial Peptides with High Anticancer Activity and Selectivity. PLoS ONE 2015, 10, e0126390. [Google Scholar] [CrossRef]
- Gaglione, R.; Pirone, L.; Farina, B.; Fusco, S.; Smaldone, G.; Aulitto, M.; Dell’Olmo, E.; Roscetto, E.; Del Gatto, A.; Fattorusso, R.; et al. Insights into the anticancer properties of the first antimicrobial peptide from Archaea. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 2155–2164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, I.-W.; Kim, S.-H.; Yun, E.-Y.; Nam, S.-H.; Ahn, M.-Y.; Kang, D.-C.; Hwang, J.S. Anticancer activity of CopA3 dimer peptide in human gastric cancer cells. BMB Rep. 2015, 48, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Cui, Z.; Li, Y.-H.; Hsu, W.-H.; Lee, B.-H. In Vitro and in Vivo Anticancer Activity of Pardaxin against Proliferation and Growth of Oral Squamous Cell Carcinoma. Mar. Drugs 2015, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Reffuveille, F.; De La Fuente-Núñez, C.; Mansour, S.; Hancock, R.E.W. A Broad-Spectrum Antibiofilm Peptide Enhances Antibiotic Action against Bacterial Biofilms. Antimicrob. Agents Chemother. 2014, 58, 5363–5371. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hilchie, A.L.; de la Fuente-Núñez, C.; Hancock, R.E. High throughput screening methods for assessing antibiofilm and immunomodulatory activities of synthetic peptides. Peptides 2015, 71, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.C.; De La Fuente-Núñez, C.; Hancock, R. Peptide IDR-1018: Modulating the immune system and targeting bacterial biofilms to treat antibiotic-resistant bacterial infections. J. Pept. Sci. 2014, 21, 323–329. [Google Scholar] [CrossRef]
- Nijnik, A.; Hancock, R.E.W. The roles of cathelicidin LL-37 in immune defences and novel clinical applications. Curr. Opin. Hematol. 2009, 16, 41–47. [Google Scholar] [CrossRef]
- Silva, O.; De La Fuente-Núñez, C.; Haney, E.F.; Fensterseifer, I.C.M.; Ribeiro, S.; Porto, W.; Brown, P.; Faria-Junior, C.; Rezende, T.; Moreno, S.E.; et al. An anti-infective synthetic peptide with dual antimicrobial and immunomodulatory activities. Sci. Rep. 2016, 6, 35465. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; Brezden, A.; Mohammad, H.; Chmielewski, J.; Seleem, M. A short D-enantiomeric antimicrobial peptide with potent immunomodulatory and antibiofilm activity against multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Sci. Rep. 2017, 7, 6953. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of innate immunity by a bacterial secreted peptide: Lantibiotic nisin Z is selectively immunomodulatory. Innate Immun. 2012, 19, 315–327. [Google Scholar] [CrossRef]
- Mookherjee, N.; Brown, K.; Bowdish, D.; Doria, S.; Falsafi, R.; Hokamp, K.; Roche, F.M.; Mu, R.; Doho, G.H.; Pistolic, J.; et al. Modulation of the TLR-Mediated Inflammatory Response by the Endogenous Human Host Defense Peptide LL-37. J. Immunol. 2006, 176, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, J.A.E.; Scheenstra, M.R.; Tjeerdsma-van Bokhoven, L.M.J.; Coorens, M.; Schneider, A.F.V.; Bikker, F.J.; van Dijk, A.; Haagsman, H.P. Antimicrobial and immunomodulatory activity of PMAP-23 derived peptides. Protein Pept. Lett. 2017, 24, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mao, R.; Teng, D.; Hao, Y.; Chen, H.; Wang, X.; Wang, X.; Yang, N.; Wang, J. Antibacterial and immunomodulatory activities of insect defensins-DLP2 and DLP4 against multidrug-resistant Staphylococcus aureus. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Neshani, A.; Zare, H.; Eidgahi, M.R.A.; Khaledi, A.; Ghazvini, K. Epinecidin-1, a highly potent marine antimicrobial peptide with anticancer and immunomodulatory activities. BMC Pharmacol. Toxicol. 2019, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide cNK-2. Sci. Rep. 2017, 7, srep45099. [Google Scholar] [CrossRef]
- Ting, C.-H.; Pan, C.-Y.; Chen, Y.-C.; Lin, Y.-C.; Chen, T.-Y.; Rajanbabu, V.; Chen, J.-Y. Impact of Tilapia hepcidin 2-3 dietary supplementation on the gut microbiota profile and immunomodulation in the grouper (Epinephelus lanceolatus). Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Wang, X.; Chen, L.; Zhao, X.; Xiao, L.; Yi, S.; Kong, Y.; Jiang, Y.; Zhang, J. A cathelicidin-related antimicrobial peptide suppresses cardiac hypertrophy induced by pressure overload by regulating IGFR1/PI3K/AKT and TLR9/AMPKα. Cell Death Dis. 2020, 11, 96-15. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M. Antimicrobial peptide, cLF36, affects performance and intestinal morphology, microflora, junctional proteins, and immune cells in broilers challenged with E. coli. Sci. Rep. 2019, 9, 1–9. [Google Scholar]
- Ko, S.J.; Park, E.; Asandei, A.; Choi, J.-Y.; Lee, S.-C.; Seo, C.H.; Luchian, T.; Park, Y. Bee venom-derived antimicrobial peptide melectin has broad-spectrum potency, cell selectivity, and salt-resistant properties. Sci. Rep. 2020, 10, 1–12. [Google Scholar]
- Wei, L.; Huang, C.; Yang, H.; Li, M.; Yang, J.; Qiao, X.; Mu, L.; Xiong, F.; Wu, J.; Xu, W. A potent anti-inflammatory peptide from the salivary glands of horsefly. Parasites Vectors 2015, 8, 556. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brunetti, J.; Carnicelli, V.; Ponzi, A.; Di Giulio, A.; Lizzi, A.R.; Cristiano, L.; Cresti, L.; Cappello, G.; Pollini, S.; Mosconi, L.; et al. Antibacterial and Anti-Inflammatory Activity of an Antimicrobial Peptide Synthesized with D Amino Acids. Antibiotics 2020, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jacob, B.; Jang, M.; Kwak, C.; Lee, Y.; Son, K.; Lee, S.; Jung, I.D.; Jeong, M.S.; Kwon, S.-H.; et al. Development of a novel short 12-meric papiliocin-derived peptide that is effective against Gram-negative sepsis. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Leake, I. Cathelicidin can reverse intestinal fibrosis in models of colitis. Nat. Rev. Gastroenterol. Hepatol. 2014, 12, 3. [Google Scholar] [CrossRef]
- Qiu, X.; Macchietto, M.G.; Liu, X.; Lu, Y.; Ma, Y.; Guo, H.; Saqui-Salces, M.; Bernlohr, D.A.; Chen, C.; Shen, S.; et al. Identification of gut microbiota and microbial metabolites regulated by an antimicrobial peptide lipocalin 2 in high fat diet-induced obesity. Int. J. Obes. 2020, 45, 143–154. [Google Scholar] [CrossRef]
- Carlile, S.R.; Shiels, J.; Kerrigan, L.; Delaney, R.; Megaw, J.; Gilmore, B.F.; Weldon, S.; Dalton, J.P.; Taggart, C.C. Sea snake cathelicidin (Hc-cath) exerts a protective effect in mouse models of lung inflammation and infection. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Silva, J.P.; Dhall, S.; Garcia, M.; Chan, A.; Costa, C.; Gama, M.; Martins-Green, M. Improved burn wound healing by the antimicrobial peptide LLKKK18 released from conjugates with dextrin embedded in a carbopol gel. Acta Biomater. 2015, 26, 249–262. [Google Scholar] [CrossRef]
- Ramos, R.; Silva, J.P.; Rodrigues, A.C.; Costa, R.; Guardão, L.; Schmitt, F.; Soares, R.; Vilanova, M.; Domingues, L.; Gama, M. Wound healing activity of the human antimicrobial peptide LL37. Peptides 2011, 32, 1469–1476. [Google Scholar] [CrossRef]
- Song, D.W.; Kim, S.H.; Kim, H.H.; Lee, K.H.; Ki, C.S.; Park, Y.H. Multi-biofunction of antimicrobial peptide-immobilized silk fibroin nanofiber membrane: Implications for wound healing. Acta Biomater. 2016, 39, 146–155. [Google Scholar] [CrossRef]
- Jiao, J.; Peng, C.; Li, C.; Qi, Z.; Zhan, J.; Pan, S. Dual bio-active factors with adhesion function modified electrospun fibrous scaffold for skin wound and infections therapeutics. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Di Grazia, A.; Cappiello, F.; Imanishi, A.; Mastrofrancesco, A.; Picardo, M.; Paus, R.; Mangoni, M.L. The frog skin-derived antimicrobial peptide esculentin-1a (1-21) NH2 promotes the migration of human HaCaT keratinocytes in an EGF receptor-dependent manner: A novel promoter of human skin wound healing? PLoS ONE 2015, 10, e0128663. [Google Scholar]
- Bucekova, M.; Sojka, M.; Valachova, I.; Martinotti, S.; Ranzato, E.; Szep, Z.; Majtan, V.; Klaudiny, J.; Majtan, J. Bee-derived antibacterial peptide, defensin-1, promotes wound reepithelialisation in vitro and in vivo. Wound Heal. S. Afr. 2017, 10, 25–35. [Google Scholar]
- Chung, E.M.C.; Dean, S.N.; Propst, C.N.; Bishop, B.M.; Van Hoek, M.L. Komodo dragon-inspired synthetic peptide DRGN-1 promotes wound-healing of a mixed-biofilm infected wound. NPJ Biofilms Microbiomes 2017, 3, 1–13. [Google Scholar] [CrossRef]
- Tang, J.; Liu, H.; Gao, C.; Mu, L.; Yang, S.; Rong, M.; Zhang, Z.; Liu, J.; Ding, Q.; Lai, R. A Small Peptide with Potential Ability to Promote Wound Healing. PLoS ONE 2014, 9, e92082. [Google Scholar]
- Liu, S.; Long, Q.; Xu, Y.; Wang, J.; Xu, Z.; Wang, L.; Zhou, M.; Wu, Y.; Chen, T.; Shaw, C. Assessment of antimicrobial and wound healing effects of Brevinin-2Ta against the bacterium Klebsiella pneumoniae in dermally-wounded rats. Oncotarget 2017, 8, 111369–111385. [Google Scholar] [CrossRef]
- Etienne, O.; Picart, C.; Taddei, C.; Haikel, Y.; Dimarcq, J.L.; Schaaf, P.; Voegel, J.C.; Ogier, J.A.; Egles, C. Multilayer Polyelectrolyte Films Functionalized by Insertion of Defensin: A New Approach to Protection of Implants from Bacterial Colonization. Antimicrob. Agents Chemother. 2004, 48, 3662–3669. [Google Scholar] [CrossRef]
- Gomes, A.; Mano, J.; Queiroz, J.; Gouveia, I. Incorporation of antimicrobial peptides on functionalized cotton gauzes for medical applications. Carbohydr. Polym. 2015, 127, 451–461. [Google Scholar] [CrossRef]
- Webber, J.L.; Namivandi-Zangeneh, R.; Drozdek, S.; Wilk, K.A.; Boyer, C.; Wong, E.H.H.; Bradshaw-Hajek, B.H.; Krasowska, M.; Beattie, D.A. Incorporation and antimicrobial activity of nisin Z within carrageenan/chitosan multilayers. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Conn, C.; Drummond, C. Nanostructured bicontinuous cubic lipid self-assembly materials as matrices for protein encapsulation. Soft Matter 2013, 9, 3449–3464. [Google Scholar] [CrossRef]
- Meikle, T.; Zabara, A.; Waddington, L.J.; Separovic, F.; Drummond, C.J.; Conn, C.E. Incorporation of antimicrobial peptides in nanostructured lipid membrane mimetic bilayer cubosomes. Colloids Surf. B Biointerfaces 2017, 152, 143–151. [Google Scholar] [CrossRef]
- Rodríguez-Vázquez, M.; Vega-Ruiz, B.; Ramos-Zúñiga, R.; Saldaña-Koppel, D.A.; Quiñones-Olvera, L.F. Chitosan and its potential use as a scaffold for tissue engineering in regenerative medicine. BioMed Res. Int. 2015. [Google Scholar] [CrossRef]
- Lee, E.Y.; Zhang, C.; Di Domizio, J.; Jin, F.; Connell, W.; Hung, M.; Malkoff, N.; Veksler, V.; Gilliet, M.; Ren, P.; et al. Helical antimicrobial peptides assemble into protofibril scaffolds that present ordered dsDNA to TLR9. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhu, X.; Mutreja, I.; Boda, S.K.; Fischer, N.G.; Zhang, A.; Lui, C.; Qi, Y.; Aparicio, C. Biomimetic mineralized hybrid scaffolds with antimicrobial peptides. Bioact. Mater. 2021, 6, 2250–2260. [Google Scholar] [CrossRef] [PubMed]
- Rabanal, F.; Grau, A.; Vila-Farrés, X.; Gonzalez-Linares, J.; Borràs, M.; Vila, J.; Manresa, A.; Cajal, Y. A bioinspired peptide scaffold with high antibiotic activity and low in vivo toxicity. Sci. Rep. 2015, 5, 10558. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Raimi-Abraham, B.T.; Luo, C. Nanofibres in Drug Delivery; UCL Press: London, UK, 2018. [Google Scholar]
- Zare, M.; Dziemidowicz, K.; Williams, G.; Ramakrishna, S. Encapsulation of Pharmaceutical and Nutraceutical Active Ingredients Using Electrospinning Processes. Nanomaterials 2021, 11, 1968. [Google Scholar] [CrossRef]
- Kluin, J.; Talacua, H.; Smits, A.; Emmert, M.Y.; Brugmans, M.C.; Fioretta, E.; Dijkman, P.E.; Söntjens, S.; Duijvelshoff, R.; Dekker, S.; et al. In situ heart valve tissue engineering using a bioresorbable elastomeric implant—From material design to 12 months follow-up in sheep. Biomaterials 2017, 125, 101–117. [Google Scholar] [CrossRef]
- Riool, M.; De Breij, A.; Drijfhout, J.W.; Nibbering, P.H.; Zaat, S.A.J. Antimicrobial Peptides in Biomedical Device Manufacturing. Front. Chem. 2017, 5, 63. [Google Scholar] [CrossRef]
- López-Abarrategui, C.; Alba, A.; Lima, L.A.; Neto, S.M.; Vasconcelos, I.M.; Oliveira, J.T.A.; Dias, S.C.; Otero-Gonzalez, A.J.; Franco, O.L. Screening of Antimicrobials from Caribbean Sea Animals and Isolation of Bactericidal Proteins from the Littoral Mollusk Cenchritis muricatus. Curr. Microbiol. 2012, 64, 501–505. [Google Scholar] [CrossRef]
- Viana, J.F.C.; Carrijo, J.; Freitas, C.G.; Paul, A.; Alcaraz, J.; Lacorte, C.C.; Migliolo, L.; de Andrade, C.A.S.; Falcão, R.; Santos, N.; et al. Antifungal nanofibers made by controlled release of sea animal derived peptide. Nanoscale 2015, 7, 6238–6246. [Google Scholar] [CrossRef]
- Wang, X.; Yue, T.; Lee, T.-C. Development of Pleurocidin-poly(vinyl alcohol) electrospun antimicrobial nanofibers to retain antimicrobial activity in food system application. Food Control 2015, 54, 150–157. [Google Scholar] [CrossRef]
- Avila, E.E. Functions of Antimicrobial Peptides in Vertebrates. Curr. Protein Pept. Sci. 2017, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Selsted, M.E.; Lehrer, R.I. Defensins. Eur. J. Haematol. 1990, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Ouellette, A.J. Mammalian defensins in the antimicrobial immune response. Nat. Immunol. 2005, 6, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Lynn, D.J.; Bradley, D.G. Discovery of α-defensins in basal mammals. Dev. Comp. Immunol. 2007, 31, 963–967. [Google Scholar] [CrossRef]
- Selsted, M.E.; Tang, Y.Q.; Morris, W.L.; McGuire, P.A.; Novotny, M.J.; Smith, W.; Henschen, A.H.; Cullor, J.S. Purification, primary structures, and antibacterial activities of beta-defensins, a new family of antimicrobial peptides from bovine neutrophils. J. Biol. Chem. 1993, 268, 6641–6648. [Google Scholar] [CrossRef]
- Tran, D.; Tran, P.A.; Tang, Y.Q.; Yuan, J.; Cole, T.; Selsted, M.E. Homodimeric θ-Defensins from Rhesus macaqueLeukocytes: Isolation, synthesis, antimicrobial activities, and bacterial binding properties of the cyclic peptides. J. Biol. Chem. 2002, 277, 3079–3084. [Google Scholar] [CrossRef]
- Bals, R.; Wilson, J.M. Cathelicidins-a family of multifunctional antimicrobial peptides. Cell. Mol. Life Sci. CMLS 2003, 60, 711–720. [Google Scholar] [CrossRef]
- Zanetti, M.; Gennaro, R.; Romeo, D. Cathelicidins: A novel protein family with a common proregion and a variable C-terminal antimicrobial domain. FEBS Lett. 1995, 374, 1–5. [Google Scholar] [CrossRef]
- Agerberth, B.; Lee, J.-Y.; Bergman, T.; Carlquist, M.; Boman, H.G.; Mutt, V.; Jornvall, H. Amino acid sequence of PR-39. Isolation from pig intestine of a new member of the family of proline-arginine-rich antibacterial peptides. Eur. J. Biochem. 1991, 202, 849–854. [Google Scholar] [CrossRef]
- Gennaro, R.; Skerlavaj, B.; Romeo, D. Purification, composition, and activity of two bactenecins, antibacterial peptides of bovine neutrophils. Infect. Immun. 1989, 57, 3142–3146. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Human Antimicrobial Peptides and Proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [PubMed]
- Shaat, R.M.; El Meadawy, S.; Rizk, E.M.; Elgawad, M.S.A.; Elsaid, T.O. The significance of α-defensins 1-3 in Behcet’s disease: A case-control study among Egyptian patients. Egypt. Rheumatol. Rehabil. 2020, 47, 1–8. [Google Scholar] [CrossRef]
- Shukla, P.K.; Meena, A.S.; Rao, V.; Rao, R.G.; Balazs, L.; Rao, R. Human Defensin-5 Blocks Ethanol and Colitis-Induced Dysbiosis, Tight Junction Disruption and Inflammation in Mouse Intestine. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Taha-Abdelaziz, K.; Perez-Casal, J.; Schott, C.; Hsiao, J.; Attah-Poku, S.; Slavić, D.; Caswell, J.L. Bactericidal activity of tracheal antimicrobial peptide against respiratory pathogens of cattle. Vet. Immunol. Immunopathol. 2013, 152, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Coretti, L.; Natale, A.; Cuomo, M.; Florio, E.; Keller, S.; Lembo, F.; Chiariotti, L.; Pero, R. The Interplay between Defensins and Microbiota in Crohn’s Disease. Mediat. Inflamm. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, H.; Chen, R.; Zhou, J.; Xiao, Y.; Zhang, Y.; Yan, F. Human β-defensin 3 gene modification promotes the osteogenic differentiation of human periodontal ligament cells and bone repair in periodontitis. Int. J. Oral Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Cole, A.M.; Hong, T.; Boo, L.M.; Nguyen, T.; Zhao, C.; Bristol, G.; Zack, J.A.; Waring, A.J.; Yang, O.O.; Lehrer, R.I. Retrocyclin: A primate peptide that protects cells from infection by T- and M-tropic strains of HIV-1. Proc. Natl. Acad. Sci. USA 2002, 99, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Schaal, J.B.; Tran, D.; Tran, P.; Ösapay, G.; Trinh, K.; Roberts, K.D.; Brasky, K.M.; Tongaonkar, P.; Ouellette, A.J.; Selsted, M.E. Rhesus Macaque Theta Defensins Suppress Inflammatory Cytokines and Enhance Survival in Mouse Models of Bacteremic Sepsis. PLoS ONE 2012, 7, e51337. [Google Scholar] [CrossRef]
- YYasin, B.; Wang, W.; Pang, M.; Cheshenko, N.; Hong, T.; Waring, A.J.; Herold, B.C.; Wagar, E.A.; Lehrer, R.I. θ Defensins Protect Cells from Infection by Herpes Simplex Virus by Inhibiting Viral Adhesion and Entry. J. Virol. 2004, 78, 5147–5156. [Google Scholar] [CrossRef]
- Liang, Q.-L.; Zhou, K.; He, H.-X. Retrocyclin 2: A new therapy against avian influenza H5N1 virus in vivo and vitro. Biotechnol. Lett. 2009, 32, 387–392. [Google Scholar] [CrossRef]
- Rekha, R.S.; Muvva, S.S.V.J.R.; Wan, M.; Raqib, R.; Bergman, P.; Brighenti, S.; Gudmundsson, G.H.; Agerberth, B. Phenylbutyrate induces LL-37-dependent autophagy and intracellular killing of Mycobacterium tuberculosis in human macrophages. Autophagy 2015, 11, 1688–1699. [Google Scholar] [CrossRef]
- Van Eijk, M.; Boerefijn, S.; Cen, L.; Rosa, M.; Morren, M.J.; Van Der Ent, C.K.; Kraak, B.; Dijksterhuis, J.; Valdes, I.D.; Haagsman, H.P.; et al. Cathelicidin-inspired antimicrobial peptides as novel antifungal compounds. Med. Mycol. 2020, 58, 1073–1784. [Google Scholar] [CrossRef]
- Tonk, M.; Cabezas-Cruz, A.; Valdés, J.J.; Rego, R.O.; Chrudimská, T.; Strnad, M.; Šíma, R.; Bell-Sakyi, L.; Franta, Z.; Vilcinskas, A.; et al. Defensins from the tick Ixodes scapularis are effective against phytopathogenic fungi and the human bacterial pathogen Listeria grayi. Parasites Vectors 2014, 7, 1–8. [Google Scholar] [CrossRef]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol. 2019, 109, 31–42. [Google Scholar] [CrossRef]
- Tonk, M.; Vilcinskas, A.; Rahnamaeian, M. Insect antimicrobial peptides: Potential tools for the prevention of skin cancer. Appl. Microbiol. Biotechnol. 2016, 100, 7397–7405. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.R.; Kim, H.; Nam, S.H.; Yun, E.Y.; Kim, S.R.; Ahn, M.Y.; Chang, J.S.; Hwang, J.S. CopA3 peptide from Copris tripartitus induces apoptosis in human leukemia cells via a caspase-independent pathway. BMB Rep. 2012, 45, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, H.; Niu, L.; Wang, X. Efficient Screening of a Novel Antimicrobial Peptide from Jatropha curcas by Cell Membrane Affinity Chromatography. J. Agric. Food Chem. 2011, 59, 1145–1151. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 1–15. [Google Scholar] [CrossRef]
- Salimi, A.; Noorbakhsh, A.; Mamkhezri, H.; Ghavami, R. Electrocatalytic Reduction of H2O2 and Oxygen on the Surface of Thionin Incorporated onto MWCNTs Modified Glassy Carbon Electrode: Application to Glucose Detection. Electroanalysis 2007, 19, 1100–1108. [Google Scholar] [CrossRef]
- Zhang, D.; Li, W.; Wang, H.; Ma, Z. A novel immunoprobe composed of reduced graphene oxide-hemin-thionin-Au nanohybrid for ultrasensitive detection of tumor marker. Sens. Actuators B Chem. 2018, 258, 141-14. [Google Scholar] [CrossRef]
- Hu, E.; Wang, D.; Chen, J.; Tao, X. Novel cyclotides from Hedyotis diffusa induce apoptosis and inhibit proliferation and migration of prostate cancer cells. Int. J. Clin. Exp. Med. 2015, 8, 4059. [Google Scholar]
- Thery, T.; Arendt, E.K. Antifungal activity of synthetic cowpea defensin Cp-thionin II and its application in dough. Food MicroBiol. 2018, 73, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.; Kam, A.; Xiao, T.; Nguyen, G.K.T.; Liu, C.F.; Tam, J.P. Identification and Characterization of Roseltide, a Knottin-type Neutrophil Elastase Inhibitor Derived from Hibiscus sabdariffa. Sci. Rep. 2016, 6, 39401. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.T.; Velivelli, S.; Berg, R.H.; Oakley, B.; Shah, D.M. A novel bi-domain plant defensin MtDef5 with potent broad-spectrum antifungal activity binds to multiple phospholipids and forms oligomers. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Egan, K.; Field, D.; Rea, M.C.; Ross, R.; Hill, C.; Cotter, P.D. Bacteriocins: Novel Solutions to Age Old Spore-Related Problems? Front. Microbiol. 2016, 7, 461. [Google Scholar] [CrossRef]
- Lubelski, J.; Rink, R.; Khusainov, R.; Moll, G.N.; Kuipers, O.P. Biosynthesis, immunity, regulation, mode of action and engineering of the model lantibiotic nisin. Experientia 2007, 65, 455–476. [Google Scholar] [CrossRef]
- De Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Penna, T.C. Nisin biotechnological production and application: A review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP, a Bacteriocin and Food Preservative, Inhibits Head and Neck Cancer Tumorigenesis and Prolongs Survival. PLoS ONE 2015, 10, e0131008. [Google Scholar] [CrossRef]
- Preet, S.; Bharati, S.; Panjeta, A.; Tewari, R.; Rishi, P. Effect of nisin and doxorubicin on DMBA-induced skin carcinogenesis—A possible adjunct therapy. Tumor Biol. 2015, 36, 8301–8308. [Google Scholar] [CrossRef]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin-a bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef]
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA activities of enterocins DD28 and DD93 and evidences on their role in the inhibition of biofilm formation. Front. Microbiol. 2016, 7, 817. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC 1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Begley, M.; O’Connor, P.M.; Daly, K.M.; Hugenholtz, F.; Cotter, P.D.; Hill, C.; Ross, R. Bioengineered Nisin A Derivatives with Enhanced Activity against Both Gram Positive and Gram Negative Pathogens. PLoS ONE 2012, 7, e46884. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.M.; Ateia, I.; Paulus, J.R.; Liu, H.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Antimicrobial nisin acts against saliva derived multi-species biofilms without cytotoxicity to human oral cells. Front. Microbiol. 2015, 6, 617. [Google Scholar] [CrossRef] [PubMed]
- Tallet, L.; Gribova, V.; Ploux, L.; Vrana, N.E.; LaValle, P. New Smart Antimicrobial Hydrogels, Nanomaterials, and Coatings: Earlier Action, More Specific, Better Dosing? Adv. Healthc. Mater. 2020, 10, e2001199. [Google Scholar] [CrossRef]
- Alvarez-Lorenzo, C.; Concheiro, A. Intelligent drug delivery systems: Polymeric micelles and hydrogels. Mini Rev. Med. Chem. 2008, 8, 1065–1074. [Google Scholar] [CrossRef]
- Sousa, M.G.; Rezende, T.M.; Franco, O.L. Nanofibers as drug-delivery systems for antimicrobial peptides. Drug Discov. Today 2021, 26, 2064–2074. [Google Scholar] [CrossRef]
- Greber, K.E.; Dawgul, M. Antimicrobial peptides under clinical trials. Curr. Top. Med. Chem. 2017, 17, 620. [Google Scholar] [CrossRef]
- Azharuddin, M.; Zhu, G.H.; Das, D.; Ozgur, E.; Uzun, L.; Turner, A.P.F.; Patra, H.K. A repertoire of biomedical applications of noble metal nanoparticles. Chem. Commun. 2019, 55, 6964. [Google Scholar] [CrossRef]
- Kaur, J.; Gill, G.S.; Jeet, K. Applications of Carbon Nanotubes in Drug Delivery: A Comprehensive Review. In Characterization and Biology of Nanomaterials for Drug Delivery: Nanoscience and Nanotechnology in Drug Delivery, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2019; p. 133. [Google Scholar]
- Weiner, N.; Martin, F.; Riaz, M. Liposomes as a drug delivery system. Drug Dev. Ind. Pharm. 1989, 15, 1523. [Google Scholar] [CrossRef]
- Lancelot, A.; Sierra, T.; Serrano, J.L. Nanostructured liquid-crystalline particles for drug delivery. Expert Opin. Drug Deliv. 2014, 11, 547. [Google Scholar] [CrossRef]
- Sherje, A.P.; Jadhav, M.; Dravyakar, B.R.; Kadam, D. Dendrimers: A versatile nanocarrier for drug delivery and targeting. Int. J. Pharm. 2018, 548, 707. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; White, R.R. Aptamers for targeted drug delivery. Pharmaceuticals 2010, 3, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Hoare, T.R.; Kohane, D.S. Hydrogels in drug delivery: Progress and challenges. Polymers 2008, 49, 1993. [Google Scholar] [CrossRef]
- Joseph, B.; George, A.; Gopi, S.; Kalarikkal, N.; Thomas, S. Polymer sutures for simultaneous wound healing and drug delivery—A review. Int. J. Pharm. 2017, 524, 454. [Google Scholar] [CrossRef] [PubMed]
- Rajewski, R.A.; Stella, V.J. Pharmaceutical applications of cyclodextrins. 2. In vivo drug delivery. J. Pharm. Sci. 1996, 85, 1142. [Google Scholar]
- Matthews, J.A.; Wnek, G.E.; Simpson, D.G.; Bowlin, G.L. Electrospinning of Collagen Nanofibers. Biomacromolecules 2002, 3, 232–238. [Google Scholar] [CrossRef]
- Li, J.; Fan, C.; Pei, H.; Shi, J.; Huang, Q. Smart Drug Delivery Nanocarriers with Self-Assembled DNA Nanostructures. Adv. Mater. 2013, 25, 4386–4396. [Google Scholar] [CrossRef]
- Wang, Q.; Uzunoglu, E.; Wu, Y.; Libera, M. Self-Assembled Poly(ethylene glycol)-co-Acrylic Acid Microgels to Inhibit Bacterial Colonization of Synthetic Surfaces. ACS Appl. Mater. Interfaces 2012, 4, 2498–2506. [Google Scholar] [CrossRef] [PubMed]
- Bennet, D.; Kim, S. Polymer nanoparticles for smart drug delivery. In Application of Nanotechnology in Drug Delivery; InTech Open: London, UK, 2014. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Antibiotics | AMPs | References |
|---|---|---|---|
| Similarities | Destruction of microorganism | [28,29] | |
| Differences | No immunomodulatory effect observed | Has immunomodulatory effect | [30] |
| Microorganisms easily develop resistivity against antibiotics | No easy development of resistivity | ||
| No inflammatory response | Has effective anti-inflammatory activity | [14] | |
| Effective at low concentration | High concentration is required | [31] | |
| AMPs | Source | Bacteria Inhibited | Antimicrobial Activity | References |
|---|---|---|---|---|
| ZmD32 | Corn | E. coli, Bacillus subtilis, P. aeruginosa, and S. aureus | 50% Inhibitory concentration of ZmD32 ranged between 0.4 and 1.7 μM | [68] |
| LL-37 | Human cathelicidin hCAP18 | Methicillin-resistant Staphylococcus aureus, methicillin-susceptible S. aureus, Vancomycin Intermediate Staphylococcus aureus (VISA) and Vancomycin Resistant Staphylococcus aureus (VRSA) | Minimum inhibitory concentration was recorded as 64,128,64 and 256 µg/mL for VISA, MSSA, VRSA, and MRSA, respectively | [69] |
| Melimine and Mel4 | P. aeruginosa | It took 4 and 30 min for Mel4 and melimine, respectively, to permeate through the cytoplasmic layer | [70] | |
| Cecropin A | Moth | Uropathogenic E. coli (UPEC) | Incorporation of 0.25 µM−1 µM CecA with nalidixic acid was able to permeate through UPEC cell membrane by 15%. | [71] |
| BING | Japanese medaka plasma | Broad spectrum including E. coli, Enterococcus faecalis, S. aureus and P. aeruginosa A | Minimum inhibitory concentration of BONG ranged between 4 and 50 µg/mL | [72] |
| D-Cateslytin | Human | Methicillin-susceptible Staphylococcus aureus, Methicillin-resistant Staphylococcus aureus, Pseudomonas micra, Pseudomonas intermedia and F. nucleatum | Minimum inhibitory concentration of D-Cateslyt ranged between 8 and 24 μg/mL | [73] |
| Guavanin 2 | guava | E. coli, Listeria ivanovii and Candida parapsilosis | Minimum inhibitory concentration for E. coli, Listeria ivanovii, and C. parapsilosis was recorded as 6.25, 50, and 50 µM | [74] |
| Thanatin | E. coliandK. pneumoniae | Thanatin replaces divalent cations from bacterial membrane and causes disruption | [75] | |
| Temporin B | Frog skin | Staphylococcus epidermidis | Chitosan nanoparticles containing Temporin B showed 4 log reduction of S. epidermis compared to chitosan nanoparticles | [76] |
| Oncocin | Milkweed bug | P. aeruginosa, E. coli and Acinetobacter baumannii | Minimum inhibitory concentration of Oncocin was recorded as 0.125 to 8 μg/mL | [77] |
| AMPs | Source | Fungi Inhibited | Results | References |
|---|---|---|---|---|
| Tk-AMP-X1 and Tk-AMP-X2 | Triticum kiharae | Fusarium graminearum, Diplodia maydi and Fusarium verticillioides | 50% inhibition concentration of Tk-AMP-X1 and Tk-AMP-X2 range between 7.5 and 30 µg mL−1 | [80] |
| OsAFP1 | Rice | Candida Albicans | Inhibited C. albicans growth at 4µM concentration | [81] |
| LL-37 | Human | Aspergillus fumigatus | After 30 min of incubation, LL-37 binds to mycelia and damages the cell wall | [82] |
| oAP2 and NDBP-5.7 | Tityus obscurus and Opisthacanthus cayaporum scorpions | C. Albicans | MIC for Oap2 and NDBP-5.7 was recorded as 25 µM and 100 µM, respectively | [83] |
| NCR044 | Botrytis cinerea, Fusarium oxysporum, F. graminearum and Fusarium virguliforme | 50% of inhibition concentration ranged between 0.52 and 1.93 µM | [84] | |
| ASP2397 | Malaysian leaf litter | A. fumigatus | MIC was recorded as 0.78 µgmL−1 | [85] |
| NoPv1 | Synthetic | Plasmopara viticola | 200µM of NoPv1 showed the complete destruction of Plasmopara viticola | [86] |
| Metchnikowin | Drosophila melanogaster | F. graminearum | 50% inhibitory concentration of Metchnikowin was found as 1 µM | [87] |
| LBM 18 | Pediococcus pentosaceus | A. niger and Aspergillus flavus | Within 2 days of incubation BLIS was able to cause destruction to A. niger and Aspergillus flavus | [88] |
| Penetratin | Synthetic | C. albicans and C. glabrata | 50% inhibitory concentration was recorded in a range of 1 to 50 µM | [89] |
| APP | ppTG20 | Saccharomyces cerevisiae, C. albicans, A. niger, Trichopyton rubrum, A. flavus and Cryptococcus neoformans | MIC was recorded as 8, 16, and 32 µM for C. albicans and A. flavus, Saccharomyces cerevisiae, and Cryptococcus neoformans and A. niger and Trichopyton rubrum, respectively | [90] |
| polybia-CP | Polybia paulista | Candida strains | MIC of antimicrobial peptide was recorded in a range between 4 and 64 µM | [91] |
| AMPs | Source | Virus Inhibited | References |
|---|---|---|---|
| Melittin | Apis mellifera | Suppress the activation of cathepsin S | [95] |
| Lactoferrin | Mucosal secretions | SARS-CoV | [96] |
| HD-5 | Human | Human papillomavirus | [97] |
| PD3, PD4, and RW3 | Thrombin-induced human platelet and synthetic repeats of arginine-tryptophan | Vaccinia virus | [98] |
| Human β-defensin 3 | Human | human immunodeficiency virus and herpes simplex virus | [99] |
| ALFPm3 | Penaeus monodon | white spot syndrome virus | [100] |
| HS-1 | Hypsiboas semilineatus | Dengue virus | [101] |
| Myticin C | Mussel | Herpes viruses | [102] |
| P9 (β-defensin-4) | Mouse | Influenza A virus H1N1, H3N2, H5N1, H7N7, H7N9, SARS-CoV and MERS-CoV. | [103] |
| AMPs | Source | Significance | References |
|---|---|---|---|
| Poca A, Poca B and CyO4 | Pombalia calceolaria | Reduced the breast cancer cell up to 80% | [108] |
| Aurein 1.2 | Frog Litoria aurea | Among 54 cancer cells, 52 are inhibited in NCI testing method | [109] |
| Bmattacin2 | Bombyx mori | Disrupted A375 and HCT116 cancer cells | [110] |
| Laterosporulin10 | Brevibacillus sp. | MCF-7, H1299, HEK293T, HT1080, and HeLa cancer cells were disrupted | [111] |
| Dermaseptin-PD-1 and dermaseptin-PD-2 | Phyllomedusine leaf frogs | Growth of H157, PC-3, and U251 MG cancer cell was inhibited | [112] |
| Scolopendrasin VII | Centipede | Reduction in viability of leukemia cells | [113] |
| Myristoyl-CM4 | Synthetic | Activates caspase 9, caspase 3, and cleavage of PARP in breast cancer cells | [114] |
| K4R2-Nal2-S1 | Binds with lung cancer cells and results in apoptosis | [115] | |
| VLL-28 | Sulfolobus islandicus | Inhibits murine and human tumor cells | [116] |
| CopA3 | Copris tripartitus | Reduction in cell viability of gastric cancer cells | [117] |
| Pardaxin | Pardachirus marmoratus | Improved the activation of caspase-3 | [118] |
| AMPs | Source | Mechanism | References |
|---|---|---|---|
| Nisin Z | Gram positive bacteria | suppress LPS-induced pro-inflammatory cytokines | [125] |
| LL-37 | Human | Reduces pro-inflamatory mediators | [126] |
| PMAP-23 | Porcine | Induces production of IL-8 in porcine epithelial cells | [127] |
| Defensins-DLP2 and DLP4 | Hermetia illucens | Decreases the pro inflammatory cytokines production | [128] |
| Epinecidin-1 | Epinephelus coioides | Increased the expression of TNF-1 | [129] |
| cNK-2 | Chicken | Induces the expression of CCL4, CCL5 and interleukin(IL)-1β | [130] |
| Tilapia hepcidin (TH)2-3 | Pichia pastoris | Produces certain short-chain fatty acids to improve immunity | [131] |
| CRAMP | Human | Increases TLR9 expression, which suppresses cardiac hypertrophy | [132] |
| cLF36 | Camel lactoferrin | Reduces IL-2 and MUC2 expression | [133] |
| AMPs | Source | Mechanism | References |
|---|---|---|---|
| Defensins-DLP2 and DLP4 | Hermetia illucens | Induces the production of anti- inflammatory cytokines IL-10 and GM-CSF | [128] |
| cecropin-TY1 | Tabanus yao | Inhibits the production of pro-inflammatory cytokines | [135] |
| SET-M33D | Synthetic | Reduces the production of TNF-α, COX-2 IL6, KC, IP10, MIP-1, iNOS, NF-κB | [136] |
| Papiliocin (Pap12-6) | Swallowtail butterfly | Decrease in secretion of NO, TNF-α, and IL-6 | [137] |
| L-37 | Humans | Reduces intestinal inflammation | [138] |
| Lipocalin 2 (Lcn2) | Epithelial and myeloid cells | Increases cytokine expression and NFκB activation | [139] |
| Hc-cath | Hydrophis cyanocyntus | Decrease the release of pro-inflammatory cytokine and neutrophil chemoattractant | [140] |
| AMPs | Source | Mechanism | References |
|---|---|---|---|
| Cys-KR12 | Human | Suppression the LPS-induced TNF-α | [143] |
| LLKKK18 (Analog of LL37) | Humans | Rapid wound closure and reduction in oxidative stress | [141] |
| Os | Synthetic | AMP encourages collagen deposition | [144] |
| 1a(1-21)NH2 | Frog skin | Activates epidermal growth factor receptor and STAT3 protein. Promotes migration of keratinocytes (HaCaT cells) | [145] |
| Defensin-1 | Royal jelly | Promotes matrix metalloproteinase-9 secretion and increases migration of keratinocyte | [146] |
| DRGN-1 | Komodo dragon | Stimulates the migration of HEKa keratinocyte cells and activates EGFR-STAT1/3 pathway | [147] |
| Tiger-17 | Designed antimicrobial peptides | Promotes the release of TGF-β1 and IL-6 which aids in formation of tissue | [148] |
| Brevinin-2Ta | Pelophylax kl. esculentus | Angiogenesis process was observed | [149] |
| AMPs | Therapeutic Applications | Mechanism | References |
|---|---|---|---|
| Nisin A | Head and neck squamous cell carcinoma | Induces apoptosis which abbreviates tumor formation | [207] |
| Nisin A | Prevents skin cancer | Retards the DMBA induced skin carcinogenesis | [204] |
| Nisin A S29A, S29D and S29E | Prevents food borne infections | Inhibits the growth of E. coli, Cronobacter sakazakii, Salmonella entericaandTyphimurium | [208] |
| Nisin Z | Treatment of oral issues | Inhibits the growth of oral pathogenic bacteria | [209] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, A.; Luo, H.; Ramakrishna, S. Antimicrobial Peptides and Their Applications in Biomedical Sector. Antibiotics 2021, 10, 1094. https://doi.org/10.3390/antibiotics10091094
Sultana A, Luo H, Ramakrishna S. Antimicrobial Peptides and Their Applications in Biomedical Sector. Antibiotics. 2021; 10(9):1094. https://doi.org/10.3390/antibiotics10091094
Chicago/Turabian StyleSultana, Afreen, Hongrong Luo, and Seeram Ramakrishna. 2021. "Antimicrobial Peptides and Their Applications in Biomedical Sector" Antibiotics 10, no. 9: 1094. https://doi.org/10.3390/antibiotics10091094
APA StyleSultana, A., Luo, H., & Ramakrishna, S. (2021). Antimicrobial Peptides and Their Applications in Biomedical Sector. Antibiotics, 10(9), 1094. https://doi.org/10.3390/antibiotics10091094