
Compound Prioritization through Meta-Analysis Enhances the Discovery of Antimicrobial Hits against Bacterial Pathogens

Abstract
1. Introduction
2. Results
2.1. Compounds’ Prioritization Increases the Identification of Antimicrobials Effective against Bacterial Pathogens with Diverse Taxonomic and Host Range Profile
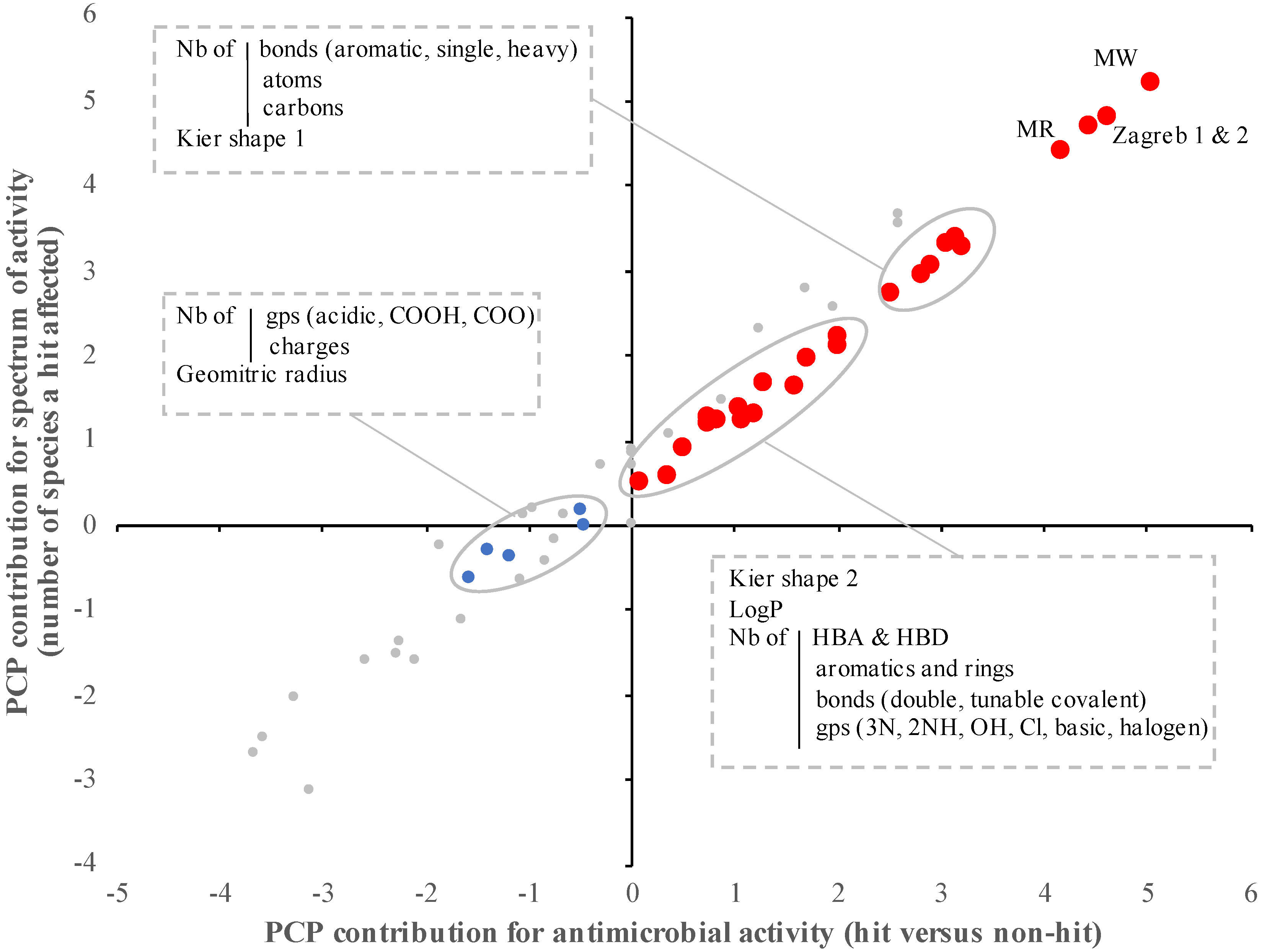
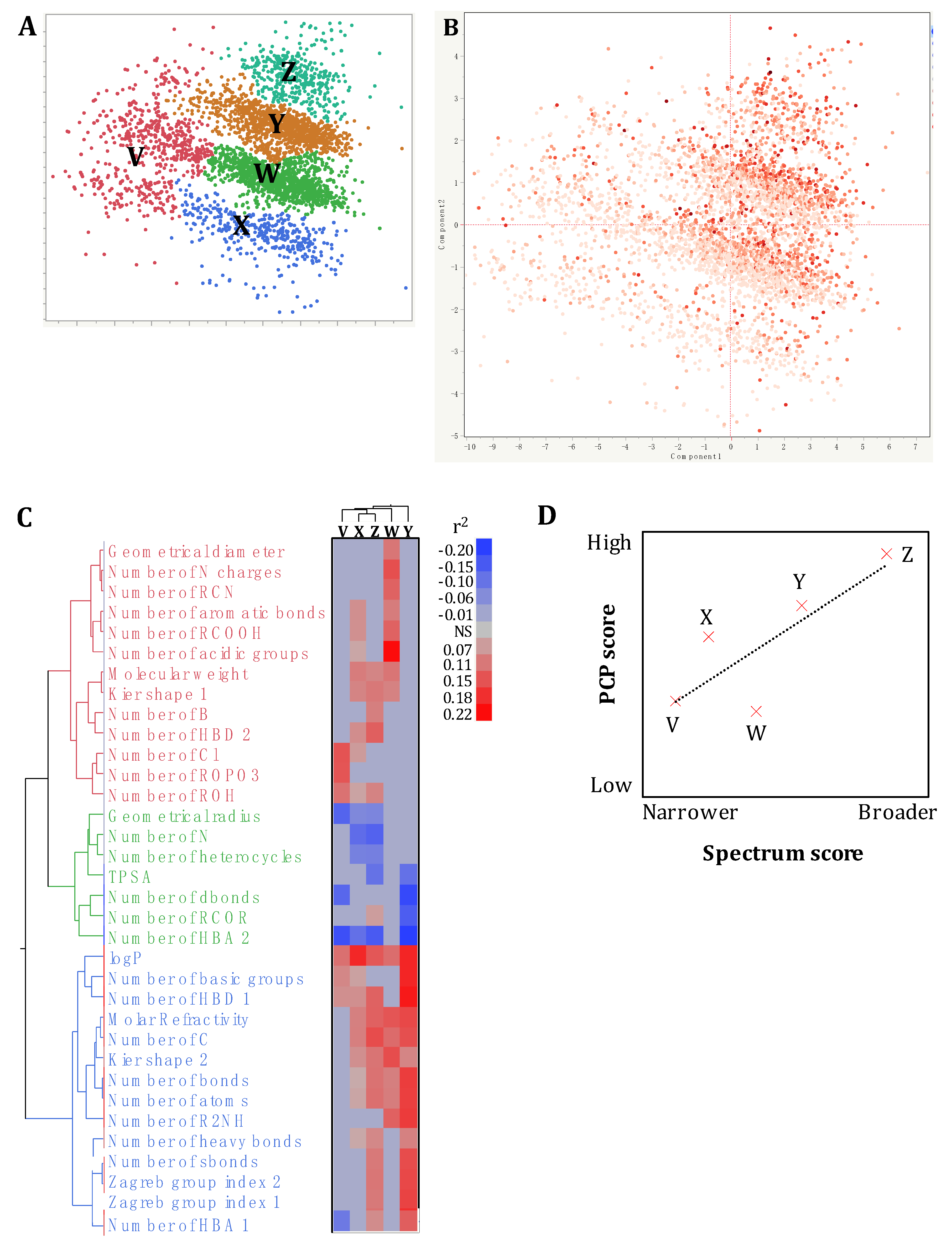
2.2. The Antimicrobial Activity and Spectrum of Activity of the SM Correlated with Specific Physico-Chemical Properties
2.3. Using Virtual Screening Tools to Prioritize the Selection of SM with Potential Antimicrobial Activity
3. Discussion
4. Materials and Methods
4.1. Physico-Chemical Properties of the Pre-Selected SM Library and High Throughput Screening Data Associated with the Pre-Selected SM Library
4.2. Statistical Analyses
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Reymond, J.-L.; Ruddigkeit, L.; Blum, L.; van Deursen, R. The Enumeration of Chemical Space. WIREs Comput. Mol. Sci. 2012, 2, 717–733. [Google Scholar] [CrossRef]
- Hughes, J.; Rees, S.; Kalindjian, S.; Philpott, K. Principles of Early Drug Discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Simpkin, V.L.; Renwick, M.J.; Kelly, R.; Mossialos, E. Incentivising Innovation in Antibiotic Drug Discovery and Development: Progress, Challenges and next Steps. J. Antibiot. 2017, 70, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Naylor, N.R.; Atun, R.; Zhu, N.; Kulasabanathan, K.; Silva, S.; Chatterjee, A.; Knight, G.M.; Robotham, J.V. Estimating the Burden of Antimicrobial Resistance: A Systematic Literature Review. Antimicrob. Resist. Infect. Control 2018, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, P.; Cooper, B.S.; Coast, J.; Oppong, R.; Do Thi Thuy, N.; Phodha, T.; Celhay, O.; Guerin, P.J.; Wertheim, H.; Lubell, Y. Enumerating the Economic Cost of Antimicrobial Resistance per Antibiotic Consumed to Inform the Evaluation of Interventions Affecting Their Use. Antimicrob. Resist. Infect. Control. 2018, 7, 98. [Google Scholar] [CrossRef]
- Segler, M.H.S.; Kogej, T.; Tyrchan, C.; Waller, M.P. Generating Focused Molecule Libraries for Drug Discovery with Recurrent Neural Networks. ACS Cent. Sci. 2018, 4, 120–131. [Google Scholar] [CrossRef]
- Rahman, A.; Choudhary, M.I. Frontiers in Anti-Infective Drug Discovery; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017; ISBN 978-1-68108-291-2. [Google Scholar]
- Ullah, H.; Ali, S. Classification of Anti-Bacterial Agents and Their Functions. Antibact. Agents 2017, 10. [Google Scholar] [CrossRef]
- Schneider, G. Prediction of Drug-Like Properties; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the Chemical Beauty of Drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Vallianatou, T.; Giaginis, C.; Tsantili-Kakoulidou, A. The Impact of Physicochemical and Molecular Properties in Drug Design: Navigation in the “Drug-like” Chemical Space. Adv. Exp. Med. Biol. 2015, 822, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Mullard, A. Re-Assessing the Rule of 5, Two Decades On. Nat. Rev. Drug Discov. 2018, 17, 777. [Google Scholar] [CrossRef]
- Wallace, I.M.; Urbanus, M.L.; Luciani, G.M.; Burns, A.R.; Han, M.K.L.; Wang, H.; Arora, K.; Heisler, L.E.; Proctor, M.; Onge, R.P.S.; et al. Compound Prioritization Methods Increase Rates of Chemical Probe Discovery in Model Organisms. Chem. Biol. 2011, 18, 1273–1283. [Google Scholar] [CrossRef]
- Xu, X.; Kumar, A.; Deblais, L.; Pina-Mimbela, R.; Nislow, C.; Fuchs, J.R.; Miller, S.A.; Rajashekara, G. Discovery of Novel Small Molecule Modulators of Clavibacter Michiganensis Subsp. Michiganensis. Front. Microbiol. 2015, 6, 1127. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Deblais, L.; Helmy, Y.A.; Kathayat, D.; Huang, H.; Miller, S.A.; Rajashekara, G. Novel Imidazole and Methoxybenzylamine Growth Inhibitors Affecting Salmonella Cell Envelope Integrity and Its Persistence in Chickens. Sci. Rep. 2018, 8, 13381. [Google Scholar] [CrossRef] [PubMed]
- Deblais, L.; Helmy, Y.A.; Kumar, A.; Antwi, J.; Kathayat, D.; Acuna, U.M.; Huang, H.; de Blanco, E.C.; Fuchs, J.R.; Rajashekara, G. Novel Narrow Spectrum Benzyl Thiophene Sulfonamide Derivatives to Control Campylobacter. J. Antibiot. 2019, 72, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Deblais, L.; Vrisman, C.; Kathayat, D.; Helmy, Y.A.; Miller, S.A.; Rajashekara, G. Imidazole and Methoxybenzylamine Growth Inhibitors Reduce Salmonella Persistence in Tomato Plant Tissues. J. Food Prot. 2019, 82, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Deblais, L.; Kassem, I.I.; Kathayat, D.; Rajashekara, G. Novel Small Molecule Modulators of Quorum Sensing in Avian Pathogenic Escherichia Coli (APEC). Virulence 2018, 9, 1640–1657. [Google Scholar] [CrossRef] [PubMed]
- Kathayat, D.; Helmy, Y.A.; Deblais, L.; Rajashekara, G. Novel Small Molecules Affecting Cell Membrane as Potential Therapeutics for Avian Pathogenic Escherichia Coli. Sci. Rep. 2018, 8, 15329. [Google Scholar] [CrossRef]
- Vrisman, C.M.; Deblais, L.; Helmy, Y.; Johnson, R.; Rajashekara, G.; Miller, S.A. Discovery and Characterization of Low Molecular Weight Inhibitors of Erwinia Tracheiphila. Phytopathology 2020, 110, 989–998. [Google Scholar] [CrossRef]
- Kumar, A.; Drozd, M.; Pina-Mimbela, R.; Xu, X.; Helmy, Y.A.; Antwi, J.; Fuchs, J.R.; Nislow, C.; Templeton, J.; Blackall, P.J.; et al. Novel Anti-Campylobacter Compounds Identified Using High Throughput Screening of a Pre-Selected Enriched Small Molecules Library. Front. Microbiol. 2016, 7, 405. [Google Scholar] [CrossRef]
- Koopman, J.A.; Marshall, J.M.; Bhatiya, A.; Eguale, T.; Kwiek, J.J.; Gunn, J.S. Inhibition of Salmonella Enterica Biofilm Formation Using Small-Molecule Adenosine Mimetics. Antimicrob. Agents Chemother. 2015, 59, 76–84. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Kathayat, D.; Ghanem, M.; Jung, K.; Closs, G.; Deblais, L.; Srivastava, V.; El-Gazzar, M.; Rajashekara, G. Identification and Characterization of Novel Small Molecule Inhibitors to Control Mycoplasma Gallisepticum Infection in Chickens. Vet. Microbiol. 2020, 247, 108799. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Deblais, L.; Kathayat, D.; Rotondo, F.; Helmy, Y.A.; Miller, S.A.; Rajashekara, G. Novel Small Molecule Growth Inhibitors of Xanthomonas spp. Causing Bacterial Spot of Tomato. Phytopathology 2020. Available online: https://apsjournals.apsnet.org/doi/abs/10.1094/PHYTO-08-20-0341-R (accessed on 30 August 2021).
- Lu, Y.; Deblais, L.; Rajashekara, G.; Miller, S.A.; Helmy, Y.A.; Zhang, H.; Wu, P.; Qiu, Y.; Xu, X. High-Throughput Screening Reveals Small Molecule Modulators Inhibitory to Acidovorax Citrulli. Plant Pathol. 2020, 69, 818–826. [Google Scholar] [CrossRef]
- Iredell, J.; Brown, J.; Tagg, K. Antibiotic Resistance in Enterobacteriaceae: Mechanisms and Clinical Implications. BMJ 2016, 352, h6420. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R. Resistance Mechanisms in Enterobacteriaceae. Pathology 2015, 47, 276–284. [Google Scholar] [CrossRef]
- O’Shea, R.; Moser, H.E. Physicochemical Properties of Antibacterial Compounds: Implications for Drug Discovery. J. Med. Chem. 2008, 51, 2871–2878. [Google Scholar] [CrossRef]
- Ebejer, J.-P.; Charlton, M.H.; Finn, P.W. Are the Physicochemical Properties of Antibacterial Compounds Really Different from Other Drugs? J. Cheminformatics 2016, 8, 30. [Google Scholar] [CrossRef]
- Sahariah, P.; Cibor, D.; Zielińska, D.; Hjálmarsdóttir, M.Á.; Stawski, D.; Másson, M. The Effect of Molecular Weight on the Antibacterial Activity of N,N,N-Trimethyl Chitosan (TMC). Int. J. Mol. Sci. 2019, 20, 1743. [Google Scholar] [CrossRef]
- Cho, C.-W.; Park, J.-S.; Stolte, S.; Yun, Y.-S. Modelling for Antimicrobial Activities of Ionic Liquids towards Escherichia Coli, Staphylococcus Aureus and Candida Albicans Using Linear Free Energy Relationship Descriptors. J. Hazard. Mater. 2016, 311, 168–175. [Google Scholar] [CrossRef]
- Mugumbate, G.; Overington, J.P. The Relationship between Target-Class and the Physicochemical Properties of Antibacterial Drugs. Bioorganic Med. Chem. 2015, 23, 5218–5224. [Google Scholar] [CrossRef]
- Bajaj, S.; Sambi, S.S.; Madan, A.K. Prediction of Anti-Inflammatory Activity of N-Arylanthranilic Acids: Computational Approach Using Refined Zagreb Indices. Croat. Chem. Acta 2005, 78, 165–174. [Google Scholar]
- Verma, V.; Singh, K.; Kumar, D.; Narasimhan, B. QSAR Studies of Antimicrobial Activity of 1,3-Disubstituted-1H-Naphtho[1,2-e][1,3]Oxazines Using Topological Descriptors. Arab. J. Chem. 2017, 10, S747–S756. [Google Scholar] [CrossRef]
- Rezaee, S.; Khalaj, A.; Adibpour, N.; Saffary, M. Correlation between Lipophilicity and Antimicrobial Activity of Some 2-(4-Substituted Phenyl)-3(2H)-Isothiazolones. Daru-J. Fac. Pharm. 2009, 17, 256–263. [Google Scholar]
- Echeverría, J.; Urzúa, A.; Sanhueza, L.; Wilkens, M. Enhanced Antibacterial Activity of Ent-Labdane Derivatives of Salvic Acid (7α-Hydroxy-8(17)-Ent-Labden-15-Oic Acid): Effect of Lipophilicity and the Hydrogen Bonding Role in Bacterial Membrane Interaction. Molecules 2017, 22, 1039. [Google Scholar] [CrossRef]
- Chibane, L.B.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant Antimicrobial Polyphenols as Potential Natural Food Preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Arfa, A.B.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier, P. Antimicrobial Activity of Carvacrol Related to Its Chemical Structure. Lett. Appl. Microbiol. 2006, 43, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, J.; Opazo, J.; Mendoza, L.; Urzúa, A.; Wilkens, M. Structure-Activity and Lipophilicity Relationships of Selected Antibacterial Natural Flavones and Flavanones of Chilean Flora. Molecules 2017, 22, 608. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial Properties of Plant Essential Oils against Human Pathogens and Their Mode of Action: An Updated Review. Available online: https://www.hindawi.com/journals/ecam/2016/3012462/ (accessed on 16 March 2020).
- Almeida, R.P.P.; Killiny, N.; Newman, K.L.; Chatterjee, S.; Ionescu, M.; Lindow, S.E. Contribution of RpfB to Cell-to-Cell Signal Synthesis, Virulence, and Vector Transmission of Xylella Fastidiosa. Mol. Plant Microbe Interact. 2012, 25, 453–462. [Google Scholar] [CrossRef]
- ’t Hart, P.; Wood, T.M.; Tehrani, K.H.M.E.; van Harten, R.M.; Śleszyńska, M.; Rentero Rebollo, I.; Hendrickx, A.P.A.; Willems, R.J.L.; Breukink, E.; Martin, N.I. De Novo Identification of Lipid II Binding Lipopeptides with Antibacterial Activity against Vancomycin-Resistant Bacteria. Chem. Sci. 2017, 8, 7991–7997. [Google Scholar] [CrossRef]
- Hsiao, C.P.; Siebert, K.J. Modeling the Inhibitory Effects of Organic Acids on Bacteria. Int. J. Food Microbiol. 1999, 47, 189–201. [Google Scholar] [CrossRef]
- Totaro, G.; Cruciani, L.; Vannini, M.; Mazzola, G.; Di Gioia, D.; Celli, A.; Sisti, L. Synthesis of Castor Oil-Derived Polyesters with Antimicrobial Activity. Eur. Polym. J. 2014, 56, 174–184. [Google Scholar] [CrossRef]
- Sayin, Z.; Ucan, U.S.; Sakmanoglu, A. Antibacterial and Antibiofilm Effects of Boron on Different Bacteria. Biol. Trace Elem. Res. 2016, 173, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Urzúa, A.; Rezende, M.C.; Mascayano, C.; Vásquez, L. A Structure-Activity Study of Antibacterial Diterpenoids. Molecules 2008, 13, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Backman, T.W.H.; Cao, Y.; Girke, T. ChemMine Tools: An Online Service for Analyzing and Clustering Small Molecules. Nucleic Acids Res. 2011, 39, W486–W491. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 Update: Improved Access to Chemical Data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminformatics 2011, 3, 33. [Google Scholar] [CrossRef]
- Guha, R.; Howard, M.T.; Hutchison, G.R.; Murray-Rust, P.; Rzepa, H.; Steinbeck, C.; Wegner, J.; Willighagen, E.L. The Blue Obelisk—Interoperability in Chemical Informatics. J. Chem. Inf. Model. 2006, 46, 991–998. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Pathogens Screened | [SM] (μM) | Growing Conditions | Hit Rate (%) | References |
|---|---|---|---|---|
| Growth Inhibitor Screenings | ||||
| Acidovorax citrulli Xu9-15 | 100 | 50% NBY A | 1.4 | Lu et al., 2020 |
| Clavibacter michiganensis subsp michiganensis C280 | 100 | NBY A | 11.2 | Xu et al., 2016 |
| Erwinia tracheiphila TedCu10 | 100 | 50% NBY A | 11.1 | Vrisman et al., 2020 |
| Xanthomonas gardneri SM761 | 100 | MMX B | 29.3 | Srivastava et al., 2020 |
| Xanthomonas perforans SM755-12 | 100 | MMX B | 17.9 | Srivastava et al., 2020 |
| Avian pathogenic Escherichia coli O78 | 100 | M63 C | 1 | Kathayat et al., 2019 |
| Mycoplasma gallisepticum MG37 | 100 | FREY C | 14.1 | Helmy et al., 2020 |
| Campylobacter jejuni 81–176 | 100 | MH D | 18.7 | Kumar et al., 2017 |
| Salmonella enterica subsp. enterica serotype Typhimurium LT2 | 200 | M9 C | 0.5 | Deblais et al., 2018 |
| Virulence Inhibitor Screenings | ||||
| Avian pathogenic Escherichia coli O78 | 100 A | M63 C | 2.4 F | Helmy et al., 2020 |
| Salmonella enterica subsp. enterica serotype Typhimurium LT2 | 10 B | TSB E | 5.2 G | Koopman et al., 2015 |
| Antimicrobial Spectrum | Number of Hits per Pathogen Category | Number of Hits Across All the Pathogen Tested (n = 9) | ||
|---|---|---|---|---|
| Plant (n = 5) | Animal (n = 2) | Foodborne (n = 2) | ||
| Nonhit | 2601 | 3491 | 3231 | 2109 |
| 1 species | 752 | 651 | 885 | 926 |
| 2 species | 438 | 40 | 66 | 442 |
| 3 species | 243 | 277 | ||
| 4 species | 135 | 202 | ||
| 5 species | 13 | 135 | ||
| 6 species | 59 | |||
| 7 species | 23 | |||
| 8 species | 8 | |||
| 9 species | 1 | |||
| Total number of hits across the whole pre-selected library (n = 4182) | 1581 | 691 | 951 | 2073 |
| Screening Type | Bacterial Pathogens | Lead Compounds (Pubchem ID) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1529361 | 2827372 | 1380897 | 2847561 | 2848076 | 5731123 | 16876368 | 42115615 | 42115777 | 45191821 | 45192477 | 25365835 | 45195011 | 42525758 | 25304876 | 25313118 | 42520454 | 45238750 | ||
| GI | Et | X | X | X | X | X | |||||||||||||
| Cmm | X | X | X | ||||||||||||||||
| Xp | X | X | X | X | |||||||||||||||
| Xg | X | X | X | X | |||||||||||||||
| APEC | X | ||||||||||||||||||
| Mg | X | X | |||||||||||||||||
| Cj | X | ||||||||||||||||||
| ST | X | X | |||||||||||||||||
| VI | APEC | X | |||||||||||||||||
| Physico-Chemical Properties of the SM Used for This Study (n = 60) | Antimicrobial Activity (Hit versus Nonactive SM) | Spectrum of Activity (Nb of Species Affected by SM) | Growth Inhibitors versus Virulence Inhibitors | Lead Compounds versus Other Hits | ||||
|---|---|---|---|---|---|---|---|---|
| Contribution Score | p-Value | Contribution Score | p-Value | Contribution Score | p-Value | Contribution Score | p-Value | |
| Geometrical diameter | 7.61 | 0.203 | 29.63 | 0.305 | 169.84 | <0.001 | 6.25 | 0.253 |
| Geometrical radius | 0.07 | 0.021 | 0.46 | <0.001 | 0.02 | 0.254 | 0.02 | 0.197 |
| Kier shape 1 | 326.23 | <0.001 | 568.69 | <0.001 | 574.78 | <0.001 | 6.76 | 0.739 |
| Kier shape 2 | 99.72 | <0.001 | 176.7 | <0.001 | 138.93 | <0.001 | 6.83 | 0.128 |
| Lipophilicity (logP) | 101.87 | <0.001 | 134.83 | <0.001 | 17.47 | <0.001 | 12.21 | 0.071 |
| Molar Refractivity | 14,978.5 | <0.001 | 26,375.9 | <0.001 | 26,292.7 | <0.001 | 572.9 | 0.426 |
| Molecular weight | 109,599 | <0.001 | 166,671 | <0.001 | 163,353 | <0.001 | 1249.76 | 0.878 |
| Number of acidic groups | 0.04 | 0.423 | 0.53 | 0.001 | 0.02 | 0.757 | 0.18 | 0.009 |
| Number of aromatic bonds | 1617.33 | <0.001 | 1952.13 | <0.001 | 559.94 | <0.001 | 13.63 | 0.632 |
| Number of aromatic groups | 37.09 | <0.001 | 44.38 | <0.001 | 22.15 | <0.001 | 0.68 | 0.328 |
| Number of atoms | 1079.04 | <0.001 | 2205.34 | <0.001 | 4288.17 | <0.001 | 187.23 | 0.177 |
| Number of basic groups | 3.08 | 0.004 | 8.42 | 0.001 | 5.5 | <0.001 | 0.42 | 0.334 |
| Number of bonds | 1339.92 | <0.001 | 2580.42 | <0.001 | 4965.25 | <0.001 | 252.18 | 0.101 |
| Number of carbon | 645.53 | <0.001 | 959.85 | <0.001 | 764.19 | <0.001 | 29.41 | 0.120 |
| Number of charges | 0.31 | 0.031 | 1.57 | 0.002 | 0.27 | 0.066 | 2.31 | <0.001 |
| Number of chlorine | 2.21 | <0.001 | 3.85 | <0.001 | 0.17 | 0.637 | 0.94 | 0.033 |
| Number of double bonds | 11.3 | <0.001 | 18.39 | <0.001 | 6.75 | <0.001 | 4.49 | 0.010 |
| Number of hydrogen | 89.86 | 0.164 | 374.23 | 0.019 | 1185.4 | <0.001 | 75.43 | 0.187 |
| Number of halogens | 5.22 | 0.004 | 16.37 | <0.001 | 0.39 | 0.914 | 1.67 | 0.149 |
| Number of HBA1 | 17.46 | 0.897 | 208.74 | 0.436 | 1140.71 | <0.001 | 71.69 | 0.176 |
| Number of HBA2 | 48.85 | <0.001 | 93.98 | <0.001 | 18.06 | <0.001 | 3.71 | 0.123 |
| Number of HBD 1 | 19.11 | <0.001 | 47.64 | <0.001 | 3.83 | 0.006 | 2 | 0.063 |
| Number of HBD 2 | 10.67 | <0.001 | 25.51 | <0.001 | 2.05 | 0.051 | 0.58 | 0.550 |
| Number of heavy bonds | 800.79 | <0.001 | 1195.37 | <0.001 | 1320.5 | <0.001 | 48.79 | 0.079 |
| Number of heterocycles | 0.51 | 0.613 | 5.26 | 0.048 | 19.56 | <0.001 | 11.95 | <0.001 |
| Number of nitrogen | 2.8 | 0.106 | 8.25 | 0.083 | 21.19 | <0.001 | 0.47 | 0.697 |
| Number of NO2 | 0.18 | 0.081 | 0.68 | 0.393 | 0.42 | 0.009 | 0.89 | <0.001 |
| Number of R-2NH | 5.28 | <0.001 | 19.71 | <0.001 | 0.68 | 0.060 | 2.44 | <0.001 |
| Number of R-3N | 6.55 | <0.001 | 18.59 | <0.001 | 1.25 | 0.117 | 3.3 | 0.006 |
| Number of R-CN | 0.01 | 0.349 | 0.03 | 0.194 | 0 | 0.839 | 0.12 | <0.001 |
| Number of R-COO-R | 0.35 | 0.029 | 1.04 | 0.002 | 0.04 | 0.750 | 0.03 | 0.764 |
| Number of R-COOH | 0.03 | 0.072 | 0.25 | <0.001 | 0.03 | 0.252 | 0.02 | 0.296 |
| Number of R-OH | 1.18 | 0.002 | 3.44 | <0.001 | 1.36 | 0.138 | 1.54 | 0.087 |
| Number of RINGS | 14.73 | <0.001 | 21.92 | <0.001 | 22.23 | <0.001 | 7.39 | <0.001 |
| Number of single bonds | 48.57 | 0.991 | 625.48 | 0.501 | 2326.73 | <0.001 | 269.52 | 0.111 |
| Zagreb group index 1 | 27,153.5 | <0.001 | 54,499.4 | <0.001 | 133,000 | <0.001 | 10,537.2 | 0.059 |
| Zagreb group index 2 | 40,446.8 | <0.001 | 71,160 | <0.001 | 153,738 | <0.001 | 11,507.2 | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deblais, L.; Rajashekara, G. Compound Prioritization through Meta-Analysis Enhances the Discovery of Antimicrobial Hits against Bacterial Pathogens. Antibiotics 2021, 10, 1065. https://doi.org/10.3390/antibiotics10091065
Deblais L, Rajashekara G. Compound Prioritization through Meta-Analysis Enhances the Discovery of Antimicrobial Hits against Bacterial Pathogens. Antibiotics. 2021; 10(9):1065. https://doi.org/10.3390/antibiotics10091065
Chicago/Turabian StyleDeblais, Loic, and Gireesh Rajashekara. 2021. "Compound Prioritization through Meta-Analysis Enhances the Discovery of Antimicrobial Hits against Bacterial Pathogens" Antibiotics 10, no. 9: 1065. https://doi.org/10.3390/antibiotics10091065
APA StyleDeblais, L., & Rajashekara, G. (2021). Compound Prioritization through Meta-Analysis Enhances the Discovery of Antimicrobial Hits against Bacterial Pathogens. Antibiotics, 10(9), 1065. https://doi.org/10.3390/antibiotics10091065