
Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
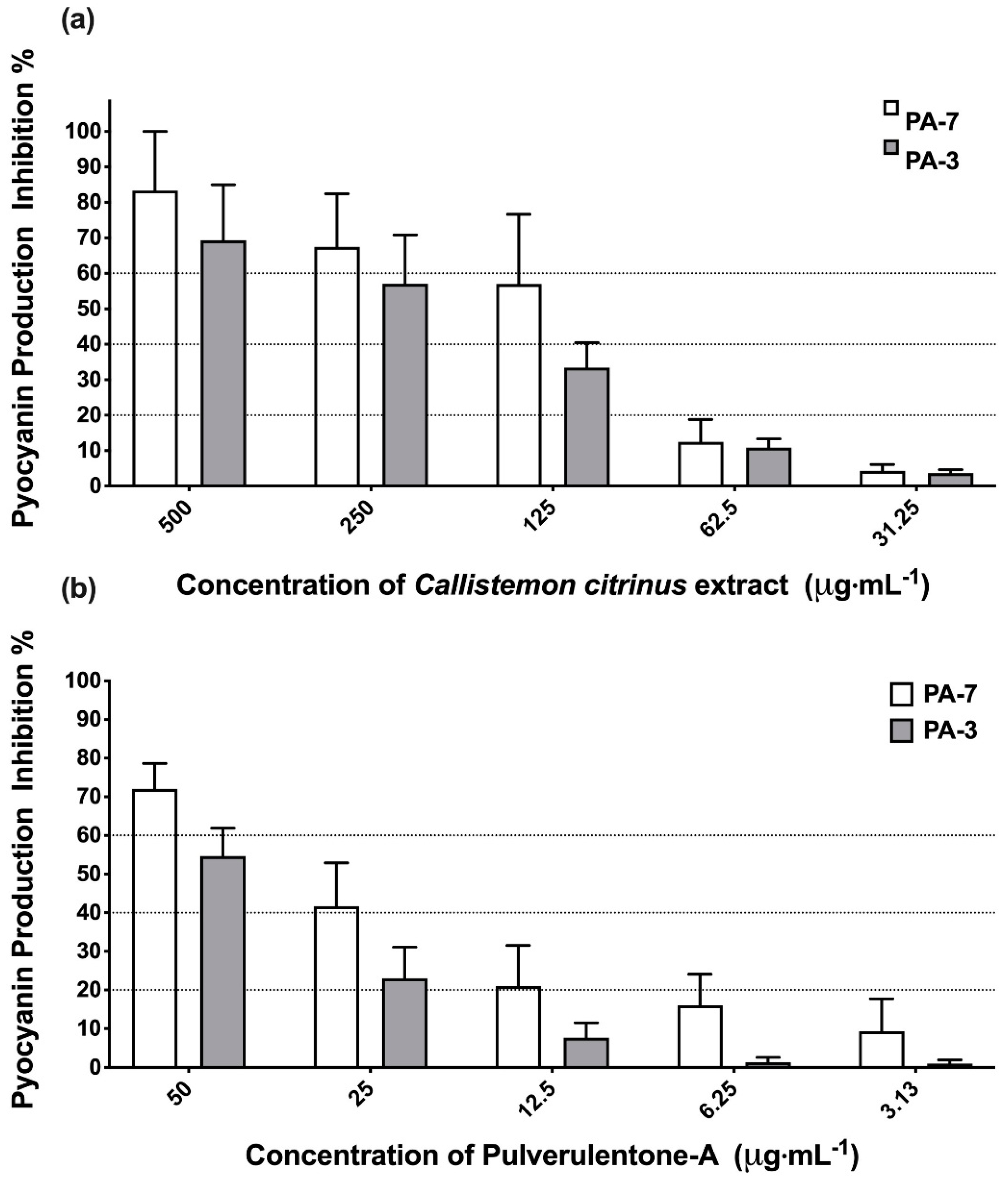
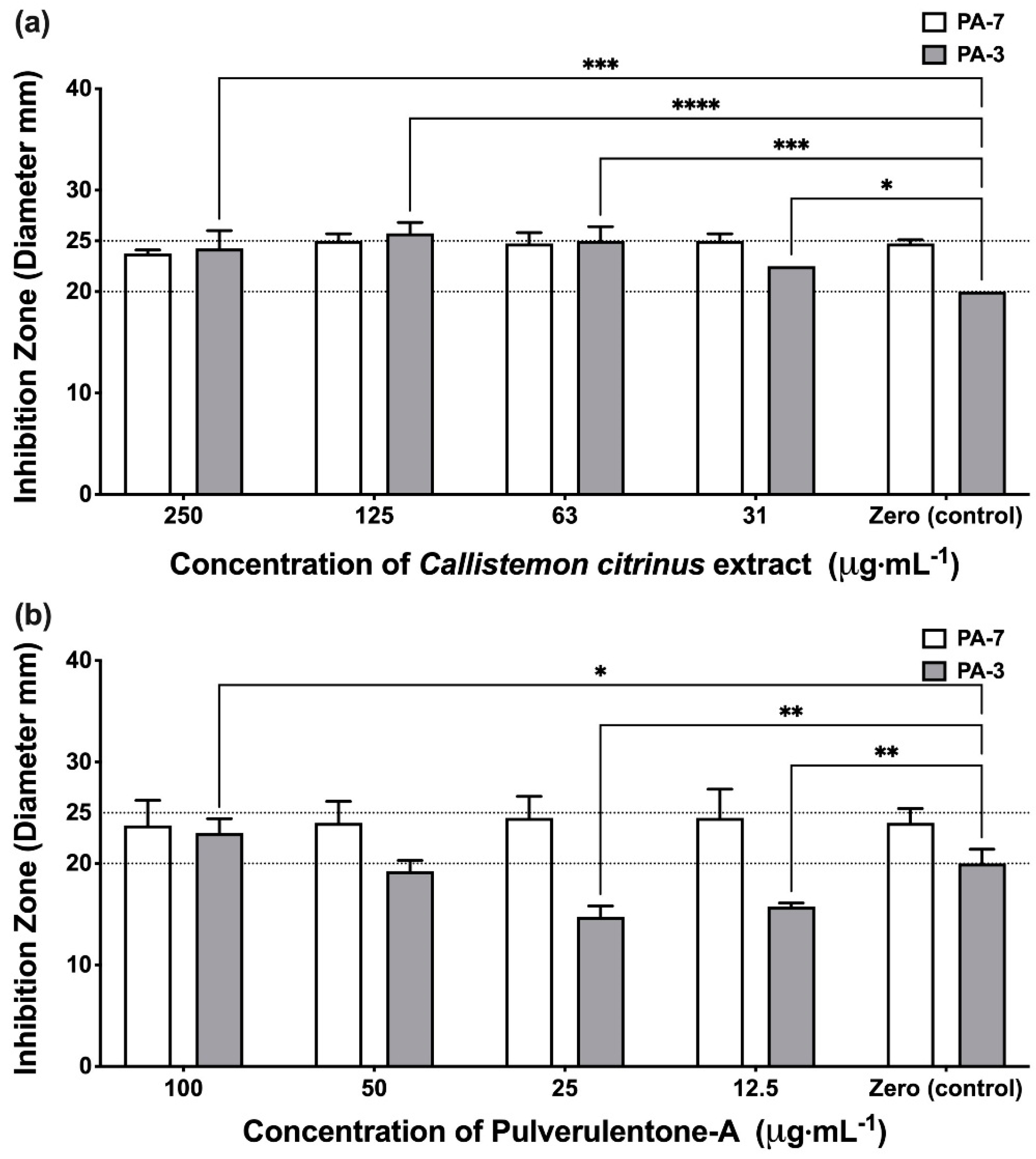
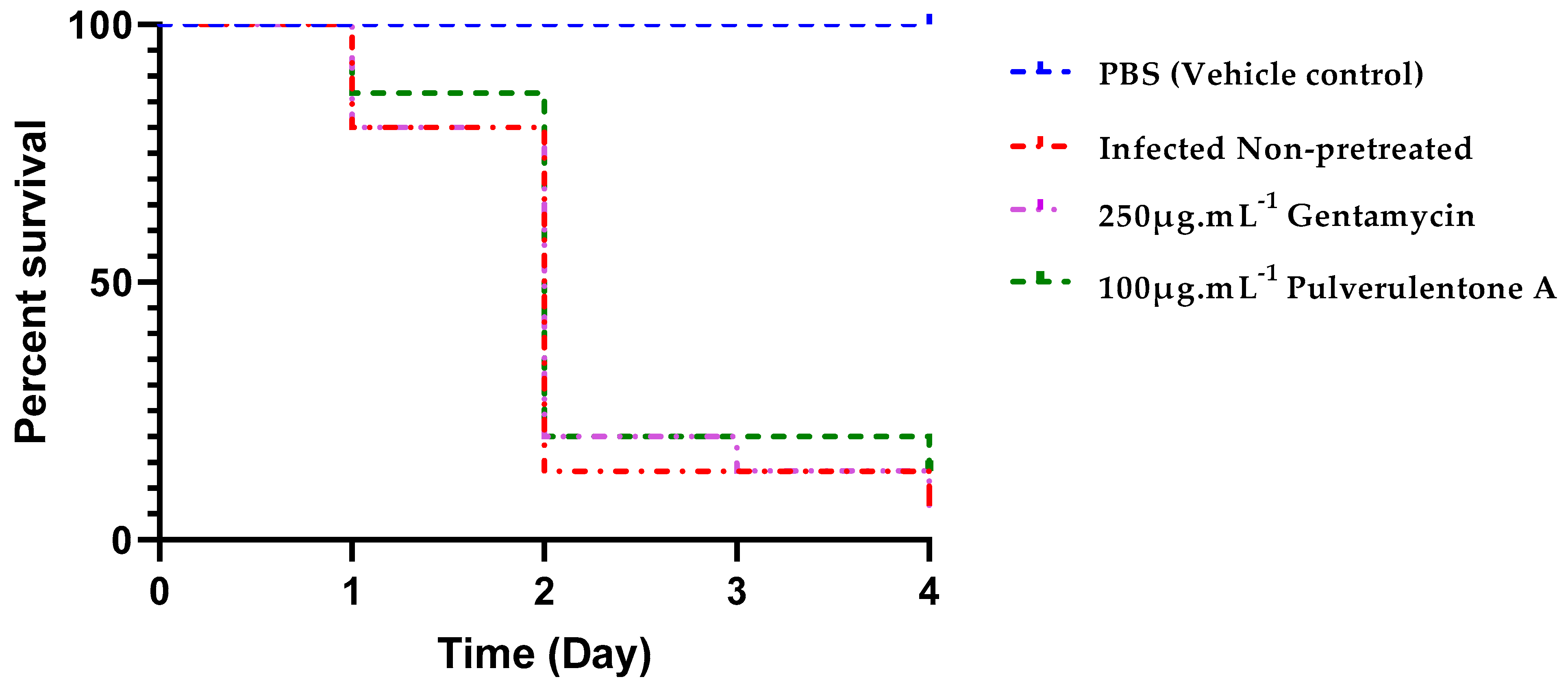
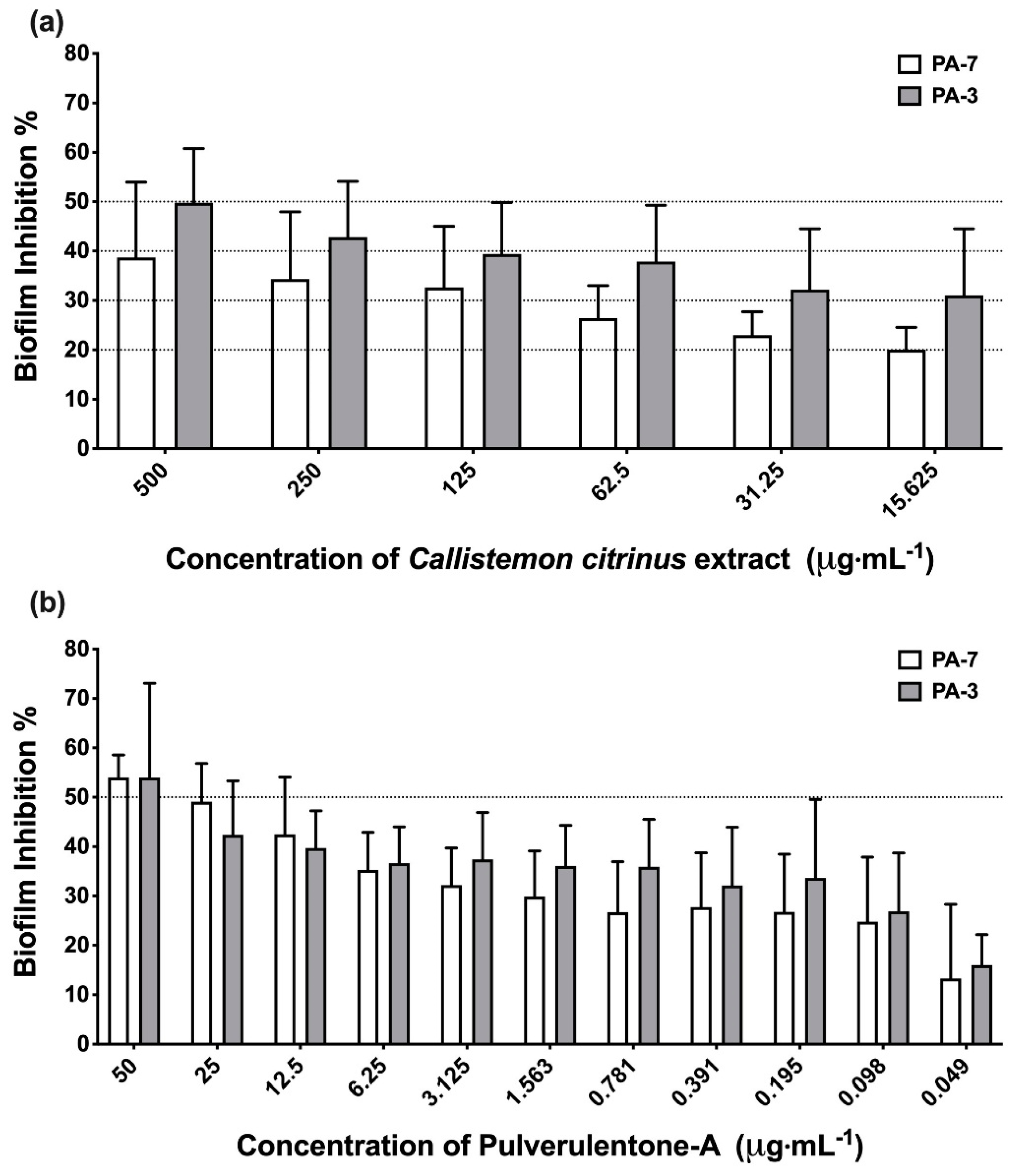
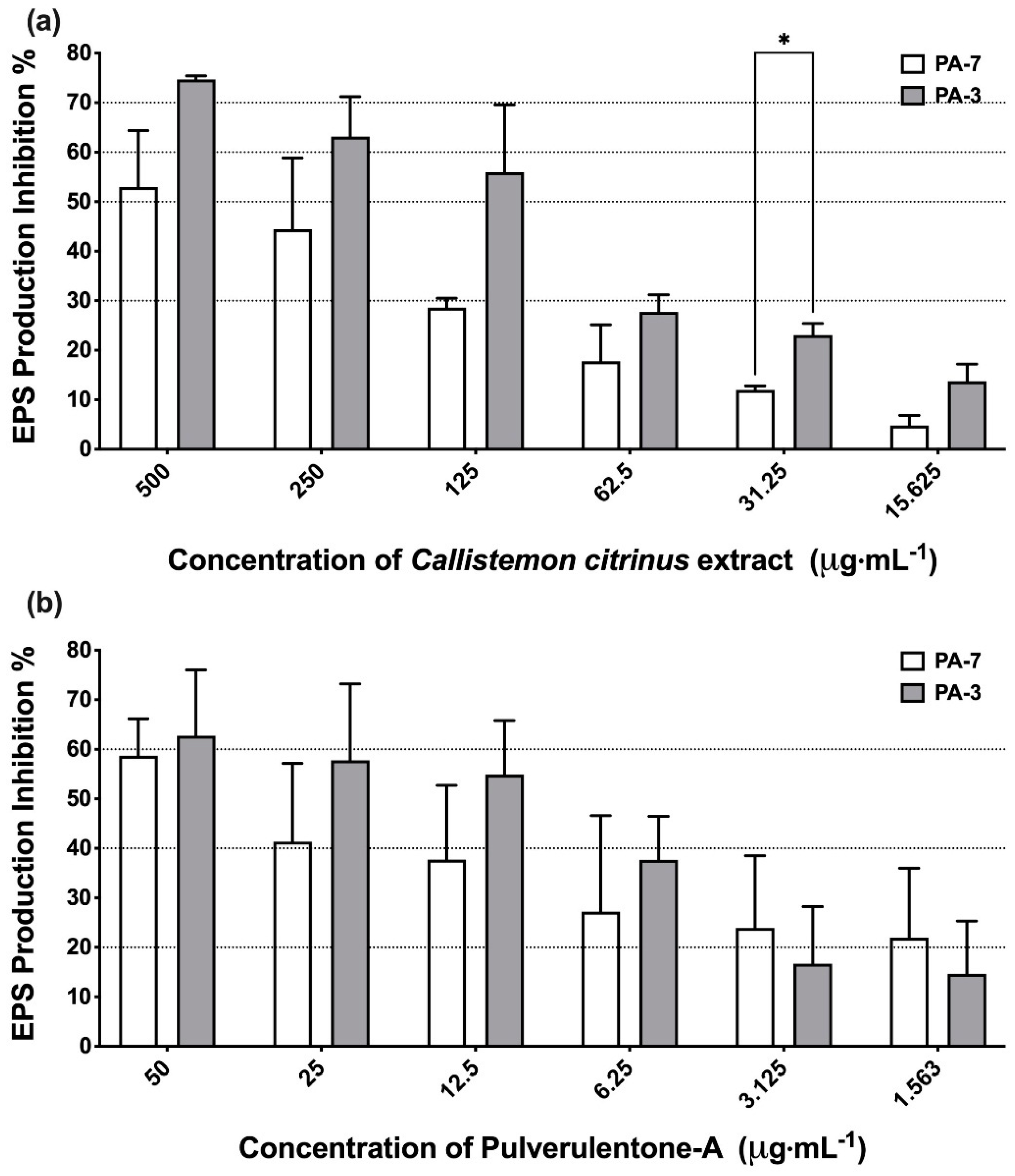
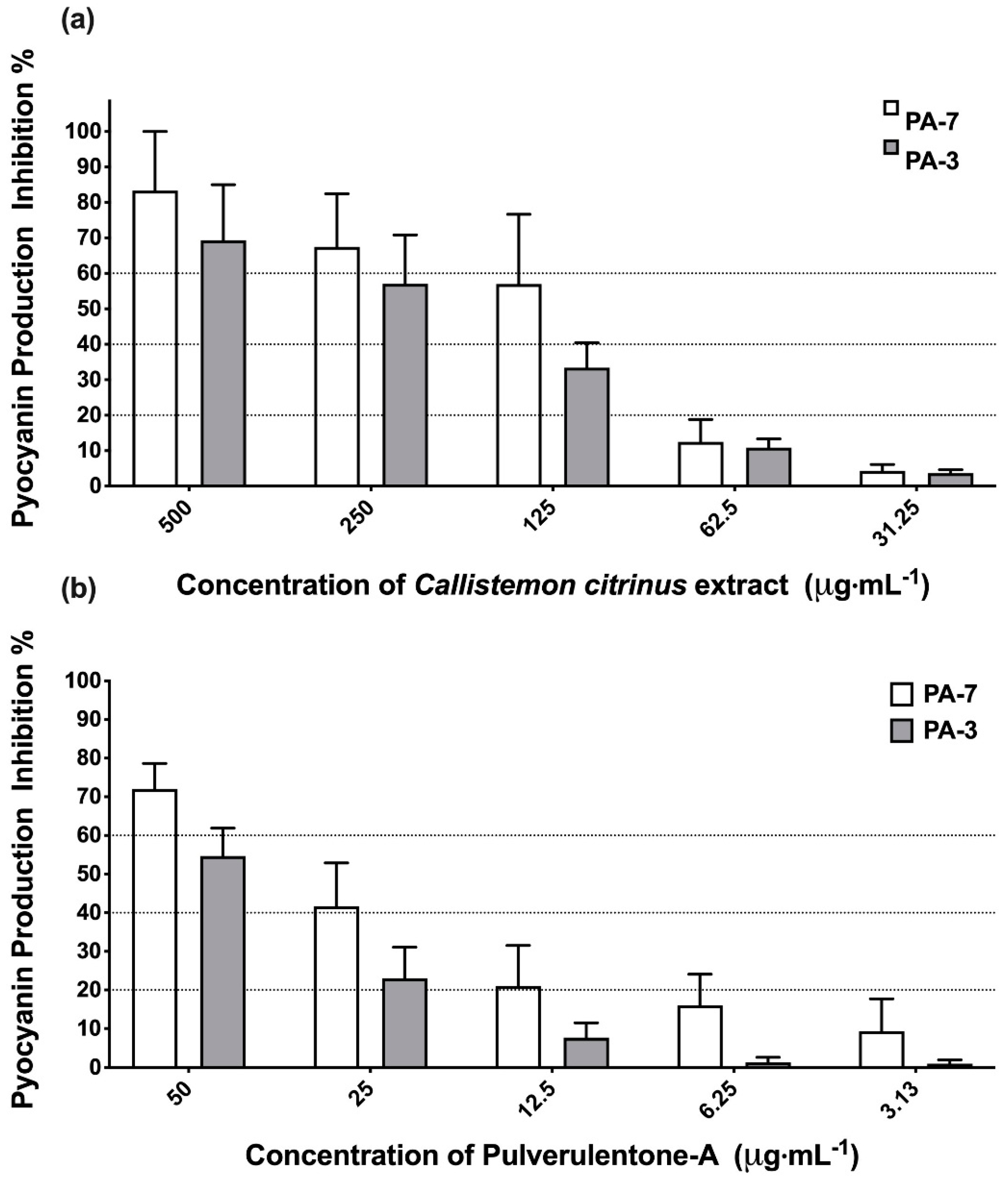
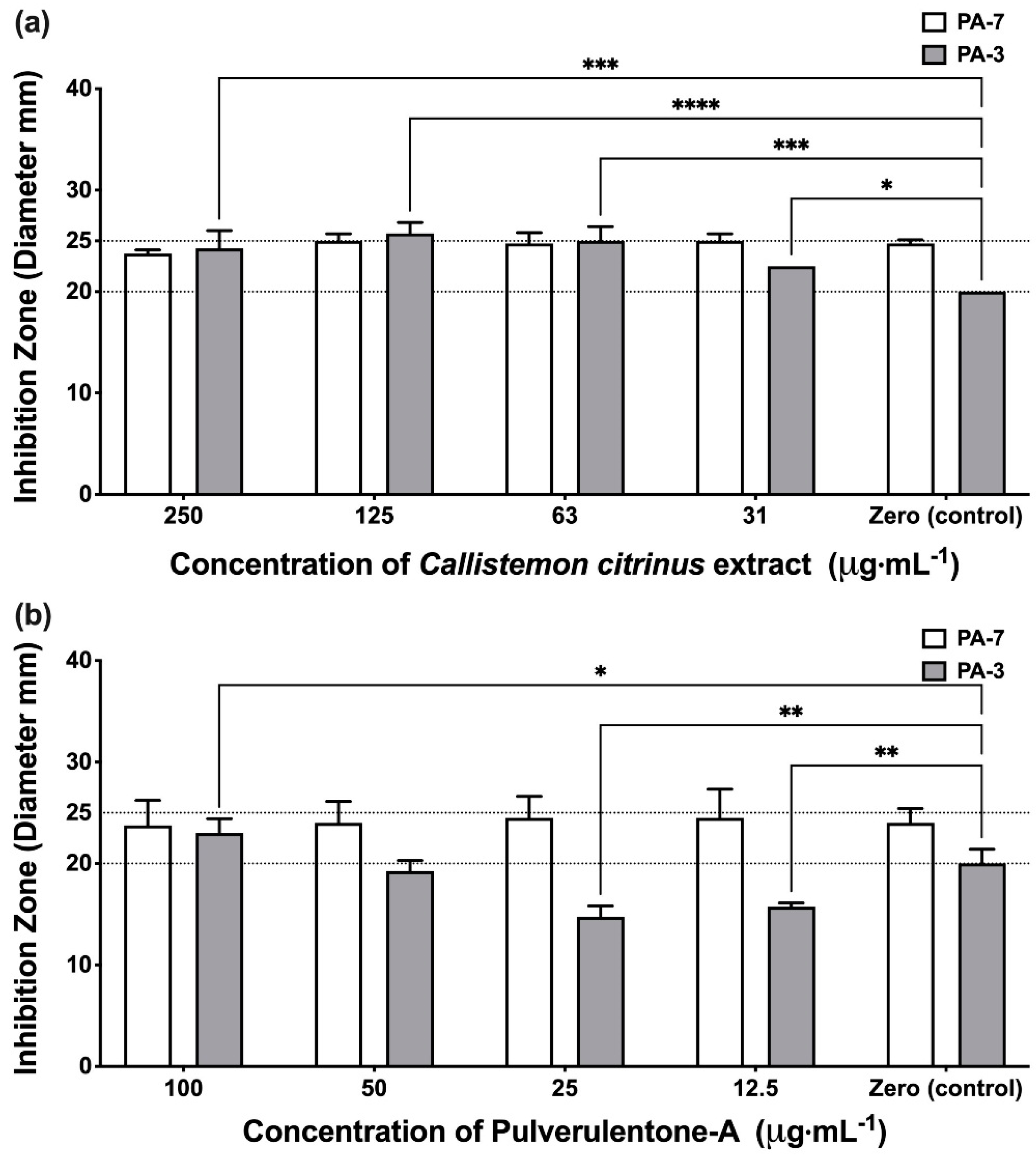
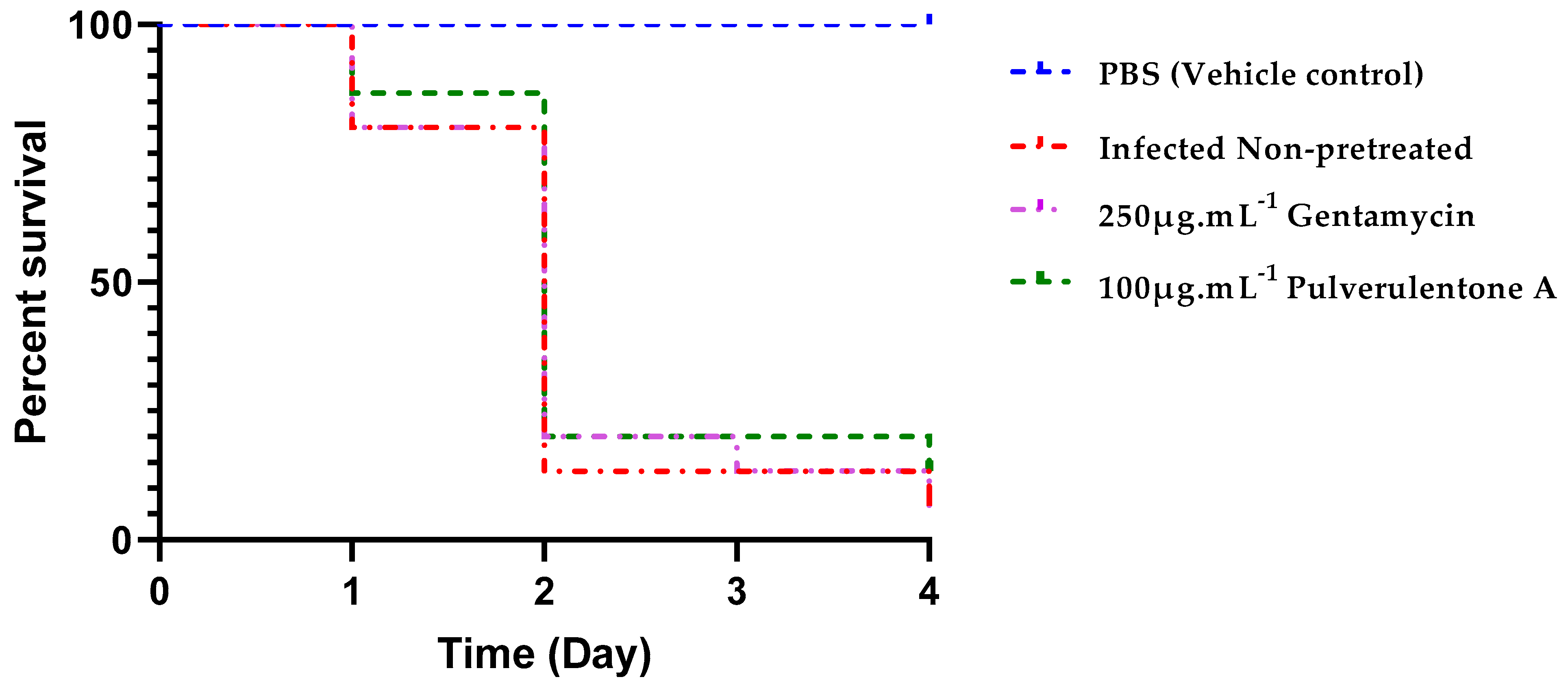
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of the Extract and Isolation of Pulverulentone A
3.3. Microorganisms and Growth Conditions
3.4. Effect of the Extract/Pulverulentone A on Biofilm Formation
3.4.1. Screening for Biofilm Formation by the Tested Isolates
3.4.2. Biofilm Inhibition Activity of the Extract/Pulverulentone A
3.5. Quantification of Extracellular Polymeric Substances Represented as Polysaccharides
3.6. Effect of the Extract/Pulverulentone A on pyocyanin Pigment Production
3.7. Effect of the Extract/Pulverulentone A on the Proteases Production
3.8. Molecular Docking Study
3.9. In Vivo Cytotoxicity of Pulverulentone A Using Galleria mellonella Larvae Model
3.10. Survival of Galleria mellonella Larvae Model Infected with Non-Pretreated or Pretreated P. aeruginosa with Pulverulentone A
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kierbel, A.; Gassama-Diagne, A.; Rocha, C.; Radoshevich, L.; Olson, J.; Mostov, K.; Engel, J. Pseudomonas aeruginosa exploits a PIP3-dependent pathway to transform apical into basolateral membrane. J. Cell Biol. 2007, 177, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Pericolini, E.; Colombari, B.; Ferretti, G.; Iseppi, R.; Ardizzoni, A.; Girardis, M.; Sala, A.; Peppoloni, S.; Blasi, E. Real-time monitoring of Pseudomonas aeruginosa biofilm formation on endotracheal tubes in vitro. BMC Microbiol. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Centers for Disease Control and Prevention: Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019.
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D. Plant Natural products targeting bacterial virulence factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.M.; Samir, R.; Saber, F.R.; Ahmed, S.R.; Farag, M.A. Pimenta Oil as A Potential Treatment for Acinetobacter Baumannii wound infection: In vitro and in vivo bioassays in relation to its chemical composition. Antibiotics 2020, 9, 679. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-W.; Muhammad, J.; Sun, B.; Yang, R.; Wadood, A.; Wang, J.-S.; Jia, A.-Q. Metabolomic analysis of quorum sensing inhibitor hordenine on Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2019, 103, 6271–6285. [Google Scholar] [CrossRef]
- Zhong, L.; Ravichandran, V.; Zhang, N.; Wang, H.; Bian, X.; Zhang, Y.; Li, A. Attenuation of Pseudomonas aeruginosa Quorum Sensing by Natural Products: Virtual Screening, Evaluation and Biomolecular Interactions. Int. J. Mol. Sci. 2020, 21, 2190. [Google Scholar] [CrossRef] [Green Version]
- Ezzat, M.I.; Hassan, M.; Abdelhalim, M.A.; El-Desoky, A.M.; Mohamed, S.O.; Ezzat, S.M. Immunomodulatory effect of Noni fruit and its isolates: Insights into cell-mediated immune response and inhibition of LPS-induced THP-1 macrophage inflammation. Food Funct. 2021, 12, 3170–3179. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.-H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Jensen, P.Ø.; Jakobsen, T.H.; Phipps, R.; Nielsen, A.K.; Rybtke, M.T.; Tolker-Nielsen, T.; Givskov, M.; Høiby, N.; Ciofu, O. Quorum Sensing and Virulence of Pseudomonas aeruginosa during Lung Infection of Cystic Fibrosis Patients. PLoS ONE 2010, 5, e10115. [Google Scholar] [CrossRef] [Green Version]
- Froes, T.; Baldini, R.L.; Vajda, S.; Castilho, M.S. Structure-based Druggability assessment of anti-virulence targets from pseudomonas aeruginosa. Curr. Protein Pept. Sci. 2019, 20, 1189–1203. [Google Scholar] [CrossRef]
- Cordeiro, L.; Figueiredo, P.; Souza, H.; Sousa, A.; Andrade-Júnior, F.; Barbosa-Filho, J.; Lima, E. Antibacterial and Antibiofilm Activity of Myrtenol against Staphylococcus aureus. Pharmaceuticals 2020, 13, 133. [Google Scholar] [CrossRef] [PubMed]
- Elzeini, H.M.; Ali, A.R.A.A.; Nasr, N.F.; Hassan, M.; Hassan, A.A.M.; Elenany, Y.E. Probiotic capability of novel lactic acid bacteria isolated from worker honey bees gut microbiota. FEMS Microbiol. Lett. 2021, 368, fnab030. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.; Pillai, S.D.; Patil, B.S. Citrus limonoids interfere with Vibrio harveyi cell–cell signalling and biofilm formation by modulating the response regulator LuxO. Microbiology 2011, 157, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynertson, K.A.; Yang, H.; Jiang, B.; Basile, M.J.; Kennelly, E.J. Quantitative analysis of antiradical phenolic constituents from fourteen edible Myrtaceae fruits. Food Chem. 2008, 109, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Siani, A.C.; Nakamura, M.J.; D’Avila, L.A.; Lima, A.M.B. Selective and cost effective protocol to separate bioactive triterpene acids from plant matrices using alkalinized ethanol: Application to leaves of Myrtaceae species. Pharmacogn. Mag. 2015, 11, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saber, F.R.; Abdelbary, G.A.; Salama, M.M.; Saleh, D.O.; Fathy, M.M.; Soliman, F.M. UPLC/QTOF/MS profiling of two Psidium species and the in-vivo hepatoprotective activity of their nano-formulated liposomes. Food Res. Int. 2018, 105, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Mahrous, E.A.; Al-Abd, A.; Salama, M.; Fathy, M.; Soliman, F.; Saber, F.R. Cattleianal and Cattleianone: Two New Meroterpenoids from Psidium cattleianum leaves and their selective antiproliferative action against human carcinoma cells. Molecules 2021, 26, 2891. [Google Scholar] [CrossRef]
- Oyedeji, O.O.; Lawal, O.A.; Shode, F.O.; Oyedeji, A. Chemical Composition and Antibacterial Activity of the Essential Oils of Callistemon citrinus and Callistemon viminalis from South Africa. Molecules 2009, 14, 1990–1998. [Google Scholar] [CrossRef]
- Siddique, S.; Perveen, Z.; Nawaz, S.; Shahzad, K.; Ali, Z. Chemical Composition and Antimicrobial Activities of Essential Oils of Six Species from Family Myrtaceae. J. Essent. Oil Bear. Plants 2015, 18, 950–956. [Google Scholar] [CrossRef]
- Xiang, Y.-Q.; Liu, H.; Zhao, L.-Y.; Xu, Z.-F.; Tan, H.-B.; Qiu, S.-X. Callistemenonone A, a novel dearomatic dibenzofuran-type acylphloroglucinol with antimicrobial activity from Callistemon viminalis. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shehabeldine, A.M.; Ashour, R.M.; Okba, M.M.; Saber, F.R. Callistemon citrinus bioactive metabolites as new inhibitors of methicillin-resistant Staphylococcus aureus biofilm formation. J. Ethnopharmacol. 2020, 254, 112669. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, R.; Hassan, M.; Hashem, R.; Abdel-Sattar, E. Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming multidrug-resistant pathogens. Antibiotics 2021, 10, 811. [Google Scholar] [CrossRef]
- Martínez, O.F.; Cardoso, M.; Ribeiro, S.; Franco, O.L. Recent advances in anti-virulence therapeutic strategies with a focus on dismantling bacterial membrane microdomains, toxin neutralization, quorum-sensing interference and biofilm inhibition. Front. Cell. Infect. Microbiol. 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef]
- Hoge, R.; Pelzer, A.; Rosenau, F.; Wilhelm, S.; Duesseldorf, H.-H.-U. Weapons of a pathogen: Proteases and their role in vir-ulence of Pseudomonas aeruginosa. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 383–395. [Google Scholar]
- Sarkisian, S.A.; Janssen, M.J.; Matta, H.; Henry, G.E.; Laplante, K.L.; Rowley, D.C. Inhibition of Bacterial Growth and Biofilm Production by Constituents from Hypericum spp. Phytother. Res. 2011, 26, 1012–1016. [Google Scholar] [CrossRef] [Green Version]
- Biessy, A.; Filion, M. Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions. Metabolites 2021, 11, 182. [Google Scholar] [CrossRef]
- El-Sayed, N.; Samir, R.; Abdel-Hafez, L.J.M.; Ramadan, M. Olive Leaf Extract Modulates Quorum Sensing Genes and Biofilm Formation in Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics 2020, 9, 526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-W.; Luo, H.-Z.; Jiang, H.; Jian, T.-K.; Chen, Z.-Q.; Jia, A.-Q. Hordenine: A Novel Quorum Sensing Inhibitor and Antibiofilm Agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 2018, 66, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Froes, T.; Nicastro, G.G.; Pereira, T.D.O.; Carneiro, K.D.O.; Reis, I.M.A.; Conceição, R.S.; Branco, A.; Ifa, D.R.; Baldini, R.L.; Castilho, M.S. Calycopterin, a major flavonoid from Marcetia latifolia, modulates virulence-related traits in Pseudomonas aeruginosa. Microb. Pathog. 2020, 144, 104142. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Liu, H.; Zhao, L.; Yuan, Y.; Li, B.; Jiang, Y.; Gong, L.; Qiu, S. Structure-activity relationships and optimization of acyclic acylphloroglucinol analogues as novel antimicrobial agents. Eur. J. Med. Chem. 2017, 125, 492–499. [Google Scholar] [CrossRef]
- Ardizzoni, A.; Hansen, H.; Dombernowsky, P.; Gamucci, T.; Kaplan, S.; Postmus, P.; Giaccone, G.; Schaefer, B.; Wanders, J.; Verweij, J. Topotecan, a new active drug in the second-line treatment of small-cell lung cancer: A phase II study in patients with refractory and sensitive disease. The European Organization for Research and Treatment of Cancer Early Clinical Studies Group and New Drug Development Office, and the Lung Cancer Cooperative Group. J. Clin. Oncol. 1997, 15, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.A.; Shaldam, M.; Eldamasi, D. Curtailing Quorum Sensing in Pseudomonas aeruginosa by Sitagliptin. Curr. Microbiol. 2020, 77, 1051–1060. [Google Scholar] [CrossRef]
- Jander, G.; Rahme, L.G.; Ausubel, F.M. Positive Correlation between Virulence of Pseudomonas aeruginosa Mutants in Mice and Insects. J. Bacteriol. 2000, 182, 3843–3845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, M.A.; Hager, R.; Reheem, F.A.-E.; Mahmoud, E.E.; Samir, T.; Moawad, S.S.; Hefzy, E.M. A Study of the Virulence Traits of Carbapenem-Resistant Klebsiella pneumoniae Isolates in a Galleria mellonella Model. Microb. Drug Resist. 2019, 25, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.A.F.; Moawad, S.S.; Hefzy, E.M. In vivo activity of co-trimoxazole combined with colistin against Acinetobacter baumannii producing OXA-23 in a Galleria mellonella model. J. Med. Microbiol. 2019, 68, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Coote, P.J. Chapter 2—Utility of Greater Wax Moth Larva (Galleria mellonella) for Evaluating the Toxicity and Efficacy of New Antimicrobial Agents. In Advances in Applied Microbiology; Laskin, A.I., Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 25–53. [Google Scholar]
- Allegra, E.; Titball, R.W.; Carter, J.; Champion, O.L. Galleria mellonella larvae allow the discrimination of toxic and non-toxic chemicals. Chemosphere 2018, 198, 469–472. [Google Scholar] [CrossRef]
- Tharmalingam, N.; Khader, R.; Fuchs, B.B.; Mylonakis, E. The Anti-virulence Efficacy of 4-(1,3-Dimethyl-2,3-Dihydro-1H-Benzimidazol-2-yl)Phenol Against Methicillin-Resistant Staphylococcus aureus. Front. Microbiol. 2019, 10, 1557. [Google Scholar] [CrossRef] [Green Version]
- Bloor, S.J. Antiviral Phloroglucinols from New Zealand Kunzea Species. J. Nat. Prod. 1992, 55, 43–47. [Google Scholar] [CrossRef]
- Salem, M.A.; El-Shiekh, R.A.; Hashem, R.A.; Hassan, M. In vivo Antibacterial Activity of Star Anise (Illicium verum Hook.) Extract Using Murine MRSA skin infection model in relation to its metabolite profile. Infect. Drug Resist. 2021, 14, 33–48. [Google Scholar] [CrossRef]
- Sultan, A.M.; Nabiel, Y. Tube method and Congo red agar versus tissue culture plate method for detection of biofilm production by uropathogens isolated from midstream urine: Which one could be better? Afr. J. Clin. Exp. Microbiol. 2018, 20, 60. [Google Scholar] [CrossRef] [Green Version]
- Nithya, C.; Devi, M.G.; Pandian, S.K. A novel compound from the marine bacteriumBacillus pumilusS6-15 inhibits biofilm formation in Gram-positive and Gram-negative species. Biofouling 2011, 27, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Karatuna, O.; Yagci, A. Analysis of quorum sensing-dependent virulence factor production and its relationship with antimicrobial susceptibility in Pseudomonas aeruginosa respiratory isolates. Clin. Microbiol. Infect. 2010, 16, 1770–1775. [Google Scholar] [CrossRef] [Green Version]
- Vijayaraghavan, P.; Vincent, S.G.P. A simple method for the detection of protease activity on agar plate using bromocresolgreen dye. J. Biochem. Technol. 2013, 4, 628–630. [Google Scholar]
- Peleg, A.Y.; Jara, S.; Monga, D.; Eliopoulos, G.M.; Moellering, R.C.; Mylonakis, E. Galleria mellonella as a Model System To Study Acinetobacter baumannii Pathogenesis and Therapeutics. Antimicrob. Agents Chemother. 2009, 53, 2605–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 26th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
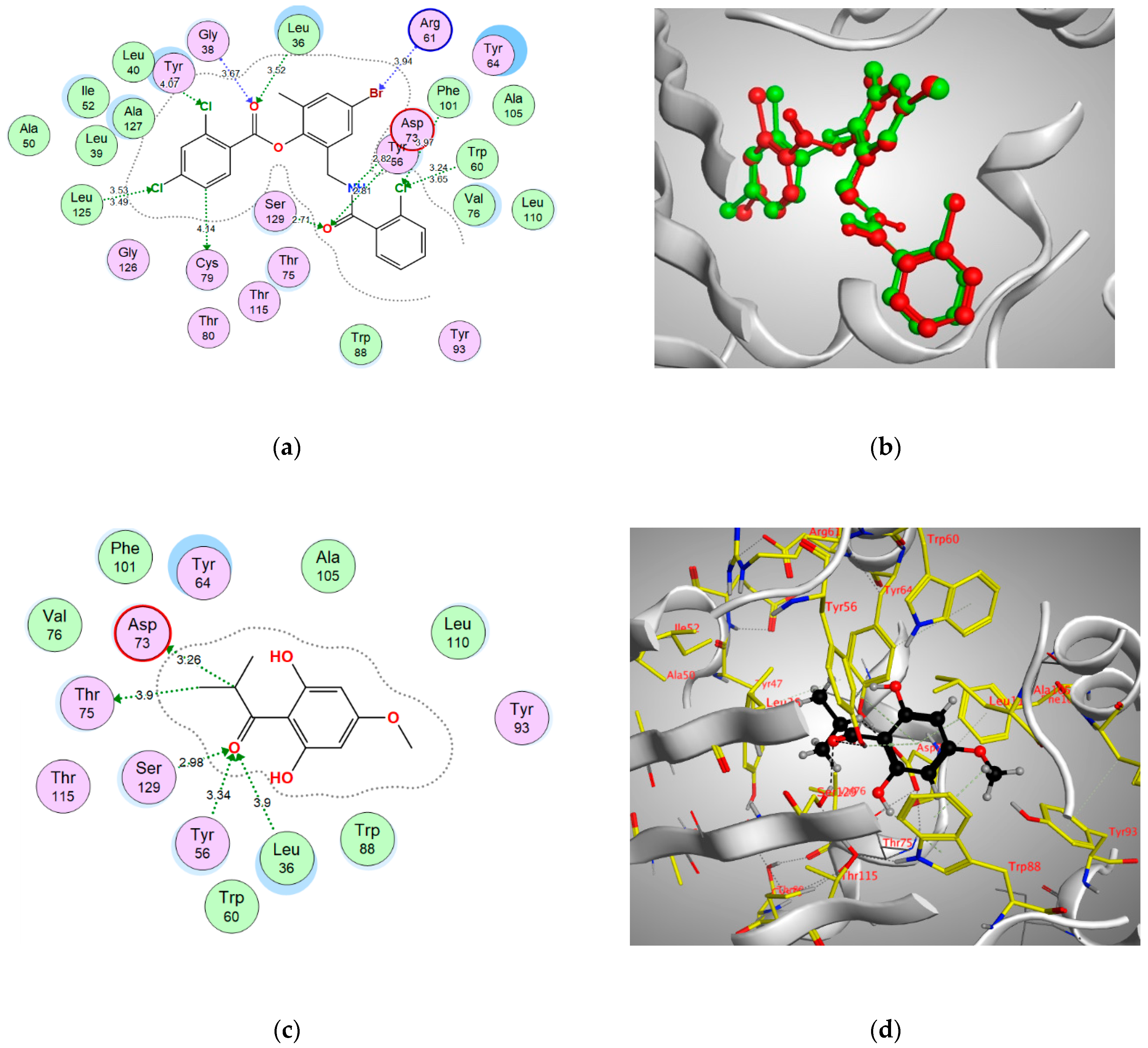
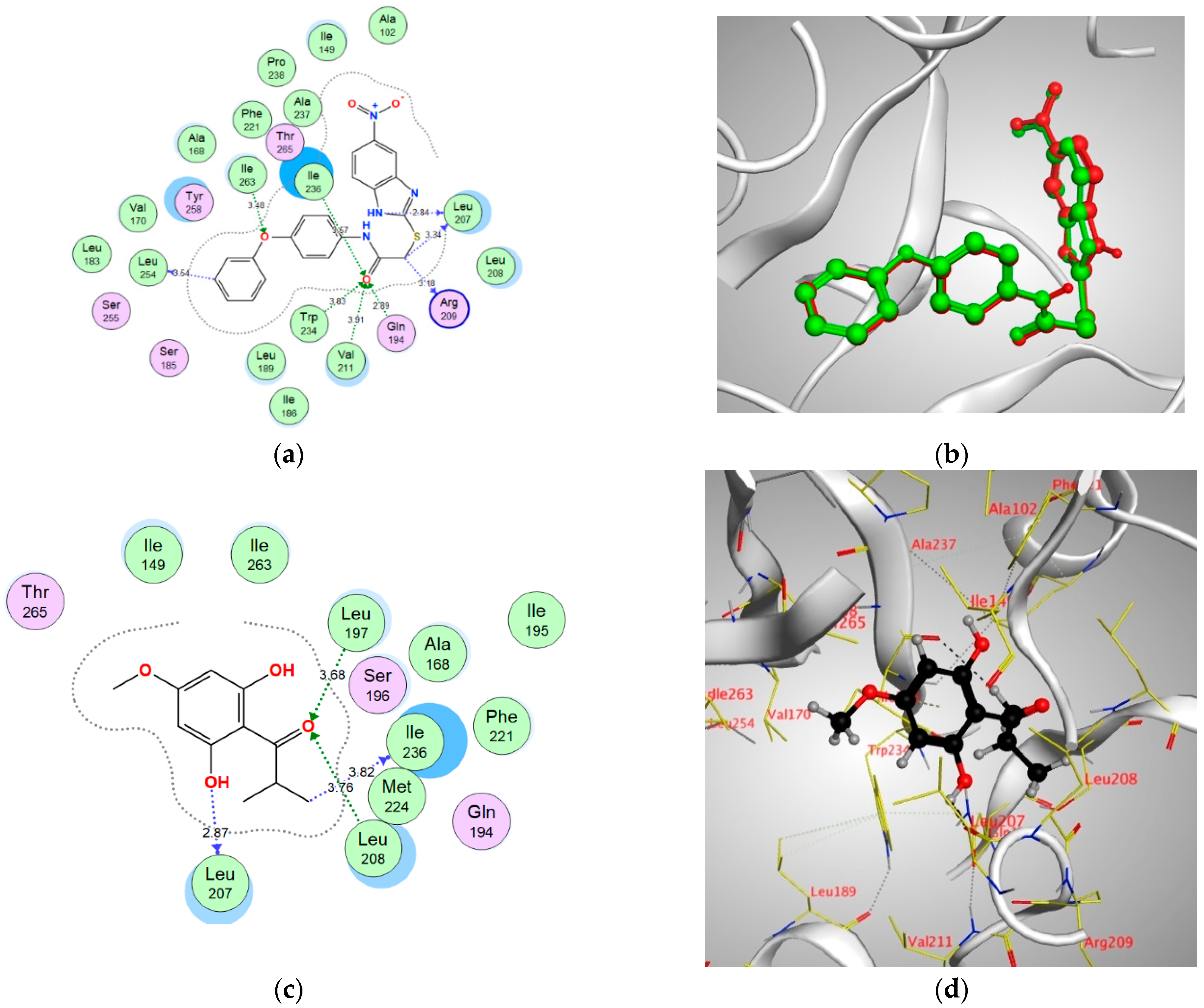
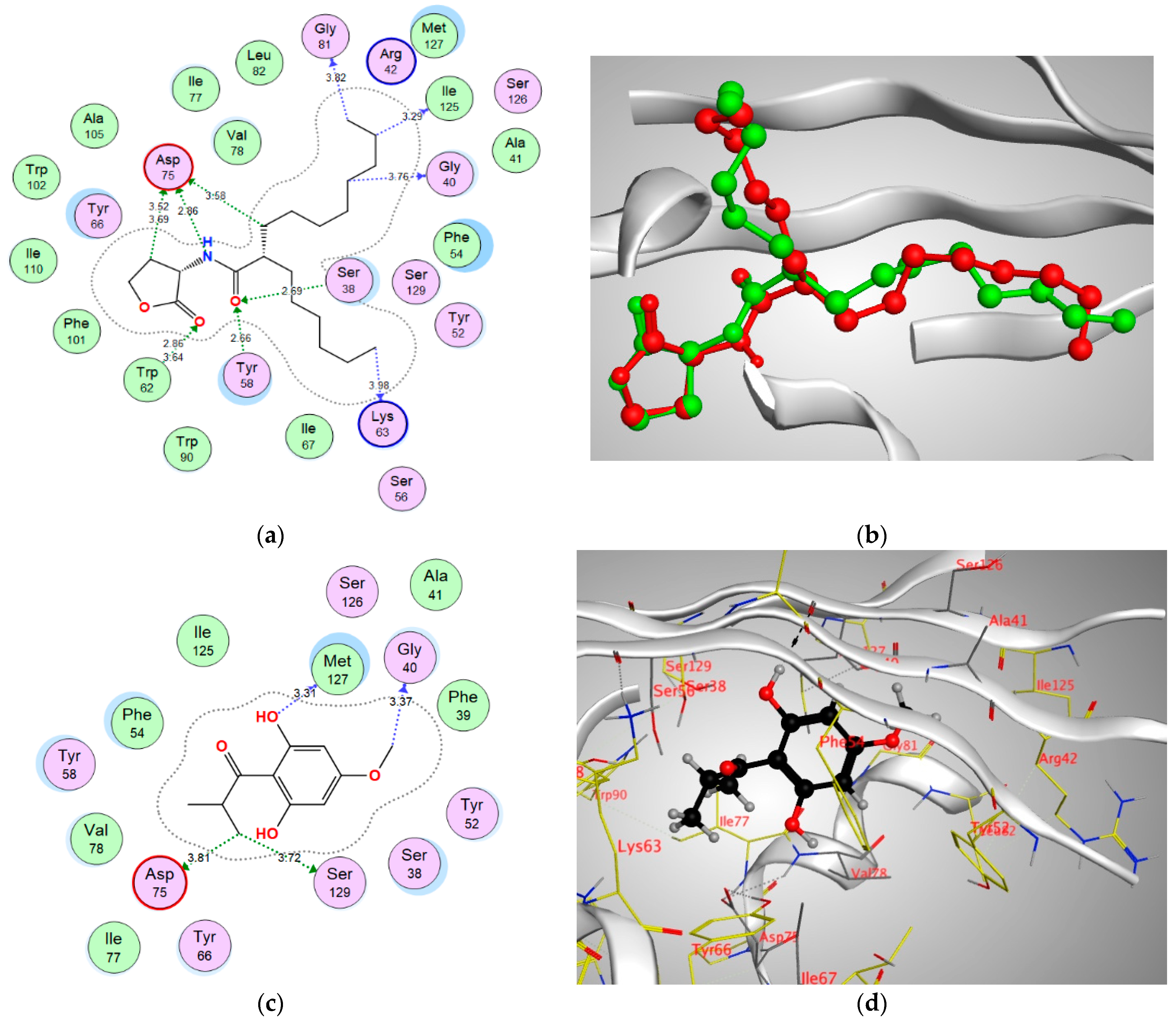
| Enzyme | Binding Score (kcal/mol) | Amino Acids | Interacting Groups | Type of Interaction | Bond Length |
|---|---|---|---|---|---|
| LasR | −12.6517 | Leu36 | O (C=O) | H-bond acceptor | 3.90 |
| Tyr56 | O (C=O) | H-bond acceptor | 3.34 | ||
| Asp73 | CH | H-bond (non-conventional) | 3.26 | ||
| Thr75 | CH3 | H-bond (non-conventional) | 3.90 | ||
| Ser129 | O (C=O) | H-bond acceptor | 2.98 | ||
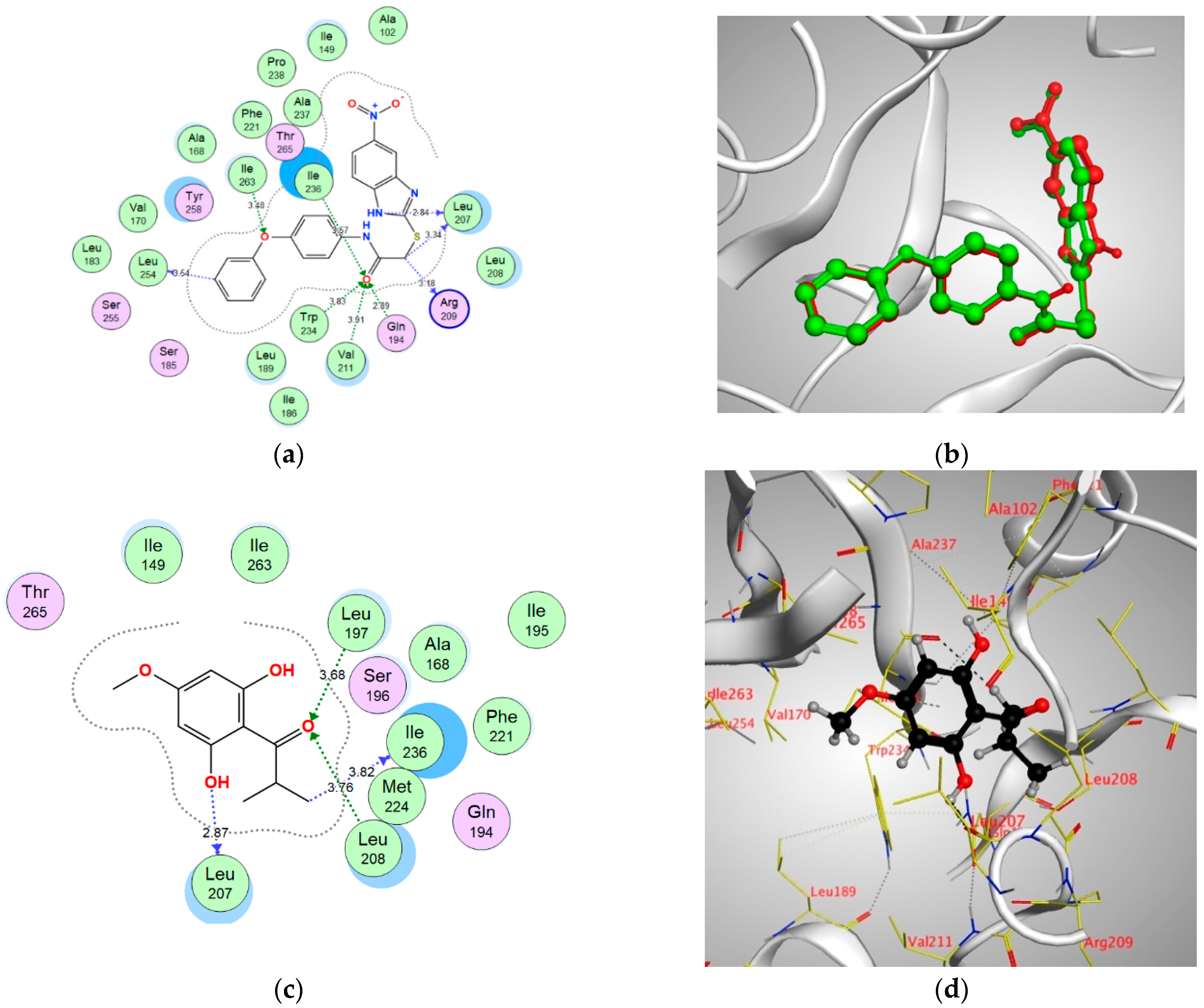
| MyfR/PqsR | −9.2847 | Leu197 | O (C=O) | H-bond acceptor | 3.68 |
| Leu207 | OH | H-bond donor | 2.87 | ||
| Leu208 | O (C=O) | H-bond acceptor | 3.76 | ||
| Ile236 | CH3 | H-bond (non-conventional) | 3.82 | ||
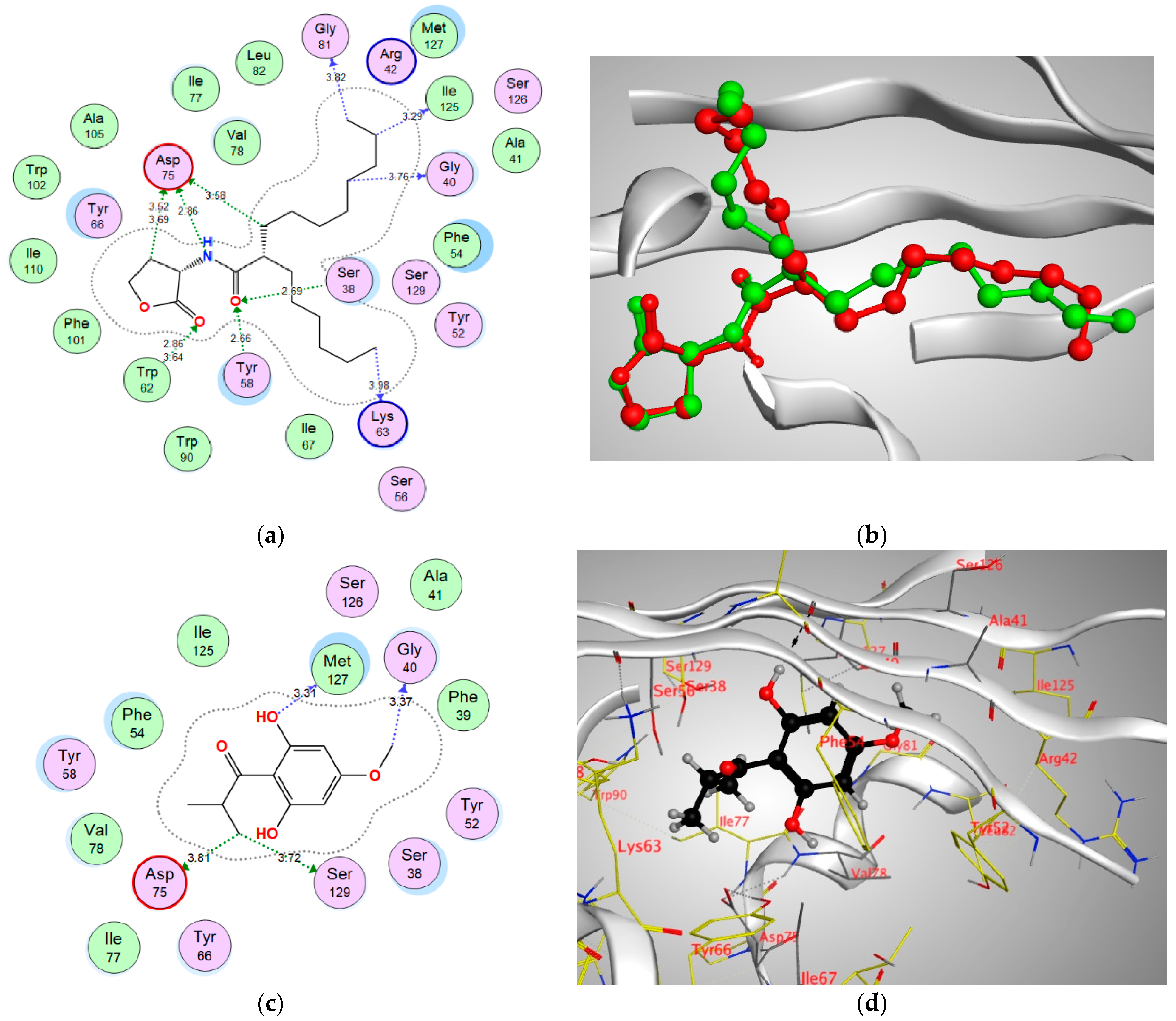
| QscR | −13.3577 | Gly40 | CH3 | H-bond (non-conventional) | 3.37 |
| Asp75 | CH3 | H-bond (non-conventional) | 3.81 | ||
| Met127 | OH | H-bond donor | 3.31 | ||
| Ser129 | CH3 | H-bond (non-conventional) | 3.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, M.M.; Hassan, M.; Moawad, S.S.; Okba, M.M.; Ashour, R.M.; Fayek, N.M.; Saber, F.R. Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 907. https://doi.org/10.3390/antibiotics10080907
Ismail MM, Hassan M, Moawad SS, Okba MM, Ashour RM, Fayek NM, Saber FR. Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics. 2021; 10(8):907. https://doi.org/10.3390/antibiotics10080907
Chicago/Turabian StyleIsmail, Maha M., Mariam Hassan, Sawsan S. Moawad, Mona M. Okba, Rehab M. Ashour, Nesrin M. Fayek, and Fatema R. Saber. 2021. "Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa" Antibiotics 10, no. 8: 907. https://doi.org/10.3390/antibiotics10080907
APA StyleIsmail, M. M., Hassan, M., Moawad, S. S., Okba, M. M., Ashour, R. M., Fayek, N. M., & Saber, F. R. (2021). Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics, 10(8), 907. https://doi.org/10.3390/antibiotics10080907