
Synergy between Phage Sb-1 and Oxacillin against Methicillin-Resistant Staphylococcus aureus

Abstract
1. Introduction
2. Results and Discussion
3. Methods
3.1. Description of S. aureus Strains
3.2. Description of Phage Sb-1
3.3. Time-Kill Assays
3.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Molina, K.C.; Morrisette, T.; Miller, M.A.; Huang, V.; Fish, D.N. The Emerging Role of β-Lactams in the Treatment of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Torres-Barceló, C.; Hochberg, M.E. Evolutionary Rationale for Phages as Complements of Antibiotics. Trends Microbiol. 2016, 24, 249–256. [Google Scholar] [CrossRef]
- Tagliaferri, T.L.; Jansen, M.; Horz, H.-P. Fighting Pathogenic Bacteria on Two Fronts: Phages and Antibiotics as Combined Strategy. Front. Cell. Infect. Microbiol. 2019, 9, 22. [Google Scholar] [CrossRef]
- Kirby, A.E. Synergistic Action of Gentamicin and Bacteriophage in a Continuous Culture Population of Staphylococcus aureus. PLoS ONE 2012, 7, e51017. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage Selection Restores Antibiotic Sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef]
- Rahman, M.; Kim, S.; Kim, S.M.; Seol, S.Y.; Kim, J. Characterization of Induced Staphylococcus aureus Bacteriophage SAP-26 and Its Anti-Biofilm Activity with Rifampicin. Biofouling 2011, 27, 1087–1093. [Google Scholar] [CrossRef]
- Chhibber, S.; Kaur, T. Sandeep Kaur Co-Therapy Using Lytic Bacteriophage and Linezolid: Effective Treatment in Eliminating Methicillin Resistant Staphylococcus aureus (MRSA) from Diabetic Foot Infections. PLoS ONE 2013, 8, e56022. [Google Scholar] [CrossRef] [PubMed]
- Kebriaei, R.; Lev, K.; Morrisette, T.; Stamper, K.C.; Abdul-Mutakabbir, J.C.; Lehman, S.M.; Morales, S.; Rybak, M.J. Bacteriophage-Antibiotic Combination Strategy: An Alternative against Methicillin-Resistant Phenotypes of Staphylococcus aureus. Antimicrob. Agents Chemother. 2020, 64, e00461-20. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, D.; Taha, M.; Yi, Q.; Ramirez-Arcos, S.; Diallo, J.-S.; Carli, A.; Abdelbary, H. Does Treatment Order Matter? Investigating the Ability of Bacteriophage to Augment Antibiotic Activity against Staphylococcus aureus Biofilms. Front. Microbiol. 2018, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tkhilaishvili, T.; Trampuz, A.; Gonzalez Moreno, M. Evaluation of Staphylococcal Bacteriophage Sb-1 as an Adjunctive Agent to Antibiotics Against Rifampin-Resistant Staphylococcus aureus Biofilms. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Dickey, J.; Perrot, V. Adjunct Phage Treatment Enhances the Effectiveness of Low Antibiotic Concentration against Staphylococcus aureus Biofilms in Vitro. PLoS ONE 2019, 14, e0209390. [Google Scholar] [CrossRef]
- Berryhill, B.A.; Huseby, D.L.; McCall, I.C.; Hughes, D.; Levin, B.R. Evaluating the Potential Efficacy and Limitations of a Phage for Joint Antibiotic and Phage Therapy of Staphylococcus aureus Infections. Proc. Natl. Acad. Sci. USA 2021, 118, e2008007118. [Google Scholar] [CrossRef]
- Jansen, M.; Wahida, A.; Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.M.; Ritter, K.; Horz, H.-P. Enhanced Antibacterial Effect of the Novel T4-like Bacteriophage KARL-1 in Combination with Antibiotics against Multi-Drug Resistant Acinetobacter baumannii. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.; Jansen, M.; Krüttgen, A.; Buhl, E.M.; Horz, H.-P. Tackling Intrinsic Antibiotic Resistance in Serratia marcescens with a Combination of Ampicillin/Sulbactam and Phage SALSA. Antibiotics 2020, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Palma Medina, L.M.; Stobernack, T.; Glasner, C.; de Jong, A.; Utari, P.; Setroikromo, R.; Quax, W.J.; Otto, A.; Becher, D.; et al. Exoproteome Heterogeneity among Closely Related Staphylococcus aureus T437 Isolates and Possible Implications for Virulence. J. Proteome Res. 2019, 18, 2859–2874. [Google Scholar] [CrossRef]
- Naorem, R.S.; Blom, J.; Fekete, C. Genome-Wide Comparison of Four MRSA Clinical Isolates from Germany and Hungary. PeerJ 2021, 9, e10185. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, J.; Munder, A.; Neugebauer, J.; Davenport, C.F.; Stanke, F.; Larbig, K.D.; Heeb, S.; Schöck, U.; Pohl, T.M.; Wiehlmann, L.; et al. Genome Diversity of Pseudomonas aeruginosa PAO1 Laboratory Strains. J. Bacteriol. 2010, 192, 1113–1121. [Google Scholar] [CrossRef]
- Hobman, J.L.; Penn, C.W.; Pallen, M.J. Laboratory Strains of Escherichia coli: Model Citizens or Deceitful Delinquents Growing Old Disgracefully? Mol. Microbiol. 2007, 64, 881–885. [Google Scholar] [CrossRef]
- Kvachadze, L.; Balarjishvili, N.; Meskhi, T.; Tevdoradze, E.; Skhirtladze, N.; Pataridze, T.; Adamia, R.; Topuria, T.; Kutter, E.; Rohde, C.; et al. Evaluation of Lytic Activity of Staphylococcal Bacteriophage Sb-1 against Freshly Isolated Clinical Pathogens: Staphylococcal Bacteriophages Sb-1. Microb. Biotechnol. 2011, 4, 643–650. [Google Scholar] [CrossRef]
- Łobocka, M.; Hejnowicz, M.S.; Dąbrowski, K.; Gozdek, A.; Kosakowski, J.; Witkowska, M.; Ulatowska, M.I.; Weber-Dąbrowska, B.; Kwiatek, M.; Parasion, S.; et al. Genomics of Staphylococcal Twort-like Phages—Potential Therapeutics of the Post-Antibiotic Era. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2012; Volume 83, pp. 143–216. ISBN 9780123944382. [Google Scholar]
- Tevdoradze, E.; Kvachadze, L.; Kutateladze, M.; Stewart, C.R. Bactericidal Genes of Staphylococcal Bacteriophage Sb-1. Curr. Microbiol. 2014, 68, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Sergueev, K.V.; Filippov, A.A.; Farlow, J.; Su, W.; Kvachadze, L.; Balarjishvili, N.; Kutateladze, M.; Nikolich, M.P. Correlation of Host Range Expansion of Therapeutic Bacteriophage Sb-1 with Allele State at a Hypervariable Repeat Locus. Appl. Environ. Microbiol. 2019, 85, e01209-19. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, W.N.; Concepción-Acevedo, J.; Park, T.; Andleeb, S.; Bull, J.J.; Levin, B.R. Synergy and Order Effects of Antibiotics and Phages in Killing Pseudomonas aeruginosa Biofilms. PLoS ONE 2017, 12, e0168615. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, J.; Larsen, M.; Kilstrup, M.; Nielsen, M. Metagenomic Analysis of Therapeutic PYO Phage Cocktails from 1997 to 2014. Viruses 2017, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- McCallin, S.; Sacher, J.C.; Zheng, J.; Chan, B.K. Current State of Compassionate Phage Therapy. Viruses 2019, 11, 343. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Liu, W.-T.; Chen, E.-Z.; Yang, L.; Peng, C.; Wang, Q.; Xu, Z.; Chen, D.-Q. Emerging Resistance Mechanisms for 4 Types of Common Anti-MRSA Antibiotics in Staphylococcus aureus: A Comprehensive Review. Microb. Pathog. 2021, 156, 104915. [Google Scholar] [CrossRef]
- Ye, Y.; Jiang, Q.; Wu, Q.; Zhang, J.; Lu, J.; Lin, L. The Characterization and Comparison of Staphylococcus aureus by Antibiotic Susceptibility Testing, Enterobacterial Repetitive Intergenic Consensus–Polymerase Chain Reaction, and Random Amplified Polymorphic DNA–Polymerase Chain Reaction. Foodborne Pathog. Dis. 2012, 9, 168–171. [Google Scholar] [CrossRef]
- Lehot, J.J.; Reverdy, M.E.; Etienne, J.; Corot, C.; Nervi, C.; Fleurette, J.; Estanove, S. Oxacillin and Tobramycin Serum Levels during Cardiopulmonary Bypass. J. Cardiothorac. Anesth. 1989, 3, 163–167. [Google Scholar] [CrossRef]
- Fitzgerald, R.H.; Kelly, P.J.; Snyder, R.J.; Washington, J.A. Penetration of Methicillin, Oxacillin, and Cephalothin into Bone and Synovial Tissues. Antimicrob. Agents Chemother. 1978, 14, 723–726. [Google Scholar] [CrossRef][Green Version]
- Uchiyama, J.; Shigehisa, R.; Nasukawa, T.; Mizukami, K.; Takemura-Uchiyama, I.; Ujihara, T.; Murakami, H.; Imanishi, I.; Nishifuji, K.; Sakaguchi, M.; et al. Piperacillin and Ceftazidime Produce the Strongest Synergistic Phage–Antibiotic Effect in Pseudomonas aeruginosa. Arch. Virol. 2018, 163, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barceló, C.; Gurney, J.; Gougat-Barberá, C.; Vasse, M.; Hochberg, M.E. Transient Negative Effects of Antibiotics on Phages Do Not Jeopardise the Advantages of Combination Therapies. FEMS Microbiol. Ecol. 2018, 94, fiy107. [Google Scholar] [CrossRef] [PubMed]
- Gu Liu, C.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-Antibiotic Synergy Is Driven by a Unique Combination of Antibacterial Mechanism of Action and Stoichiometry. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]



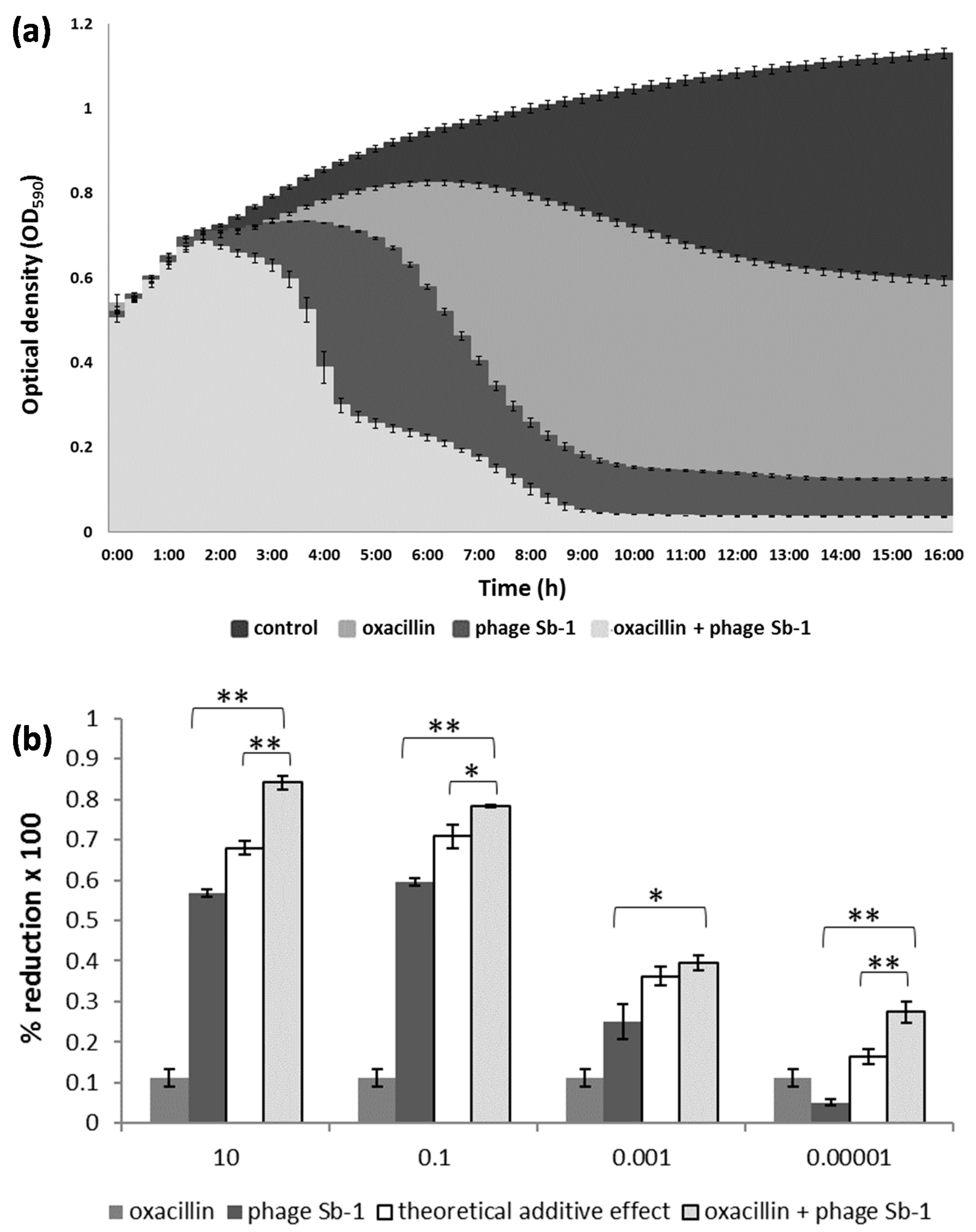{kind=link}
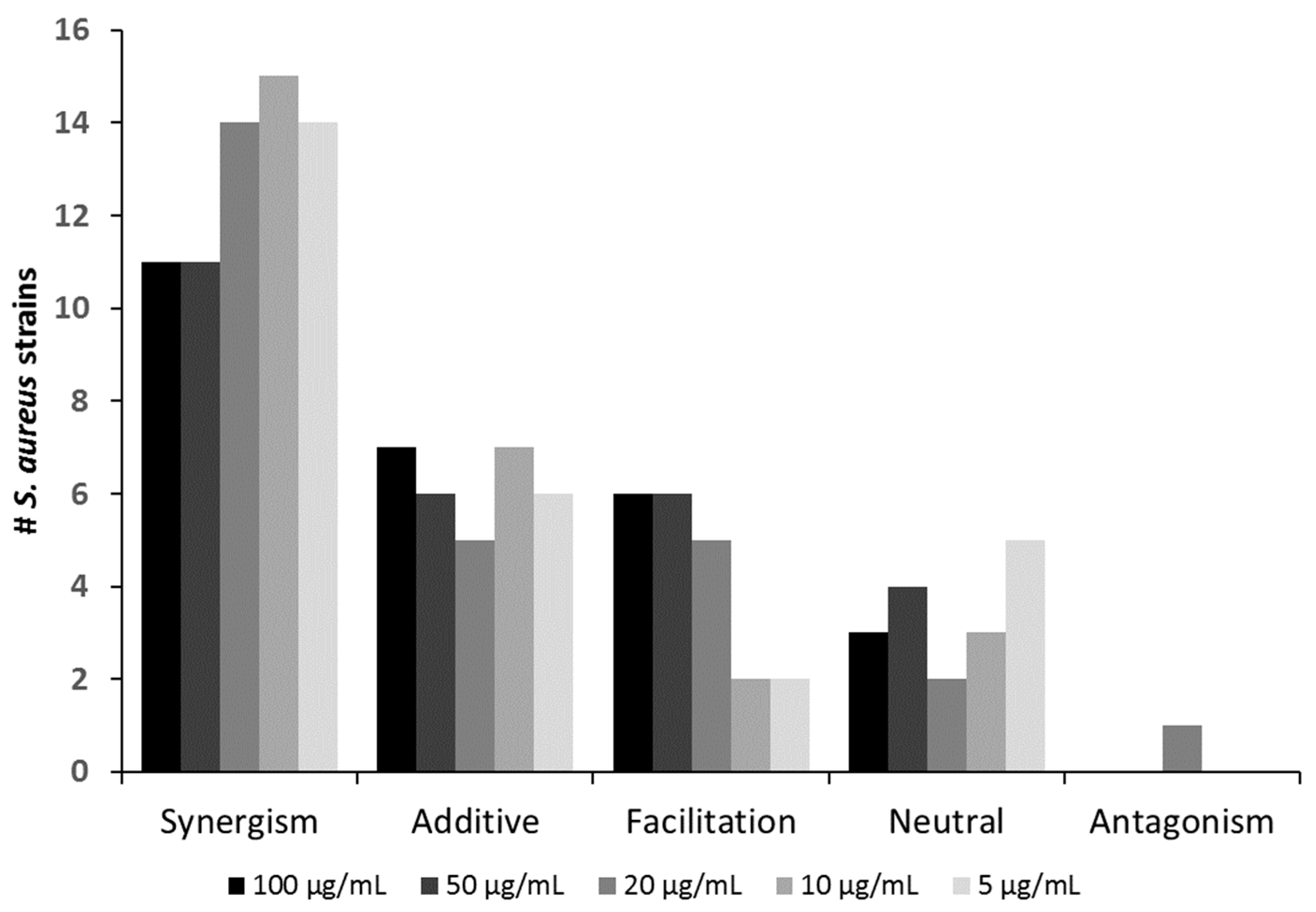{kind=link}
| Oxacillin (µg/mL) Effect | 100 | 50 | 20 | 10 | 5 |
|---|---|---|---|---|---|
| Synergism | SA7, SA9, SA11, SA12, SA15, SA16, SA18, SA20, SA22, SA24, SA ATCC 27660 | SA7, SA9, SA11, SA12, SA15, SA16, SA18, SA19, SA20, SA24, SA ATCC 27660 | SA1, SA6, SA7, SA8, SA9, SA11, SA12, SA15, SA16, SA18, SA19, SA20, SA24, SA ATCC 27660 | SA1, SA6, SA7, SA8, SA9, SA11, SA15, SA16, SA17, SA18, SA19, SA20, SA22,SA24, SA ATCC 27660 | SA1, SA6, SA7, SA8, SA9, SA11, SA16, SA18, SA19, SA20,SA24, SA ATCC14154, SA ATCC 27660, SA ATCC 29132 |
| Additive | SA1, SA6, SA8, SA13, SA14, SA19, SA23 | SA1, SA6, SA8, SA13, SA14, SA23 | SA3, SA4, SA5, SA13, SA23 | SA3, SA4, SA5, SA13, SA23, SA ATCC 14154, SA ATCC 29132 | SA3, SA4, SA5, SA13, SA15, SA23 |
| Facilitation | SA3, SA4, SA5, SA ATCC 14154, SA ATCC 25923, SA ATCC 29132 | SA3, SA4, SA5, SA ATCC 14154, SA ATCC 25923, SA ATCC 29132 | SA14, SA17, SA ATCC 14154, SA ATCC 25923, SA ATCC 29132 | SA14, SA ATCC 25923 | SA14, SA ATCC 25923 |
| Neutral | SA2, SA17, SA21 | SA2, SA17, SA21, SA22 | SA2, SA22 | SA2, SA12, SA21 | SA2, SA12, SA17, SA21, SA22 |
| Antagonism | SA21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, K.; Pier, W.; Krüttgen, A.; Horz, H.-P. Synergy between Phage Sb-1 and Oxacillin against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2021, 10, 849. https://doi.org/10.3390/antibiotics10070849
Simon K, Pier W, Krüttgen A, Horz H-P. Synergy between Phage Sb-1 and Oxacillin against Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2021; 10(7):849. https://doi.org/10.3390/antibiotics10070849
Chicago/Turabian StyleSimon, Kevin, Wolfgang Pier, Alex Krüttgen, and Hans-Peter Horz. 2021. "Synergy between Phage Sb-1 and Oxacillin against Methicillin-Resistant Staphylococcus aureus" Antibiotics 10, no. 7: 849. https://doi.org/10.3390/antibiotics10070849
APA StyleSimon, K., Pier, W., Krüttgen, A., & Horz, H.-P. (2021). Synergy between Phage Sb-1 and Oxacillin against Methicillin-Resistant Staphylococcus aureus. Antibiotics, 10(7), 849. https://doi.org/10.3390/antibiotics10070849