
Volatile Organic Compounds (VOCs) Produced by Gluconobacter cerinus and Hanseniaspora osmophila Displaying Control Effect against Table Grape-Rot Pathogens

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results
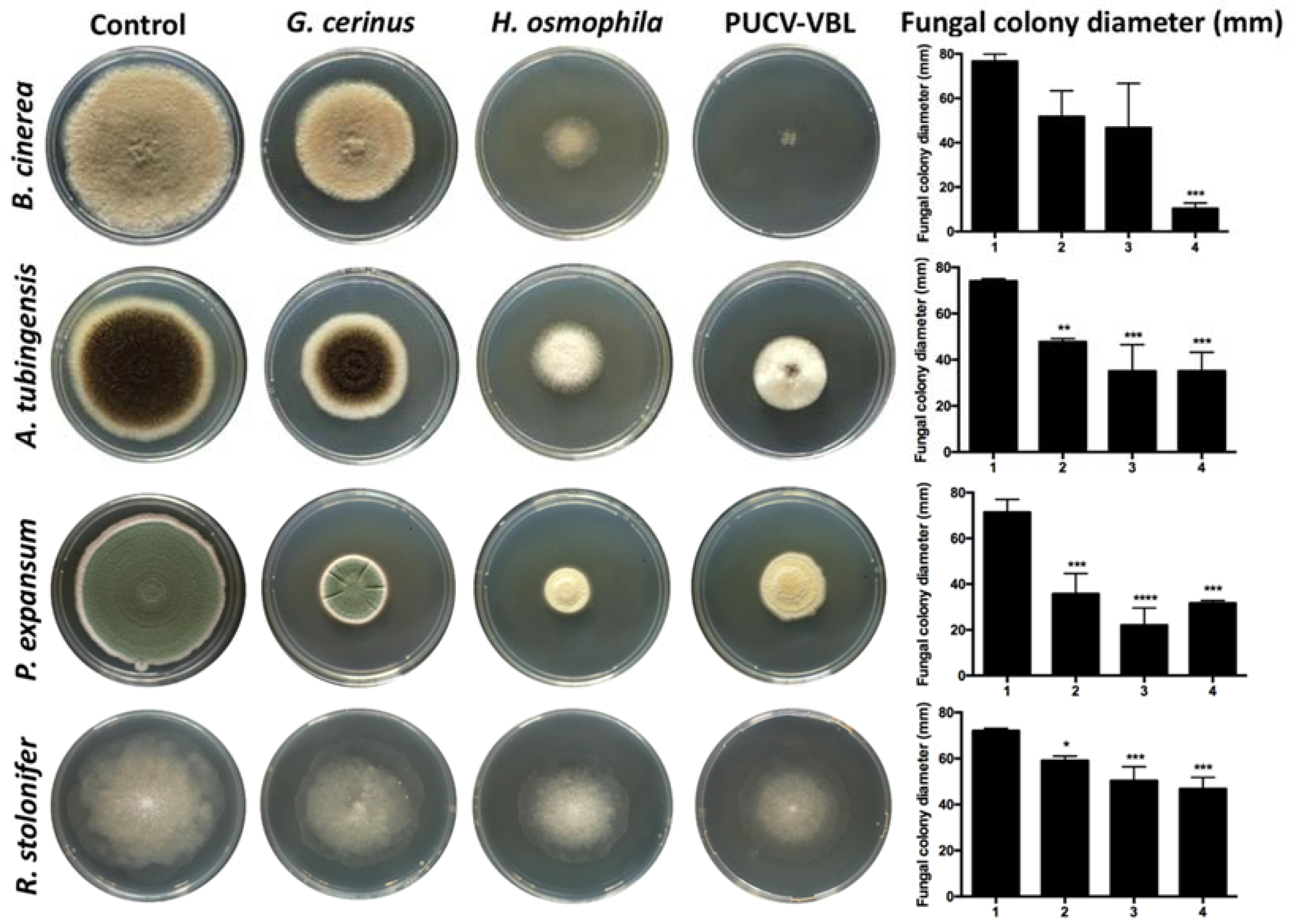
2.1. In Vitro Assay of VOC Production
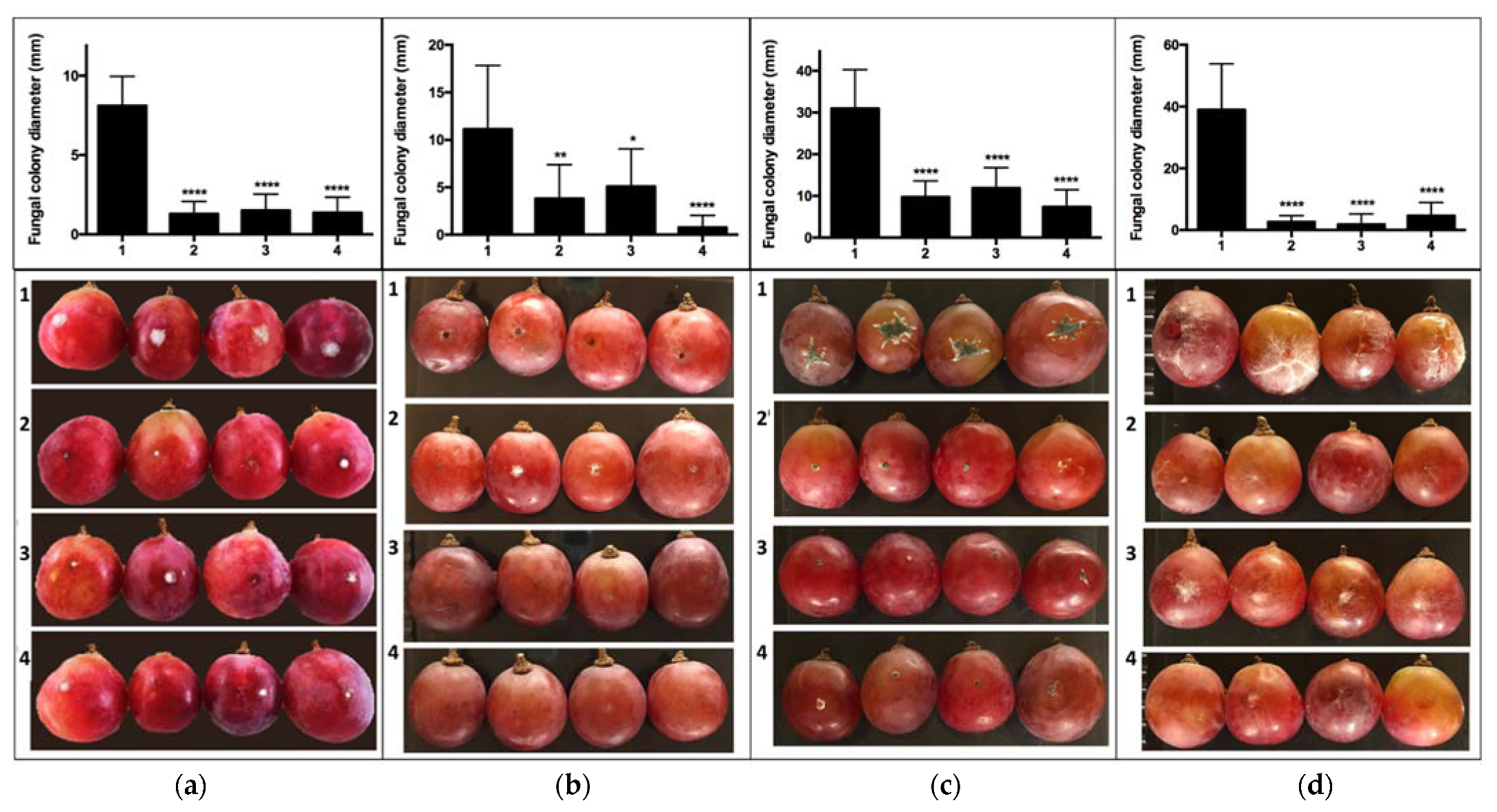
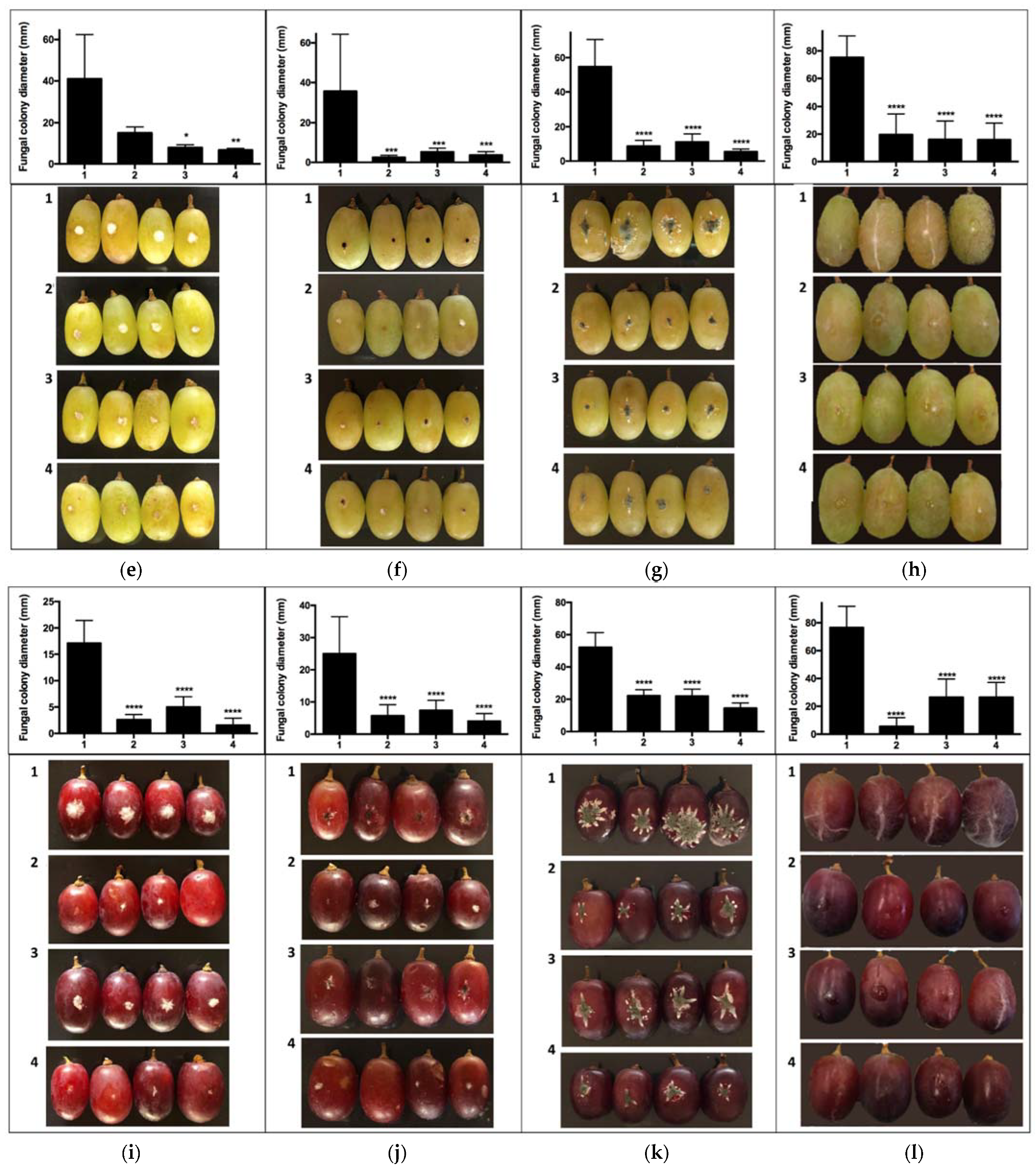
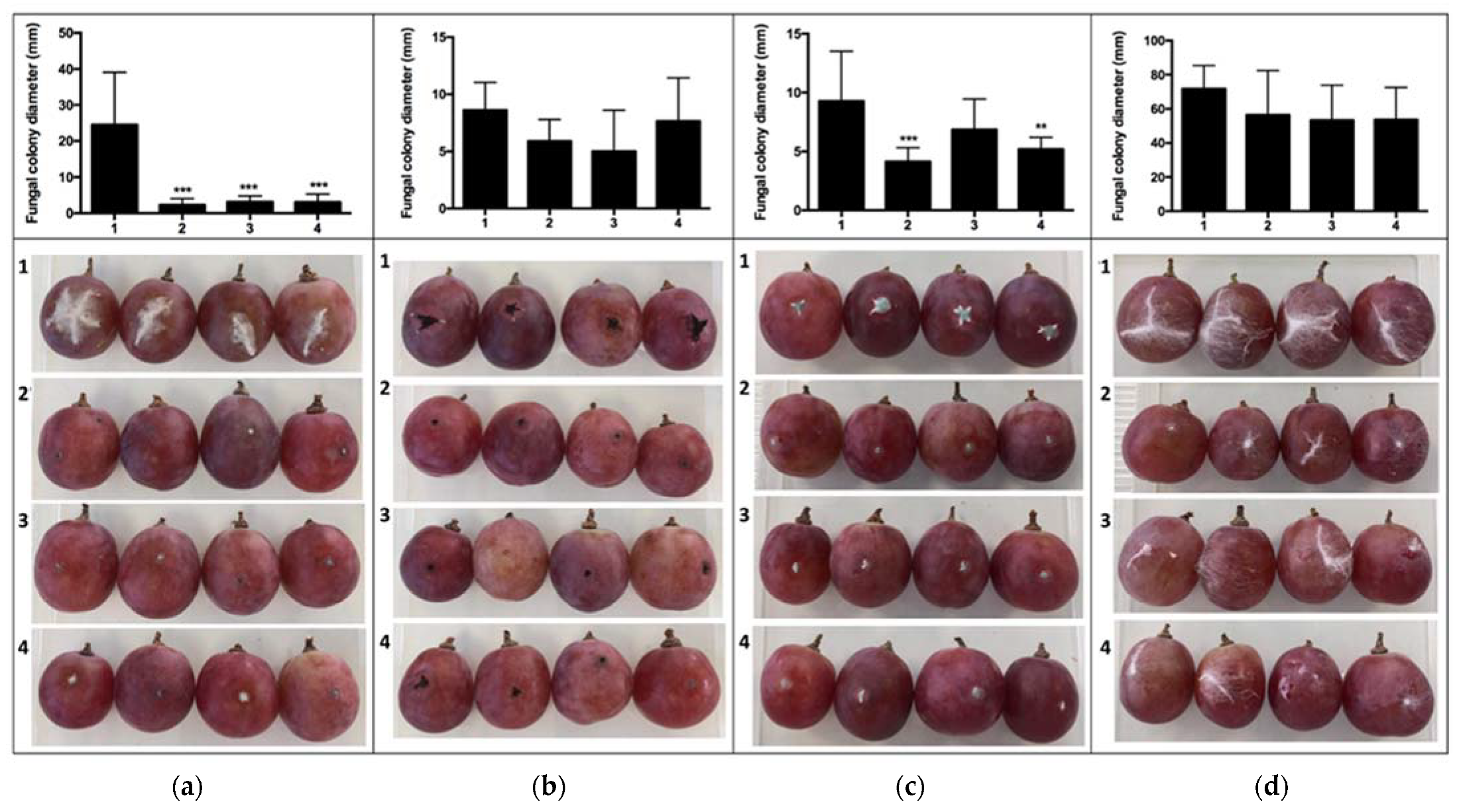
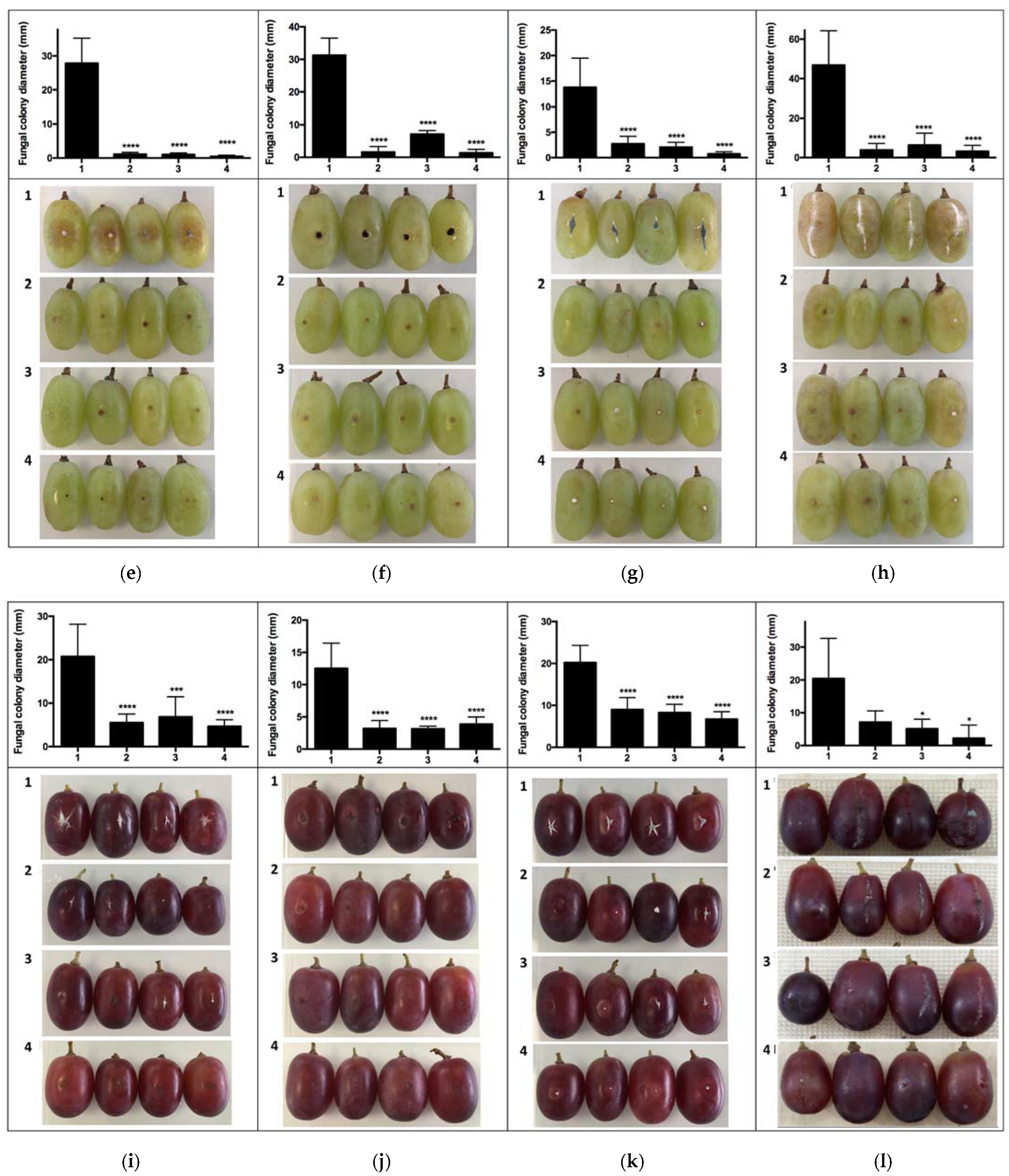
2.2. In Vivo Assays of VOC
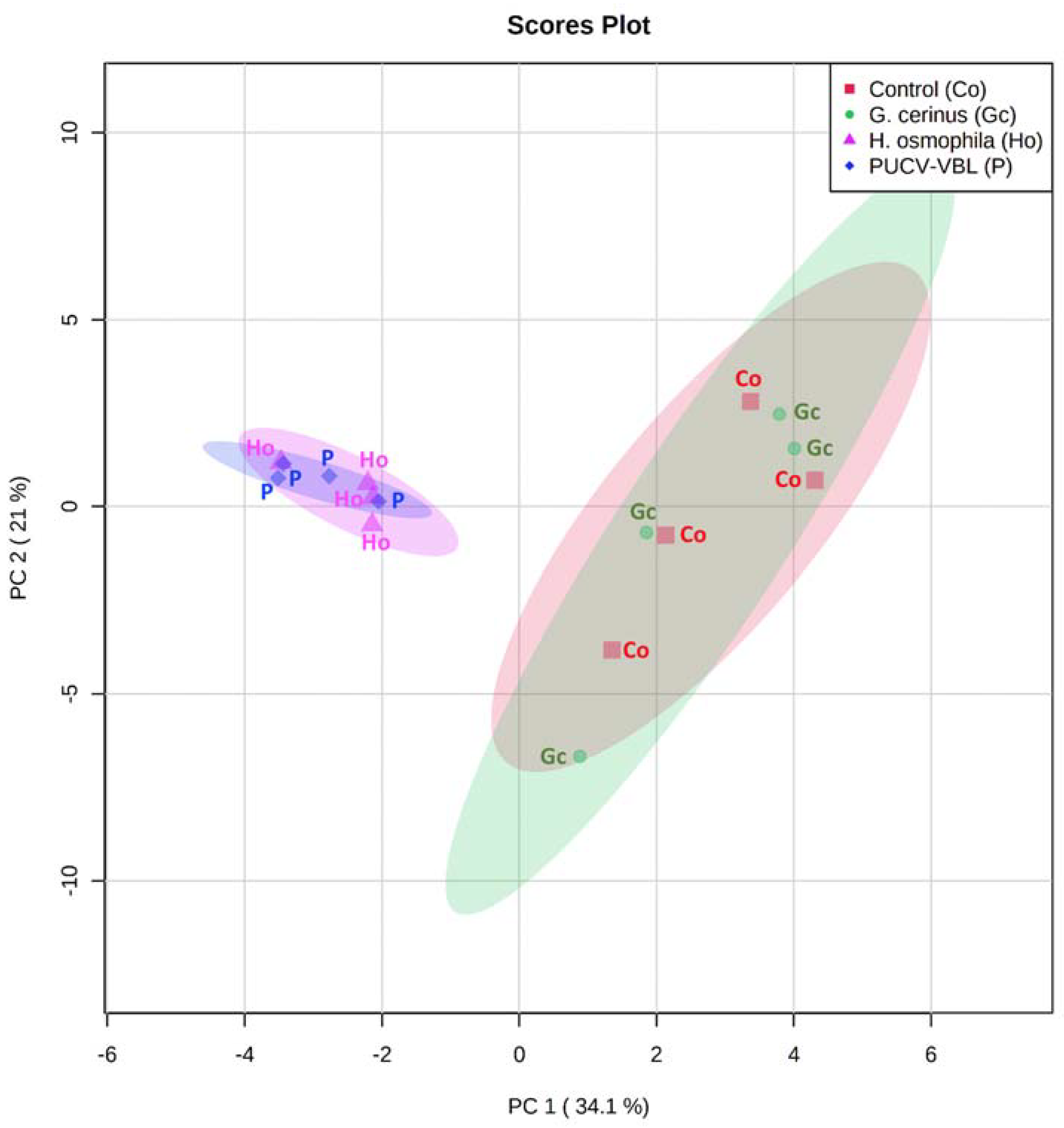
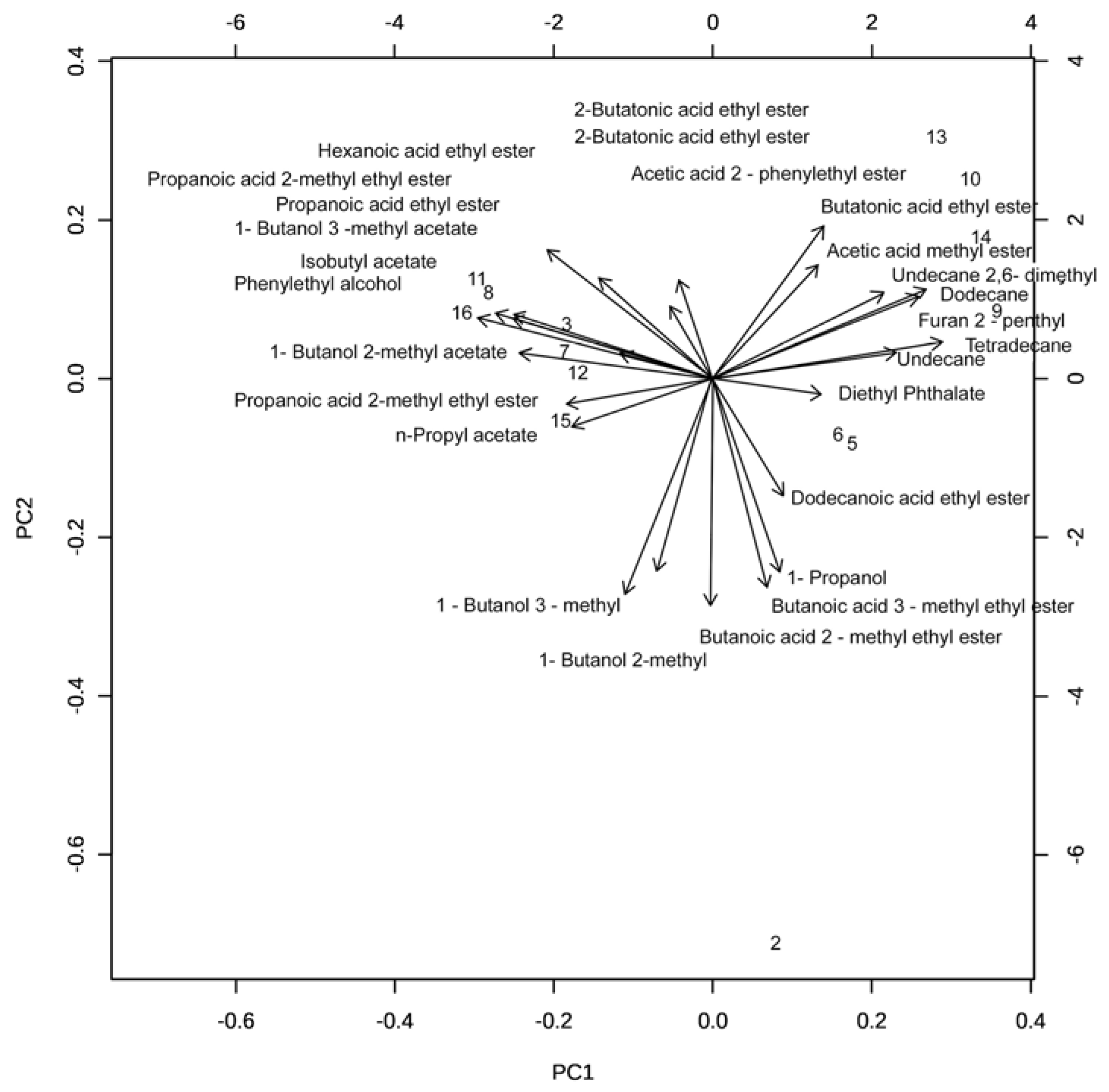
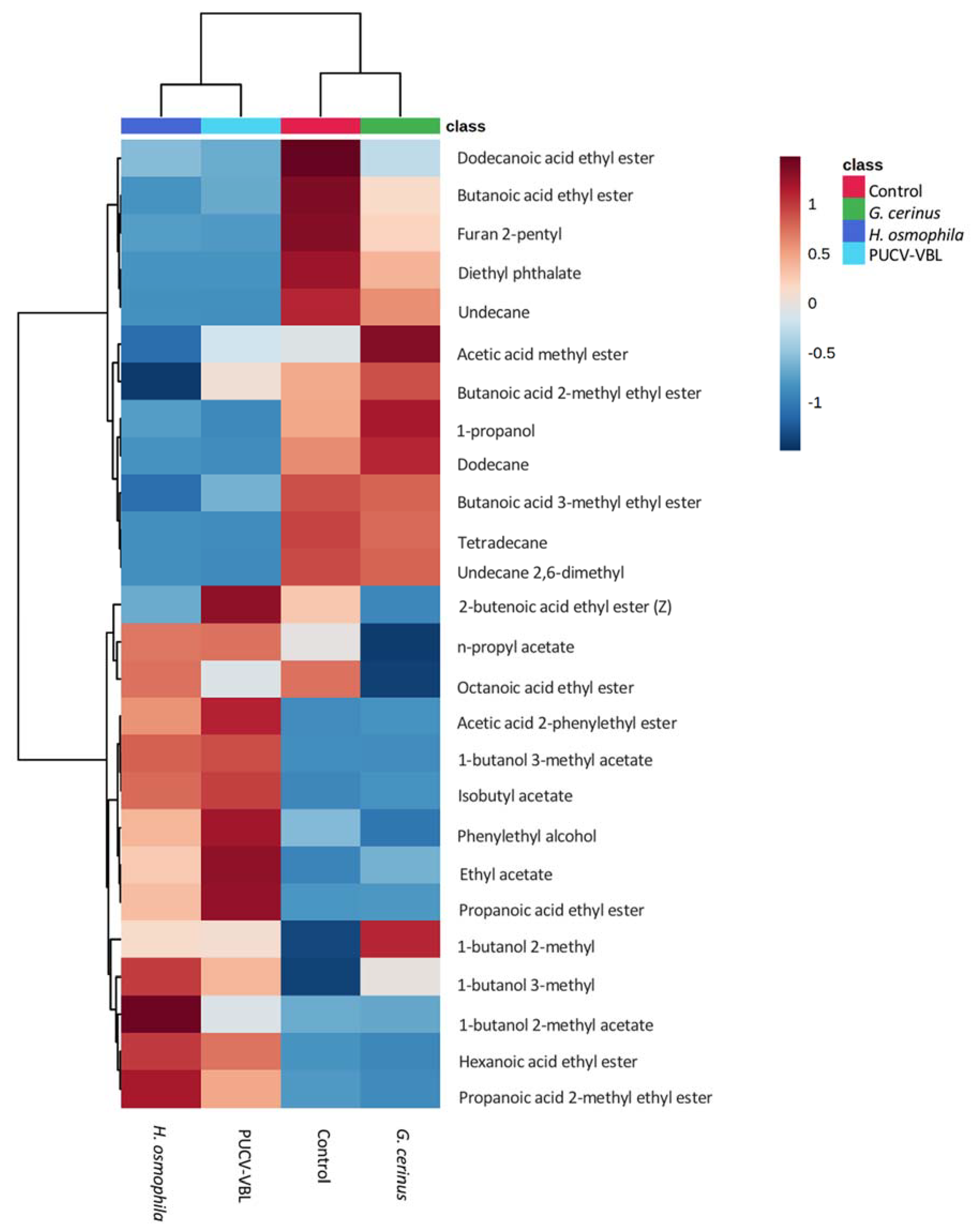
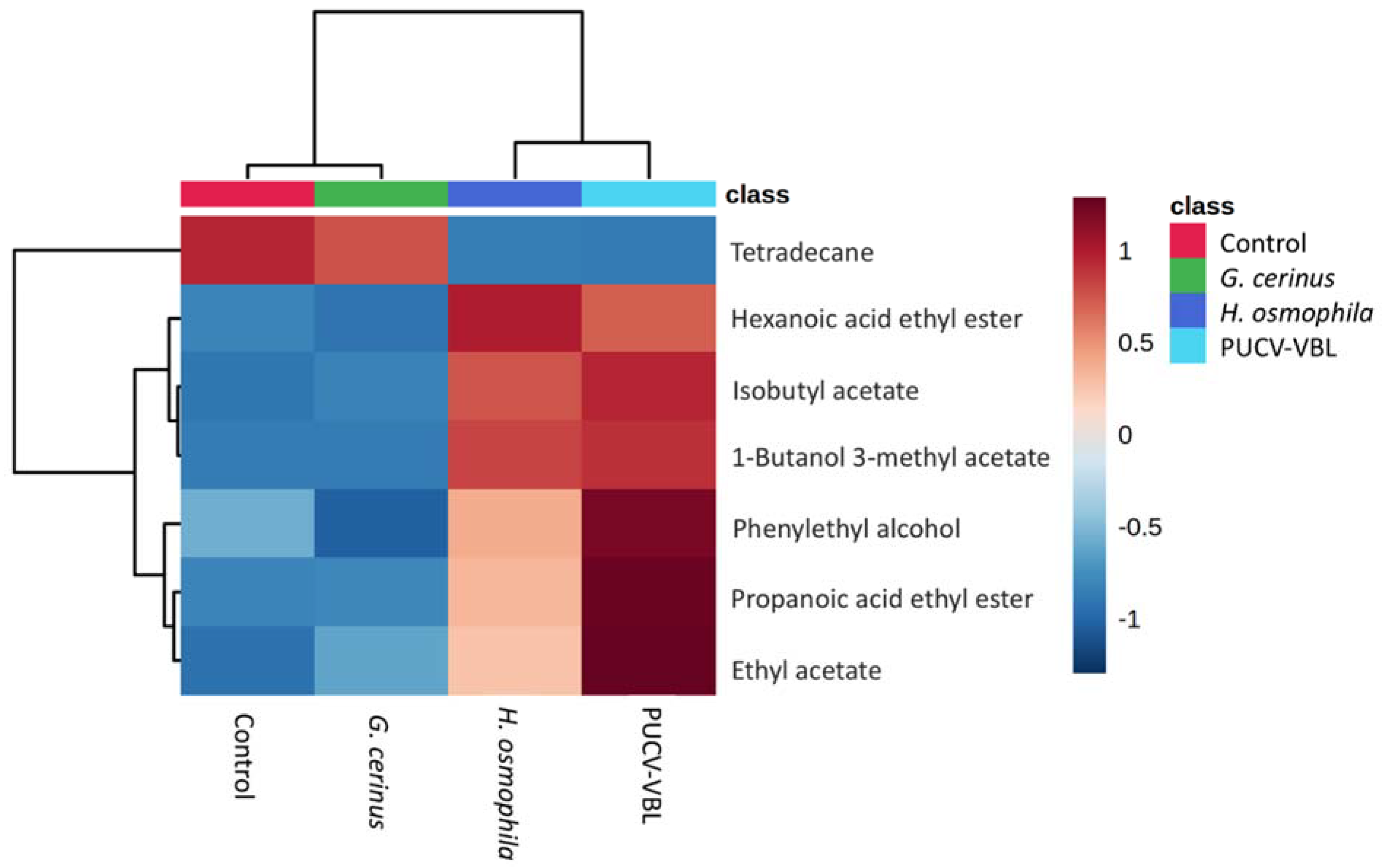
2.3. Identification of VOCs Produced by BCAs (SPME GC-MS)
3. Discussion
4. Materials and Methods
4.1. Microorganism and Vegetable Materials
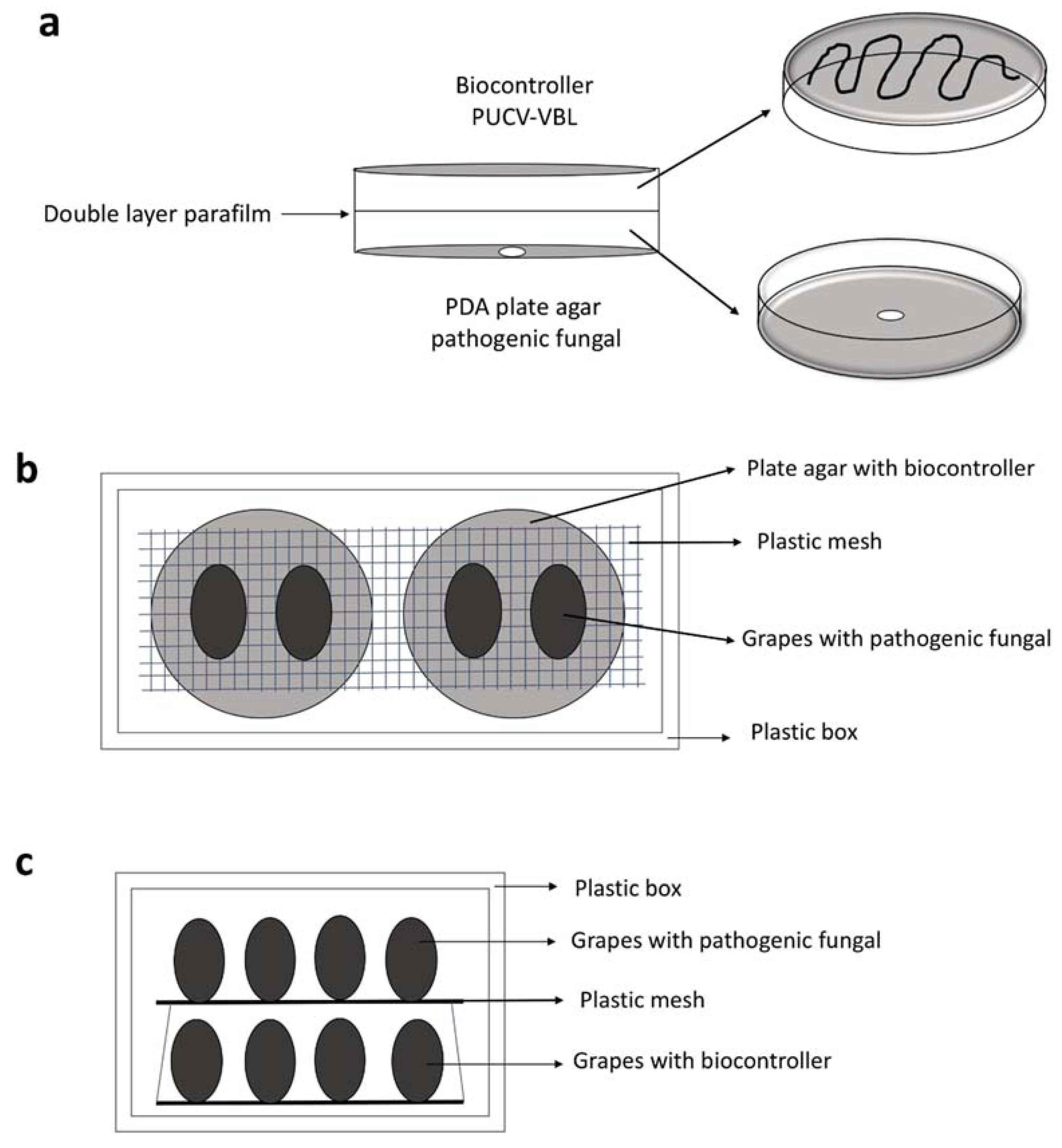
4.2. In Vitro Assays of Biological Consortium VOCs
4.3. In Vivo Assays of Biological Consortium VOCs
4.4. Identification of Biocontroller-Produced VOCs (SPME-GC-MS)
4.5. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stapleton, J.J. Leaf Removal for Nonchemical Control of the Summer Bunch Rot Complex of Wine Grapes in the San Joaquin Valley. Plant Dis. 1992, 76, 205. [Google Scholar] [CrossRef]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Toro, M.E.; Combina, M.; Castellanos de Figueroa, L.I.; Vazquez, F. Biocontrol of fungi isolated from sour rot infected table grapes by Saccharomyces and other yeast species. Postharvest Biol. Technol. 2013, 86, 456–462. [Google Scholar] [CrossRef]
- Esterio, M.; Muñoz, G.; Ramos, C.; Cofré, G.; Estévez, R.; Salinas, A.; Auger, J. Characterization of Botrytis cinerea isolates present in Thompson seedless table grapes in the Central Valley of Chile. Plant Dis. 2011, 95, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Besoain, X.A.; Araya, C.; Salgado, E.; Rendich, A.; Latorre, B.; Piontelli, E. Ethiology of Bunch Rot Complex of Red Globe Table Grapes Determined by Challenge Inoculations. XV Congreso Nacional de Fitopatología, Chile 2005. Available online: http://www.sochifit.cl/site/congreso/html/XV.html#Articulo_38 (accessed on 24 March 2021).
- Mari, M.; Martini, C.; Guidarelli, M.; Neri, F. Postharvest biocontrol of Monilinia laxa, Monilinia fructicola and Monilinia fructigena on stone fruit by two Aureobasidium pullulans strains. Biol. Control 2012, 60, 132–140. [Google Scholar] [CrossRef]
- Errampalli, D. Penicillium expansum (Blue Mold); Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124115682. [Google Scholar]
- Vico, I.; Duduk, N.; Vasic, M.; Nikolic, M. Identification of Penicillium expansum causing postharvest blue mold decay of apple fruit. Pestic. i fitomedicina 2014, 29, 257–266. [Google Scholar] [CrossRef]
- Blum, L.E.B.; Amarante, C.V.T.; Valdebenito-Sanhueza, R.M.; Guimarães, L.S.; Dezanet, A.; Hack Neto, P. Cryptococcus laurentii aplicado em pós-colheita reduz podridões em maçãs. Fitopatol. Bras. 2004, 29, 433–436. [Google Scholar] [CrossRef][Green Version]
- ASOEX Temporada 2019–2020: Exportaciones de uvas de mesa Chilena a China Crecen Potenciadas por Nuevas Variedades. Available online: https://www.asoex.cl/component/content/article/25-noticias/750-temporada-2019-2020-exportaciones-de-uvas-de-mesa-chilena-a-china-crecen-potenciadas-por-nuevas-variedades (accessed on 24 March 2021).
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Esterio, M.; Araneda, M.J.; Román, A.; Pizarro, L.; Copier, C.; Auger, J. First Report of Boscalid Resistant Botrytis cinerea Isolates Carrying the Mutations H272R, H272Y, P225L, and P225H from Table Grape in Chile. Plant Dis. 2015, 99, 891. [Google Scholar] [CrossRef]
- Esterio, M.; Ramos, C.; Walker, A.S.; Fillinger, S.; Leroux, P.; Auger, J. Phenotypic and genetic characterization of Chilean isolates of botrytis cinerea with different levels of sensitivity to fenhexamid. Phytopathol. Mediterr. 2011, 50, 414–420. [Google Scholar] [CrossRef]
- Latorre, B.A.; Flores, V.; Sara, A.M.; Roco, A. Dicarboximide-resistant isolates of Botrytis cinerea from table grape in Chile: Survey and characterization. Plant Dis. 1994, 78, 990–994. [Google Scholar] [CrossRef]
- Leroch, M.; Kretschmer, M.; Hahn, M. Fungicide resistance phenotypes of botrytis cinerea isolates from commercial vineyards in South West Germany. J. Phytopathol. 2011, 159, 63–65. [Google Scholar] [CrossRef]
- Esterio, M.; Auger, J.; Ramos, C.; García, H. First Report of Fenhexamid Resistant Isolates of Botrytis cinerea on Grapevine in Chile. Plant Dis. 2007, 91, 768. [Google Scholar] [CrossRef]
- Robin, D.C.; Marchand, P.A. Evolution of the biocontrol active substances in the framework of the European Pesticide Regulation (EC) No. 1107/2009. Pest Manag. Sci. 2019, 75, 950–958. [Google Scholar] [CrossRef]
- Kevan, P.G.; Shipp, L. Biological Control as Biotechnological Amelioration and Ecosystem Intensification in Managed Ecosystems; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; ISBN 9780128096338. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.A.; Deng, X. Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 2014, 14, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Sundh, I.; Melin, P. Safety and regulation of yeasts used for biocontrol or biopreservation in the food or feed chain. Antonie van Leeuwenhoek 2011, 99, 113–119. [Google Scholar] [CrossRef]
- Salih, E.Y.A.; Fyhrquist, P.; Abdalla, A.M.A.; Abdelgadir, A.Y.; Kanninen, M.; Sipi, M.; Luukkanen, O.; Fahmi, M.K.M.; Elamin, M.H.; Ali, H.A. LC-MS/MS tandem mass spectrometry for analysis of phenolic compounds and pentacyclic triterpenes in antifungal extracts of Terminalia brownii (Fresen). Antibiotics 2017, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Trinetta, V.; McDaniel, A.; Batziakas, K.G.; Yucel, U.; Nwadike, L.; Pliakoni, E. Antifungal packaging film to maintain quality and control postharvest diseases in strawberries. Antibiotics 2020, 9, 618. [Google Scholar] [CrossRef]
- Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F.P.; Valenzuela, M.; Ramírez, I.; Vasconez, I.N.; Álvarez, I.; Jorquera, M.A.; Seeger, M. Microbial diversity of psychrotolerant bacteria isolated from wild flora of andes mountains and patagonia of chile towards the selection of plant growth-promoting bacterial consortia to alleviate cold stress in plants. Microorganisms 2021, 9, 538. [Google Scholar] [CrossRef]
- Kaddes, A.; Fauconnier, M.L.; Sassi, K.; Nasraoui, B.; Jijakli, M.H. Endophytic fungal volatile compounds as solution for sustainable agriculture. Molecules 2019, 24, 1065. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Delory, B.M.; Delaplace, P.; du Jardin, P.; Fauconnier, M.L. Barley (Hordeum distichon L.) roots synthesise volatile aldehydes with a strong age-dependent pattern and release (E)-non-2-enal and (E,Z)-nona-2,6-dienal after mechanical injury. Plant Physiol. Biochem. 2016, 104, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Rouissi, W.; Ugolini, L.; Martini, C.; Lazzeri, L.; Mari, M. Control of postharvest fungal pathogens by antifungal compounds from penicillium expansum. J. Food Prot. 2013, 76, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Xiao, H.; Cheng, X.; Zhou, H.; Si, L. Hanseniaspora uvarum prolongs shelf life of strawberry via volatile production. Food Microbiol. 2017, 63, 205–212. [Google Scholar] [CrossRef]
- Mercier, J.; Smilanick, J.L. Control of green mold and sour rot of stored lemon by biofumigation with Muscodor albus. Biol. Control 2005, 32, 401–407. [Google Scholar] [CrossRef]
- Pagans, E.; Font, X.; Sánchez, A. Emission of volatile organic compounds from composting of different solid wastes: Abatement by biofiltration. J. Hazard. Mater. 2006, 131, 179–186. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Cheng, Z.; Ma, L.; Zhao, L.; Li, J. A comparison of electronic nose and gas chromatography–mass spectrometry on discrimination and prediction of ochratoxin A content in Aspergillus carbonarius cultured grape-based medium. Food Chem. 2019, 297, 124850. [Google Scholar] [CrossRef]
- Fischer, G.; Müller, T.; Schwalbe, R.; Ostrowski, R.; Dott, W. Exposure to airborne fungi, MVOC and mycotoxins in biowaste-handling facilities. Int. J. Hyg. Environ. Health 2000, 203, 97–104. [Google Scholar] [CrossRef]
- Werner, S.; Polle, A.; Brinkmann, N. Belowground communication: Impacts of volatile organic compounds (VOCs) from soil fungi on other soil-inhabiting organisms. Appl. Microbiol. Biotechnol. 2016, 100, 8651–8665. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial volatiles: Small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; De Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef]
- Ul Hassan, Z.; Al Thani, R.; Alnaimi, H.; Migheli, Q.; Jaoua, S. Investigation and Application of Bacillus licheniformis Volatile Compounds for the Biological Control of Toxigenic Aspergillus and Penicillium spp. ACS Omega 2019, 4, 17186–17193. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Gianotti, A.; Sacchetti, G.; Ndagijimana, M.; Ciccarone, C.; Stellarini, A.; Corsetti, A.; Paparella, A. Diversity of food-borne Bacillus volatile compounds and influence on fungal growth. J. Appl. Microbiol. 2015, 119, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of postharvest Botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia. Phytopathology 2011, 101, 859–869. [Google Scholar] [CrossRef]
- Fialho, M.B.; de Moraes, M.H.D.; Tremocoldi, A.R.; Pascholati, S.F. Potential of antimicrobial volatile organic compounds to control Sclerotinia sclerotiorum in bean seeds. Pesqui. Agropecu. Bras. 2011, 46, 137–142. [Google Scholar] [CrossRef]
- Singh, S.K.; Strobel, G.A.; Knighton, B.; Geary, B.; Sears, J.; Ezra, D. An Endophytic Phomopsis sp. Possessing Bioactivity and Fuel Potential with its Volatile Organic Compounds. Microb. Ecol. 2011, 61, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Kudalkar, P.; Strobel, G.; Riyaz-Ul-Hassan, S.; Geary, B.; Sears, J. Muscodor sutura, a novel endophytic fungus with volatile antibiotic activities. Mycoscience 2012, 53, 319–325. [Google Scholar] [CrossRef]
- Zheng, M.; Shi, J.; Shi, J.; Wang, Q.; Li, Y. Antimicrobial effects of volatiles produced by two antagonistic Bacillus strains on the anthracnose pathogen in postharvest mangos. Biol. Control 2013, 65, 200–206. [Google Scholar] [CrossRef]
- Wu, Y.; Yuan, J.; Yaoyao, E.; Raza, W.; Shen, Q.; Huang, Q. Effects of volatile organic compounds from Streptomyces albulus NJZJSA2 on growth of two fungal pathogens. J. Basic Microbiol. 2015, 55, 1104–1117. [Google Scholar] [CrossRef]
- Masoud, W.; Poll, L.; Jakobsen, M. Influence of volatile compounds produced by yeasts predominant during processing of Coffea arabica in East Africa on growth and ochratoxin A (OTA) production by Aspergillus ochraceus. Yeast 2005, 22, 1133–1142. [Google Scholar] [CrossRef]
- Jovanović, V.S.; Simonović, S.; Ilić, M.; Marković, M.; Mitić, V.; Djordjević, A.; Nikolić-Mandić, S. Chemical composition, antimicrobial and antioxidant activities of seseli pallasii besser. (syn seseli varium trev.) essential oils. Rec. Nat. Prod. 2016, 10, 277–286. [Google Scholar]
- Mayser, P. Medium chain fatty acid ethyl esters—Activation of antimicrobial effects by Malassezia enzymes. Mycoses 2015, 58, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Girija, S.; Duraipandiyan, V.; Kuppusamy, P.S.; Gajendran, H.; Rajagopal, R. Chromatographic Characterization and GC-MS Evaluation of the Bioactive Constituents with Antimicrobial Potential from the Pigmented Ink of Loligo duvauceli. Int. Sch. Res. Not. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Premjanu, N.; Jaynthy, C. Antimicrobial activity of diethyl phthalate: An insilico approach. Asian J. Pharm. Clin. Res. 2014, 7, 141–142. [Google Scholar]
- Naik, B.S. Volatile hydrocarbons from endophytic fungi and their efficacy in fuel production and disease control. Egypt. J. Biol. Pest Control 2018, 28, 1–9. [Google Scholar] [CrossRef]
- Mo, E.K.; Sung, C.K. Phenylethyl alcohol (PEA) application slows fungal growth and maintains aroma in strawberry. Postharvest Biol. Technol. 2007, 45, 234–239. [Google Scholar] [CrossRef]
- Yala, S.M.; Schmidtke, L.M.; Gambetta, J.M.; Steel, C.C. Aureobasidium pullulans volatilome identified by a novel, quantitative approach employing SPME-GC-MS, suppressed Botrytis cinerea and Alternaria alternata in vitro. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Kwasiborski, A.; Bajji, M.; Renaut, J.; Delaplace, P.; Haissam Jijakli, M. Identification of metabolic pathways expressed by pichia anomalakh6 in the presence of the pathogen botrytis cinereaon apple: New possible targets for biocontrol improvement. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Fredlund, E.; Blank, L.M.; Schnürer, J.; Sauer, U.; Passoth, V. Oxygen- and glucose-dependent regulation of central carbon metabolism in Pichia anomala. Appl. Environ. Microbiol. 2004, 70, 5905–5911. [Google Scholar] [CrossRef]
- Chang, P.-K.; Hua, S.S.T.; Sarreal, S.B.L.; Li, R.W. Suppression of Aflatoxin Biosynthesis in Aspergillus flavus by 2-Phenylethanol Is Associated with Stimulated Growth and Decreased Degradation of Branched-Chain Amino Acids. Toxins 2015, 7, 3887–3902. [Google Scholar] [CrossRef]
- Tilocca, B.; Cao, A.; Migheli, Q. Scent of a Killer: Microbial Volatilome and Its Role in the Biological Control of Plant Pathogens. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Di Francesco, A.; Zajc, J.; Aprea, N.G.C.E.; Gasperi, F.; Baraldi, N.P.F.C.E. Bioactivity of volatile organic compounds by Aureobasidium species against gray mold of tomato and table grape. World J. Microbiol. Biotechnol. 2020, 36, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mercier, J.; Jiménez, J.I. Control of fungal decay of apples and peaches by the biofumigant fungus Muscodor albus. Postharvest Biol. Technol. 2004, 31, 1–8. [Google Scholar] [CrossRef]
- Stoppacher, N.; Kluger, B.; Zeilinger, S.; Krska, R.; Schuhmacher, R. Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol. Methods 2010, 81, 187–193. [Google Scholar] [CrossRef]
- Miyazawa, M.; Kimura, M.; Yabe, Y.; Tsukamoto, D.; Sakamoto, M.; Horibe, I.; Okuno, Y. Use of solid phase microextraction (SPME) for profiling the volatile metabolites produced by Glomerella cingulata. J. Oleo Sci. 2008, 57, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Fialho, M.B.; Toffano, L.; Pedroso, M.P.; Augusto, F.; Pascholati, S.F. Volatile organic compounds produced by Saccharomyces cerevisiae inhibit the in vitro development of Guignardia citricarpa, the causal agent of citrus black spot. World J. Microbiol. Biotechnol. 2010, 26, 925–932. [Google Scholar] [CrossRef]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef]
- Spadaro, D.; Gullino, M.L. State of the art and future prospects of the biological control of postharvest fruit diseases. Int. J. Food Microbiol. 2004, 91, 185–194. [Google Scholar] [CrossRef]
- Wang, L.; Dou, G.; Guo, H.; Zhang, Q.; Qin, X.; Yu, W.; Jiang, C.; Xiao, H. Volatile organic compounds of Hanseniaspora uvarum increase strawberry fruit flavor and defense during cold storage. Food Sci. Nutr. 2019, 7, 2625–2635. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniaspora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 160–167. [Google Scholar] [CrossRef]
- Cai, Z.; Yang, R.; Xiao, H.; Qin, X.; Si, L. Effect of preharvest application of Hanseniaspora uvarum on postharvest diseases in strawberries. Postharvest Biol. Technol. 2015, 100, 52–58. [Google Scholar] [CrossRef]
- Basha, H.; Ramanujam, B. Growth promotion effect of Pichia guilliermondii in chilli and biocontrol potential of Hanseniaspora uvarum against Colletotrichum capsici causing fruit rot. Biocontrol Sci. Technol. 2015, 25, 185–206. [Google Scholar] [CrossRef]
- Li, W.; Qin, Z.; Zhang, H. Extremal hexagonal chains with respect to the coefficients sum of the permanental polynomial. Appl. Math. Comput. 2016, 291, 30–38. [Google Scholar] [CrossRef]
- Liu, H.M.; Guo, J.H.; Luo, L.; Liu, P.; Wang, B.Q.; Cheng, Y.J.; Deng, B.X.; Long, C.A. Improvement of Hanseniaspora uvarum biocontrol activity against gray mold by the addition of ammonium molybdate and the possible mechanisms involved. Crop Prot. 2010, 29, 277–282. [Google Scholar] [CrossRef]
- Romanazzi, G.; Lichter, A.; Gabler, F.M.; Smilanick, J.L. Recent advances on the use of natural and safe alternatives to conventional methods to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2012, 63, 141–147. [Google Scholar] [CrossRef]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Control 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Li, Q.; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Hsiang, T. Effects of volatile substances of Streptomyces globisporus JK-1 on control of Botrytis cinerea on tomato fruit. Biol. Control 2012, 61, 113–120. [Google Scholar] [CrossRef]
- Li, B.; Lai, T.; Qin, G.; Tian, S. Ambient pH stress inhibits spore germination of Penicillium expansum by impairing protein synthesis and folding: A proteomic-based study. J. Proteome Res. 2010, 9, 298–307. [Google Scholar] [CrossRef]
- Chen, H.; Xiao, X.; Wang, J.; Wu, L.; Zheng, Z.; Yu, Z. Antagonistic effects of volatiles generated by Bacillus subtilis on spore germination and hyphal growth of the plant pathogen, Botrytis cinerea. Biotechnol. Lett. 2008, 30, 919–923. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Performance evaluation of volatile organic compounds by antagonistic yeasts immobilized on hydrogel spheres against gray, green and blue postharvest decays. Food Microbiol. 2016, 63, 191–198. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2017, 265, 18–22. [Google Scholar] [CrossRef]
- Li, Q.; Wu, L.; Hao, J.; Luo, L.; Cao, Y.; Li, J. Biofumigation on post-harvest diseases of fruits using a new volatile-producing fungus of Ceratocystis fimbriata. PLoS ONE 2015, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.P. Recent advances in crop protection. Recent Adv. Crop Prot. 2013, 1–268. [Google Scholar] [CrossRef]
- Olivera, M.; Delgado, N.; Cádiz, F.; Riquelme, N.; Montenegro, I.; Seeger, M.; Bravo, G.; Barros, W.; Pedreschi, R.; Besoain, X. Diffusible compounds produced by Hanseniaspora osmophila and Gluconobacter cerinus help to control the causal agents of gray rot and summer bunch rot of table grapes. 2021, unpublished; manuscript accepted. 2021; unpublished; manuscript accepted. [Google Scholar]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Teuber, M. Genetic and restriction analysis of the 16S-23S rDNA internal transcribed spacer regions of the acetic acid bacteria. FEMS Microbiol. Lett. 2002, 208, 69–75. [Google Scholar] [CrossRef]
- Leaw, S.N.; Chang, H.C.; Sun, H.F.; Barton, R.; Bouchara, J.; Chang, T.C. Identification of medically important yeast species by sequence analysis of the internal transcribed spacer regions. J. Clin. Microbiol. 2006, 44, 693–699. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods an Application Ed.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G. cerinus | H. osmophila | PUCV-VBL | |
|---|---|---|---|
| B. cinerea | 32 ± 15 a | 39 ± 26 a | 86 ± 4 b |
| A. tubingensis | 36 ± 2 a | 53 ± 16 a | 52 ± 1 a |
| P. expansum | 50 ± 13 a | 68 ± 1 a | 55 ± 2 a |
| R. stolonifer | 18 ± 3 a | 29 ± 8 a,b | 35 ± 7 b |
| Cultivar | Pathogen | Biocontrol Agents (BCAs) | ||
|---|---|---|---|---|
| G. cerinus | H. osmophila | PUCV-VBL | ||
| Red Globe | B. cinerea | 82 ± 9 a | 76 ± 14 a | 79 ± 15 a |
| A. tubingensis | 75 ± 22 a,b | 68 ± 23 a | 93 ± 11 b | |
| P. expansum | 71 ± 12 a | 66 ± 14 a | 78 ± 12 a | |
| R. stolonifer | 82 ± 12 a | 73 ± 10 a | 84 ± 15 a | |
| Thompson seedless | B. cinerea | 54 ± 22 a | 75 ± 11 b | 79 ± 8 b |
| A. tubingensis | 77 ± 16 a | 73 ± 7 a | 71 ± 13 a | |
| P. expansum | 77 ± 22 a | 78 ± 9 a | 85 ± 2 a | |
| R. stolonifer | 68 ± 24 a | 71 ± 22 a | 76 ± 17 a | |
| Crimson seedless | B. cinerea | 82 ± 9 b | 54 ± 21 a | 86 ± 12 b |
| A. tubingensis | 64 ± 21 a,b | 57 ± 18 a | 75 ± 14 b | |
| P. expansum | 50 ± 11 a | 49 ± 15 a | 65 ± 13 b | |
| R. stolonifer | 89 ± 13 b | 60 ± 20 a | 64 ± 14 a | |
| Cultivar | Pathogen | G. cerinus | H. osmophila | PUCV-VBL |
|---|---|---|---|---|
| Red Globe | B. cinerea | 85 ± 17 a | 83 ± 15 a | 83 ± 12 a |
| A. tubingensis | 56 ± 10 a | 51 ± 13 a | 38 ± 15 a | |
| P. expansum | 50 ± 17 a | 44 ± 7 a | 46 ± 10 a | |
| R. stolonifer | 44 ± 15 a | 32 ± 20 a | 45 ± 19 a | |
| Thompson seedless | B. cinerea | 96 ± 2 a | 96 ± 2 a | 98 ± 1 a |
| A. tubingensis | 94 ± 7 b | 73 ± 4 a | 95 ± 4 b | |
| P. expansum | 75 ± 14 a | 81± 9 a,b | 93 ± 4 b | |
| R. stolonifer | 84 ± 14 a | 83 ± 15 a | 91 ± 3 a | |
| Crimson seedless | B. cinerea | 51 ± 19 a | 34 ± 15 a | 72 ± 10 b |
| A. tubingensis | 56 ± 16 a | 65 ± 12 a | 53 ± 17 a | |
| P. expansum | 37 ± 14 a | 41 ± 17 a | 57 ± 17 a | |
| R. stolonifer | 54 ± 17 a | 66 ± 4 a | 89 ± 20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, N.; Olivera, M.; Cádiz, F.; Bravo, G.; Montenegro, I.; Madrid, A.; Fuentealba, C.; Pedreschi, R.; Salgado, E.; Besoain, X. Volatile Organic Compounds (VOCs) Produced by Gluconobacter cerinus and Hanseniaspora osmophila Displaying Control Effect against Table Grape-Rot Pathogens. Antibiotics 2021, 10, 663. https://doi.org/10.3390/antibiotics10060663
Delgado N, Olivera M, Cádiz F, Bravo G, Montenegro I, Madrid A, Fuentealba C, Pedreschi R, Salgado E, Besoain X. Volatile Organic Compounds (VOCs) Produced by Gluconobacter cerinus and Hanseniaspora osmophila Displaying Control Effect against Table Grape-Rot Pathogens. Antibiotics. 2021; 10(6):663. https://doi.org/10.3390/antibiotics10060663
Chicago/Turabian StyleDelgado, Ninoska, Matías Olivera, Fabiola Cádiz, Guillermo Bravo, Iván Montenegro, Alejandro Madrid, Claudia Fuentealba, Romina Pedreschi, Eduardo Salgado, and Ximena Besoain. 2021. "Volatile Organic Compounds (VOCs) Produced by Gluconobacter cerinus and Hanseniaspora osmophila Displaying Control Effect against Table Grape-Rot Pathogens" Antibiotics 10, no. 6: 663. https://doi.org/10.3390/antibiotics10060663
APA StyleDelgado, N., Olivera, M., Cádiz, F., Bravo, G., Montenegro, I., Madrid, A., Fuentealba, C., Pedreschi, R., Salgado, E., & Besoain, X. (2021). Volatile Organic Compounds (VOCs) Produced by Gluconobacter cerinus and Hanseniaspora osmophila Displaying Control Effect against Table Grape-Rot Pathogens. Antibiotics, 10(6), 663. https://doi.org/10.3390/antibiotics10060663