
Allium Extract Implements Weaned Piglet’s Productive Parameters by Modulating Distal Gut Microbiota

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Piglets and Farm Facilities
2.2. Experimental Design and Sample Collection
2.3. DNA Extraction
2.4. High-Throughput Sequencing
2.5. Sequences Processing and Data Analysis
2.6. Statistics
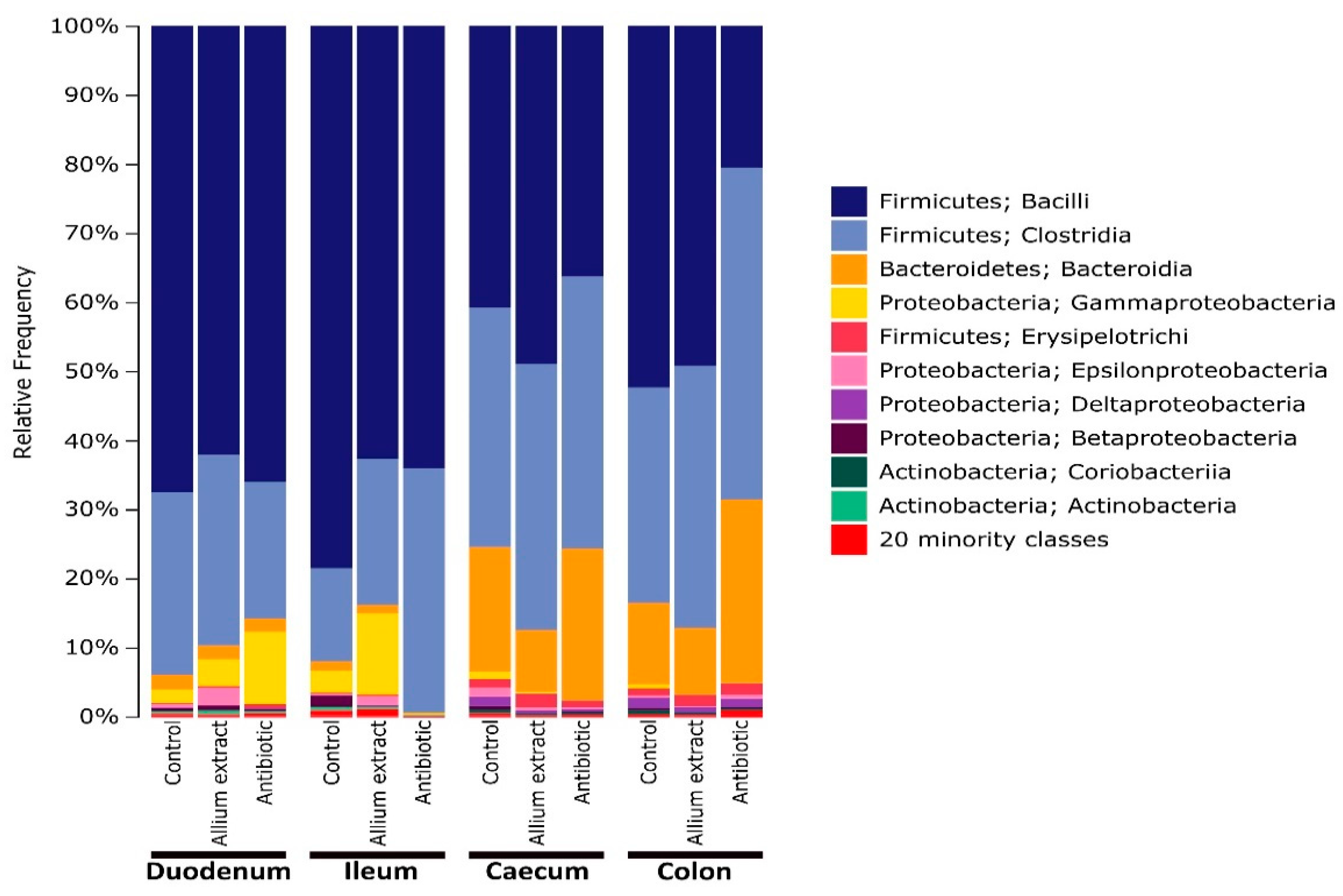
3. Results
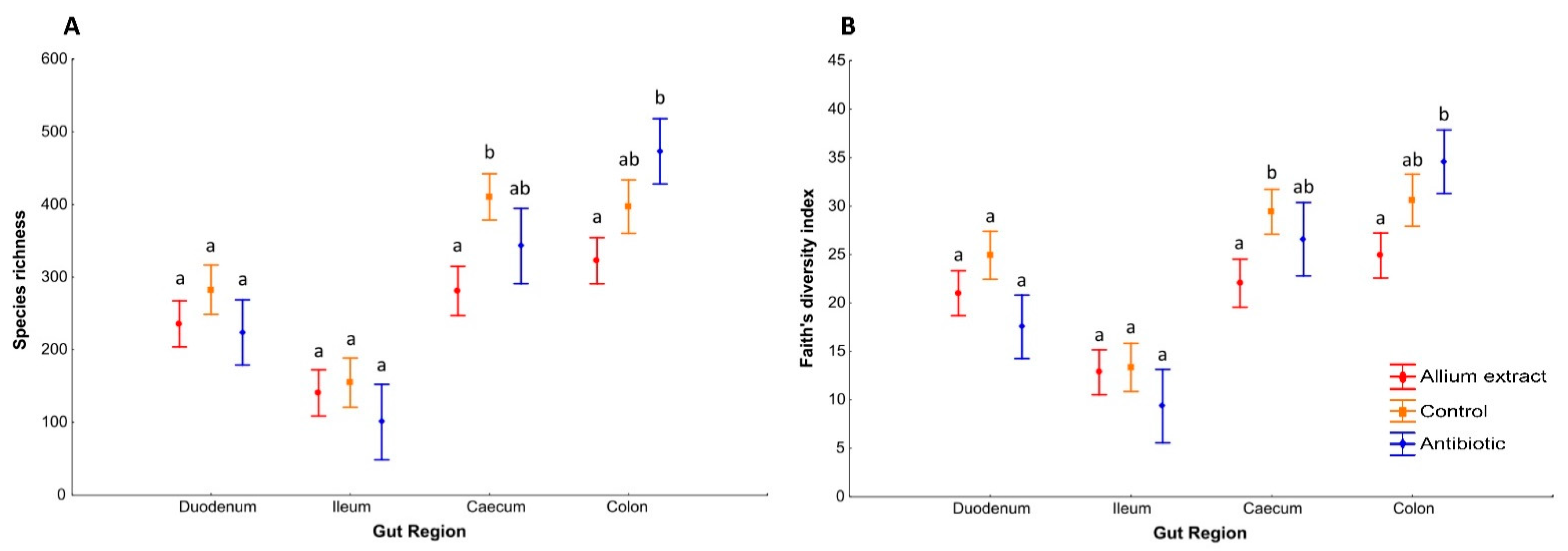
3.1. Effects of Teatment on Piglets’ Gut Bacterial Alpha Diversity
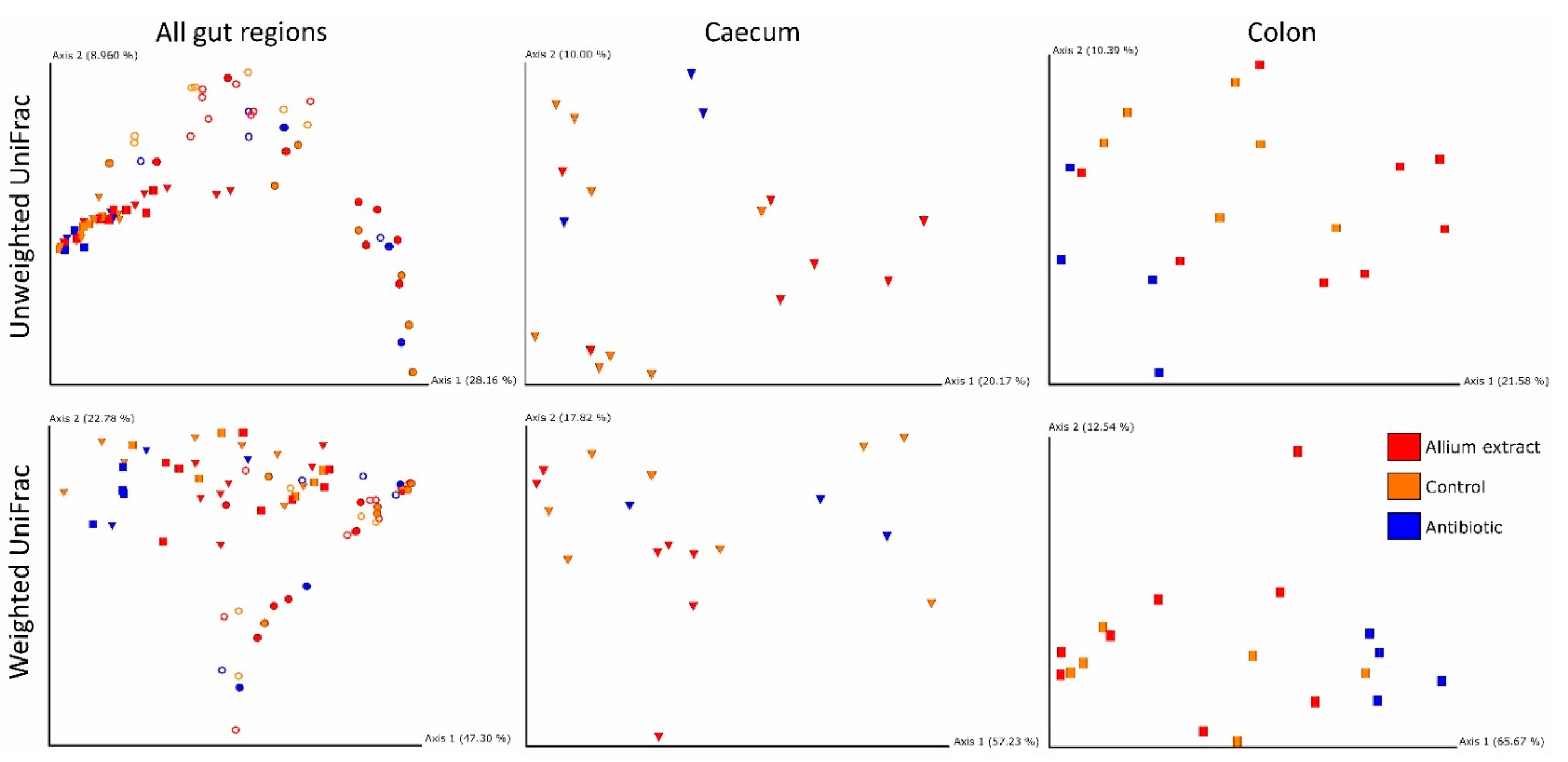
3.2. Effects of Treatment and Gut Region on Beta Diversity
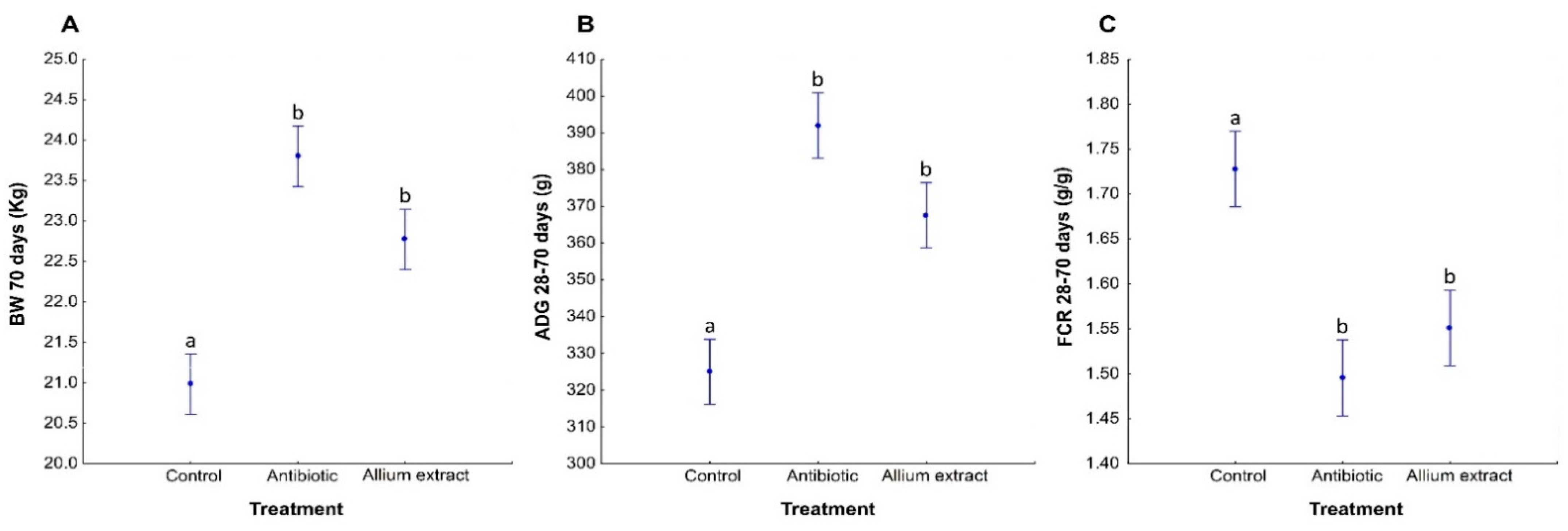
3.3. Effects of Treatment on Piglets’ Productive Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, I.; Casewell, M.; Cox, T.; De Groot, B.; Friis, C.; Jones, R.; Nightingale, C.; Preston, R.; Waddell, J. Does the use of antibiotics in food animals pose a risk to human health? A critical review of published data. J. Antimicrob. Chemother. 2003, 53, 28–52. [Google Scholar] [CrossRef]
- A Thacker, P. Alternatives to antibiotics as growth promoters for use in swine production: A review. J. Anim. Sci. Biotechnol. 2013, 4, 35. [Google Scholar] [CrossRef]
- Capita, R.; Alonso-Calleja, C. Antibiotic-resistant bacteria: A challenge for the food industry. Crit. Rev. Food Sci. Nutr. 2013, 53, 11–48. [Google Scholar] [CrossRef]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014; pp. 1–257. [Google Scholar]
- U.S. FDA. FDA Reminds Retail Establishments of Upcoming Changes to the Use of Antibiotics in Food Animals; U.S. FDA: Silver Spring, MA, USA, 2016. [Google Scholar]
- Maron, D.F.; Smith, T.J.S.; E Nachman, K. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Glob. Health 2013, 9, 48. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Dietze, K. Pigs for Prosperity (FAO Diversification Booklet 15); U.S. FAO: Hot Springs, VA, USA, 2011; ISBN 978-92-5-107068-0. [Google Scholar]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Microbial shifts in the swine distal gut in response to the treatment with antimicrobial growth promoter, tylosin. Proc. Natl. Acad. Sci. USA 2012, 109, 15485–15490. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef]
- Gaskins, H.R.; Collier, C.T.; Anderson, D.B. Antibiotics as growth promotants: Mode of action. Anim. Biotechnol. 2002, 13, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Niewold, T.A. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult. Sci. 2007, 86, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D. Impact of antibiotic use in the swine industry. Curr. Opin. Microbiol. 2014, 19, 9–15. [Google Scholar] [CrossRef]
- Faldynova, M.; Videnska, P.; Havlickova, H.; Sisak, F.; Juricova, H.; Babak, V.; Steinhauser, L.; Rychlik, I. Prevalence of antibiotic resistance genes in faecal samples from cattle, pigs and poultry. Vet. Med. 2013, 58, 298–304. [Google Scholar] [CrossRef]
- Gerzova, L.; Babak, V.; Sedlar, K.; Faldynova, M.; Videnska, P.; Cejkova, D.; Jensen, A.N.; Denis, M.; Kerouanton, A.; Ricci, A.; et al. Characterization of antibiotic resistance gene abundance and microbiota composition in feces of organic and conventional pigs from four EU countries. PLoS ONE 2015, 10, e0132892. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Neves, E.; Antunes, P.; Manageiro, V.; Gartner, F.; Caniça, M.; Da Costa, J.M.C.; Peixe, L. Clinically relevant multidrug resistant Salmonella enterica in swine and meat handlers at the abattoir. Vet. Microbiol. 2014, 168, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Peeters, L.E.; Argudín, M.A.; Azadikhah, S.; Butaye, P.; Regueiro, M. Ángeles A. Antimicrobial resistance and population structure of Staphylococcus aureus recovered from pigs farms. Vet. Microbiol. 2015, 180, 151–156. [Google Scholar] [CrossRef]
- Thakur, S.; Gebreyes, W.A. Prevalence and antimicrobial resistance of Campylobacter in antimicrobial-free and conventional pig production systems. J. Food Prot. 2005, 68, 2402–2410. [Google Scholar] [CrossRef]
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86, E140–E148. [Google Scholar] [CrossRef]
- Li, P.; Piao, X.; Ru, Y.; Han, X.; Xue, L.; Zhang, H. Effects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian Australas. J. Anim. Sci. 2012, 25, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Ragland, D.; Stevenson, D.; Hill, M.A. Oregano oil and multi-component carbohydrases as alternatives to antimicrobials in nursery diets. J. Swine Health Prod. 2008, 16, 238–243. [Google Scholar]
- Maenner, K.; Vahjen, W.; Simon, O. Studies on the effects of essential-oil-based feed additives on performance, ileal nutrient digestibility, and selected bacterial groups in the gastrointestinal tract of piglets. J. Anim. Sci. 2011, 89, 2106–2112. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Hossain, M.E.; Kim, G.M.; Hwang, J.A.; Ji, H.; Yang, C.J. Effects of resveratrol and essential oils on growth performance, immunity, digestibility and fecal microbial shedding in challenged piglets. Asian Australas. J. Anim. Sci. 2013, 26, 683–690. [Google Scholar] [CrossRef]
- Huang, Y.; Yoo, J.S.; Kim, H.J.; Wang, Y.; Chen, Y.J.; Cho, J.H.; Kim, I.H. Effects of dietary supplementation with blended essential oils on growth performance, nutrient digestibility, blood profiles and fecal characteristics in weanling pigs. Asian Australas. J. Anim. Sci. 2010, 23, 607–613. [Google Scholar] [CrossRef]
- Liu, Y.; Song, M.; Che, T.M.; Almeida, J.A.S.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli. J. Anim. Sci. 2013, 91, 5294–5306. [Google Scholar] [CrossRef]
- Harris, J.C.; Cottrell, S.L.; Plummer, S.; Lloyd, D. Antimicrobial properties of Allium sativum (garlic). Appl. Microbiol. Biotechnol. 2001, 57, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Tatara, M.R.; Sliwa, E.; Dudek, K.; Gawron, A.; Piersiak, T.; Dobrowolski, P.; Mosiewicz, J.; Siwicki, A.; Studzinski, T. Aged garlic extract and allicin improve performance and gastrointestinal tract development of piglets reared in artificial sow. Ann. Agric. Environ. Med. 2008, 15, 63–69. [Google Scholar]
- Ruiz, R.; García, M.P.; Lara, A.; Rubio, L.A. Garlic derivatives (PTS and PTS-O) differently affect the ecology of swine faecal microbiota in vitro. Vet. Microbiol. 2010, 144, 110–117. [Google Scholar] [CrossRef]
- Peinado, M.J.; Ruiz, R.; Echávarri, A.; Rubio, L.A. Garlic derivative propyl propane thiosulfonate is effective against broiler enteropathogens in vivo. Poult. Sci. 2012, 91, 2148–2157. [Google Scholar] [CrossRef]
- Sorlozano-Puerto, A.; Albertuz-Crespo, M.; Lopez-Machado, I.; Ariza-Romero, J.J.; Baños-Arjona, A.; Exposito-Ruiz, M.; Gutierrez-Fernandez, J. In vitro antibacterial activity of propyl-propane-thiosulfinate and propyl-propane-thiosulfonate derived from allium spp. against gram-negative and gram-positive multidrug-resistant bacteria isolated from human samples. BioMed Res. Int. 2018, 2018, 7861207. [Google Scholar] [CrossRef] [PubMed]
- Peinado, M.; Ruiz, R.; Echávarri, A.; Aranda-Olmedo, I.; Rubio, L. Garlic derivative PTS-O modulates intestinal microbiota composition and improves digestibility in growing broiler chickens. Anim. Feed. Sci. Technol. 2013, 181, 87–92. [Google Scholar] [CrossRef]
- Hawkins, T. DNA Purification and Isolation Using Magnetic Particles. U.S. Patent 5,705,628, 6 January 1998. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur rapidly resolves single-nucleotide community sequence patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef] [PubMed]
- Janssen, S.; McDonald, D.; Gonzalez, A.; Navas-Molina, J.A.; Jiang, L.; Xu, Z.Z.; Winker, K.; Kado, D.M.; Orwoll, E.; Manary, M.; et al. Phylogenetic placement of exact amplicon sequences improves associations with clinical information. mSystems 2018, 3, e00021-18. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Faith, D.P.; Baker, A.M. Phylogenetic diversity (PD) and biodiversity conservation: Some bioinformatics challenges. Evol. Bioinform. 2006, 2, 121–128. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Collyer, M.L.; Sekora, D.J.; Adams, D.C. A method for analysis of phenotypic change for phenotypes described by high-dimensional data. Hered. 2015, 115, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Soler, C.; Goossens, T.; Bermejo, A.; Migura-García, L.; Cusco, A.; Francino, O.; Fraile, L. Digestive microbiota is different in pigs receiving antimicrobials or a feed additive during the nursery period. PLoS ONE 2018, 13, e0197353. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Xia, M.; Zhang, X.; Wang, C.; Jiang, S.; Peng, J. Supplementing oregano essential oil in a reduced-protein diet improves growth performance and nutrient digestibility by modulating intestinal bacteria, intestinal morphology, and antioxidative capacity of growing-finishing pigs. Animals 2018, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Resano, H.; Perez-Cueto, F.J.; De Barcellos, M.D.; Veflen-Olsen, N.; Grunert, K.G.; Verbeke, W. Consumer satisfaction with pork meat and derived products in five European countries. Appetite 2011, 56, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Hanczakowska, E.; Świątkiewicz, M.; Grela, E.R. Effect of dietary inclusion of a herbal extract mixture and different oils on pig performance and meat quality. Meat Sci. 2015, 108, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Hernández, F.; Madrid, J.; García, V.; Orengo, J.; Megías, M.D. Influence of two plant extracts on broilers performance, digestibility, and digestive organ size. Poult. Sci. 2004, 83, 169–174. [Google Scholar] [CrossRef]
- Hanczakowska, E.; Swiatkiewicz, M. Effect of herbal extracts on piglet performance and small intestinal epithelial villi. Czech J. Anim. Sci. 2012, 57, 420–429. [Google Scholar] [CrossRef]
- Nowak, P.; Kasprowicz-Potocka, M.; Zaworska, A.; Nowak, W.; Stefańska, B.; Sip, A.; Grajek, W.; Juzwa, W.; Taciak, M.; Barszcz, M.; et al. The effect of eubiotic feed additives on the performance of growing pigs and the activity of intestinal microflora. Arch. Anim. Nutr. 2017, 71, 455–469. [Google Scholar] [CrossRef]
- De Rodas, B.; Youmans, B.P.; Danzeisen, J.L.; Tran, H.; Johnson, T.J. Microbiome profiling of commercial pigs from farrow to finish. J. Anim. Sci. 2018, 96, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Kong, F.; Xiang, Y.; Zhou, W.; Wang, J.; Yang, H.; Zhang, G.; Zhao, J. Comparative biogeography of the gut microbiome between Jinhua and Landrace pigs. Sci. Rep. 2018, 8, 5985. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, X.; Fang, S.; Xin, W.; Huang, L.; Chen, C. Uncovering the composition of microbial community structure and metagenomics among three gut locations in pigs with distinct fatness. Sci. Rep. 2016, 6, 27427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, W.; Lee, Y.-K.; Xie, J.; Zhang, H. Spatial Heterogeneity and Co-occurrence of Mucosal and Luminal Microbiome across Swine Intestinal Tract. Front. Microbiol. 2018, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Kim, J.; Lee, J.-S.; Rhee, S.-K.; Kim, H. Characterization of the fecal microbiome in different swine groups by high-throughput sequencing. Anaerobe 2014, 28, 157–162. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, L.; Wu, Y.-P.; Cui, Z.-W.; Wang, Y.-Q.; Huang, Y.; Zhang, X.-P.; Li, W.-F. Oral administration of Lactobacillus rhamnosus GG to newborn piglets augments gut barrier function in pre-weaning piglets. J. Zhejiang Univ. Sci. B 2019, 20, 180–192. [Google Scholar] [CrossRef]
- Warren, F.J.; Fukuma, N.M.; Mikkelsen, D.; Flanagan, B.M.; Williams, B.A.; Lisle, A.T.; Cuív, P.Ó.; Morrison, M.; Gidley, M.J. Food starch structure impacts gut microbiome composition. mSphere 2018, 3, e00086-18. [Google Scholar] [CrossRef]
- Rahman, S.U.; Khan, S.; Chand, N.; Sadique, U.; Khan, R.U. In vivo effects of Allium cepa L. on the selected gut microflora and intestinal histomorphology in broiler. Acta Histochem. 2017, 119, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.J.; Martínez-Miró, S.; Ariza, J.J.; Madrid, J.; Orengo, J.; Aguinaga, M.A.; Baños, A.; Hernández, F. Effect of Alliaceae extract supplementation on performance and intestinal microbiota of growing-finishing pig. Animals 2020, 10, 1557. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Allen, H.K.; Cantarel, B.L.; Levine, U.Y.; O Bayles, D.; Alt, D.P.; Henrissat, B.; Stanton, T.B. Bacteria, phages and pigs: The effects of in-feed antibiotics on the microbiome at different gut locations. ISME J. 2014, 8, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Arroyo-Manzanares, N.; Rivas-Montoya, E.; Ochando-Pulido, J.M.; Guillamon, E.; Garcia-Campaña, A.M.; Martinez-Ferez, A. Effects of different vehiculization strategies for the allium derivative propyl propane thiosulfonate during dynamic simulation of the pig gastrointestinal tract. Can. J. Anim. Sci. 2019, 99, 244–253. [Google Scholar] [CrossRef]
- Menni, C.; A Jackson, M.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. 2017, 41, 1099–1105. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2008, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Chae, B.J.; Kim, E.B.; Choi, Y.-J. Evaluating the association between body weight and the intestinal microbiota of weaned piglets via 16S rRNA sequencing. Appl. Microbiol. Biotechnol. 2017, 101, 5903–5911. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E. Weight gain by gut microbiota manipulation in productive animals. Microb. Pathog. 2017, 106, 162–170. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha Diversity Index | Control | Allium Extract | Antibiotic | Explanatory Variables | D.f. | F | p |
|---|---|---|---|---|---|---|---|
| Species richness | 311.75 (27.38) | 243.77 (18.71) | 294.14 (41.60) | Treatment | 2.61 | 4.03 | 0.023 |
| Gut Region | 3.61 | 26.41 | <0.001 | ||||
| Gut Region × Treatment | 6.61 | 1.57 | 0.171 | ||||
| Faith’s diversity index | 24.53 (1.89) | 20.14 (1.31) | 22.59 (2.89) | Treatment | 2.61 | 3.25 | 0.046 |
| Gut Region | 3.61 | 22.11 | <0.001 | ||||
| Gut Region × Treatment | 6.61 | 1.46 | 0.208 |
| β-Diversity Distance Matrix | Explanatory Variables | D.f. | Pseudo-F | p |
|---|---|---|---|---|
| Unweighted UniFrac | Treatment | 2.61 | 1.84 | 0.001 |
| Gut Region | 3.61 | 7.88 | 0.001 | |
| Gut Region × Treatment | 6.61 | 1.06 | 0.303 | |
| Weighted UniFrac | Treatment | 2.61 | 2.35 | 0.044 |
| Gut Region | 3.61 | 9.14 | 0.001 | |
| Gut Region × Treatment | 6.61 | 1.02 | 0.412 |
| β-Diversity Distance Matrix | D.f. | Pseudo-F | p | |
|---|---|---|---|---|
| Duodenum | Unweighted UniFrac | 2.16 | 1.23 | 0.099 |
| Weighted UniFrac | 2.16 | 0.25 | 0.977 | |
| Ileum | Unweighted UniFrac | 2.15 | 0.93 | 0.502 |
| Weighted UniFrac | 2.15 | 1.08 | 0.377 | |
| Caecum | Unweighted UniFrac | 2.15 | 1.56 | 0.007 |
| Weighted UniFrac | 2.15 | 1.48 | 0.191 | |
| Colon | Unweighted UniFrac | 2.15 | 1.55 | 0.017 |
| Weighted UniFrac | 2.15 | 4.18 | 0.009 |
| Dependent Variable | Control | Allium Extract | Antibiotic | Independent Variables | F | D.f. | p |
|---|---|---|---|---|---|---|---|
| Initial BW (28 days), kg | 7.34 (0.35) | 7.34 (0.33) | 7.32 (0.37) | Treatment | <0.01 | 2.19 | 0.998 |
| Sex | 0.21 | 1.19 | 0.653 | ||||
| Room | 0.93 | 1.19 | 0.346 | ||||
| BW 42 days, kg | 10.50 (0.49) | 10.87 (0.57) | 11.40 (0.55) | Treatment | 5.69 | 2.18 | 0.012 |
| Sex | 4.98 | 1.18 | 0.039 | ||||
| Room | 29.67 | 1.18 | <0.001 | ||||
| Initial BW | 116.14 | 1.18 | <0.001 | ||||
| BW 70 days, kg | 21.01 (0.76) | 22.79 (0.98) | 23.76 (0.92) | Treatment | 14.59 | 2.18 | <0.001 |
| Sex | 8.96 | 1.18 | 0.008 | ||||
| Room | 2.46 | 1.18 | 0.134 | ||||
| Initial BW | 86.30 | 1.18 | <0.001 | ||||
| ADG 28–70 days, g/d | 325.25 (11.01) | 367.71 (16.84) | 391.53 (15.37) | Treatment | 14.59 | 2.18 | <0.001 |
| Sex | 8.96 | 1.18 | 0.008 | ||||
| Room | 2.46 | 1.18 | 0.134 | ||||
| Initial BW | 26.13 | 1.18 | <0.001 | ||||
| ADFI 28–70 days, g/d | 562.73 (30.24) | 566.26 (23.25) | 583.79 (19.97) | Treatment | 0.49 | 2.18 | 0.620 |
| Sex | 2.14 | 1.18 | 0.161 | ||||
| Room | 18.45 | 1.18 | <0.001 | ||||
| Initial BW | 11.22 | 1.18 | 0.004 | ||||
| FCR 28–70 days, g/g | 1.73 (0.06) | 1.55 (0.06) | 1.50 (0.04) | Treatment | 8.27 | 2.18 | 0.003 |
| Sex | 1.64 | 1.18 | 0.216 | ||||
| Room | 11.88 | 1.18 | 0.003 | ||||
| Initial BW | 1.26 | 1.18 | 0.277 | ||||
| Mortality 28–70 days, % | 5.00 (2.67) | 2.50 (1.64) | 1.25 (1.25) | Treatment | 0.90 | 2.18 | 0.423 |
| Sex | 0.08 | 1.18 | 0.787 | ||||
| Room | 1.52 | 1.18 | 0.233 | ||||
| Initial BW | 0.66 | 1.18 | 0.428 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabelo-Ruiz, M.; Teso-Pérez, C.; Peralta-Sánchez, J.M.; Ariza, J.J.; Martín-Platero, A.M.; Casabuena-Rincón, Ó.; Vázquez-Chas, P.; Guillamón, E.; Aguinaga-Casañas, M.A.; Maqueda, M.; et al. Allium Extract Implements Weaned Piglet’s Productive Parameters by Modulating Distal Gut Microbiota. Antibiotics 2021, 10, 269. https://doi.org/10.3390/antibiotics10030269
Rabelo-Ruiz M, Teso-Pérez C, Peralta-Sánchez JM, Ariza JJ, Martín-Platero AM, Casabuena-Rincón Ó, Vázquez-Chas P, Guillamón E, Aguinaga-Casañas MA, Maqueda M, et al. Allium Extract Implements Weaned Piglet’s Productive Parameters by Modulating Distal Gut Microbiota. Antibiotics. 2021; 10(3):269. https://doi.org/10.3390/antibiotics10030269
Chicago/Turabian StyleRabelo-Ruiz, Miguel, Claudia Teso-Pérez, Juan Manuel Peralta-Sánchez, Juan José Ariza, Antonio Manuel Martín-Platero, Óscar Casabuena-Rincón, Patricia Vázquez-Chas, Enrique Guillamón, María Arántzazu Aguinaga-Casañas, Mercedes Maqueda, and et al. 2021. "Allium Extract Implements Weaned Piglet’s Productive Parameters by Modulating Distal Gut Microbiota" Antibiotics 10, no. 3: 269. https://doi.org/10.3390/antibiotics10030269
APA StyleRabelo-Ruiz, M., Teso-Pérez, C., Peralta-Sánchez, J. M., Ariza, J. J., Martín-Platero, A. M., Casabuena-Rincón, Ó., Vázquez-Chas, P., Guillamón, E., Aguinaga-Casañas, M. A., Maqueda, M., Valdivia, E., Baños, A., & Martínez-Bueno, M. (2021). Allium Extract Implements Weaned Piglet’s Productive Parameters by Modulating Distal Gut Microbiota. Antibiotics, 10(3), 269. https://doi.org/10.3390/antibiotics10030269