
Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter jejuni

 ,
,
Abstract
1. Introduction
2. Results
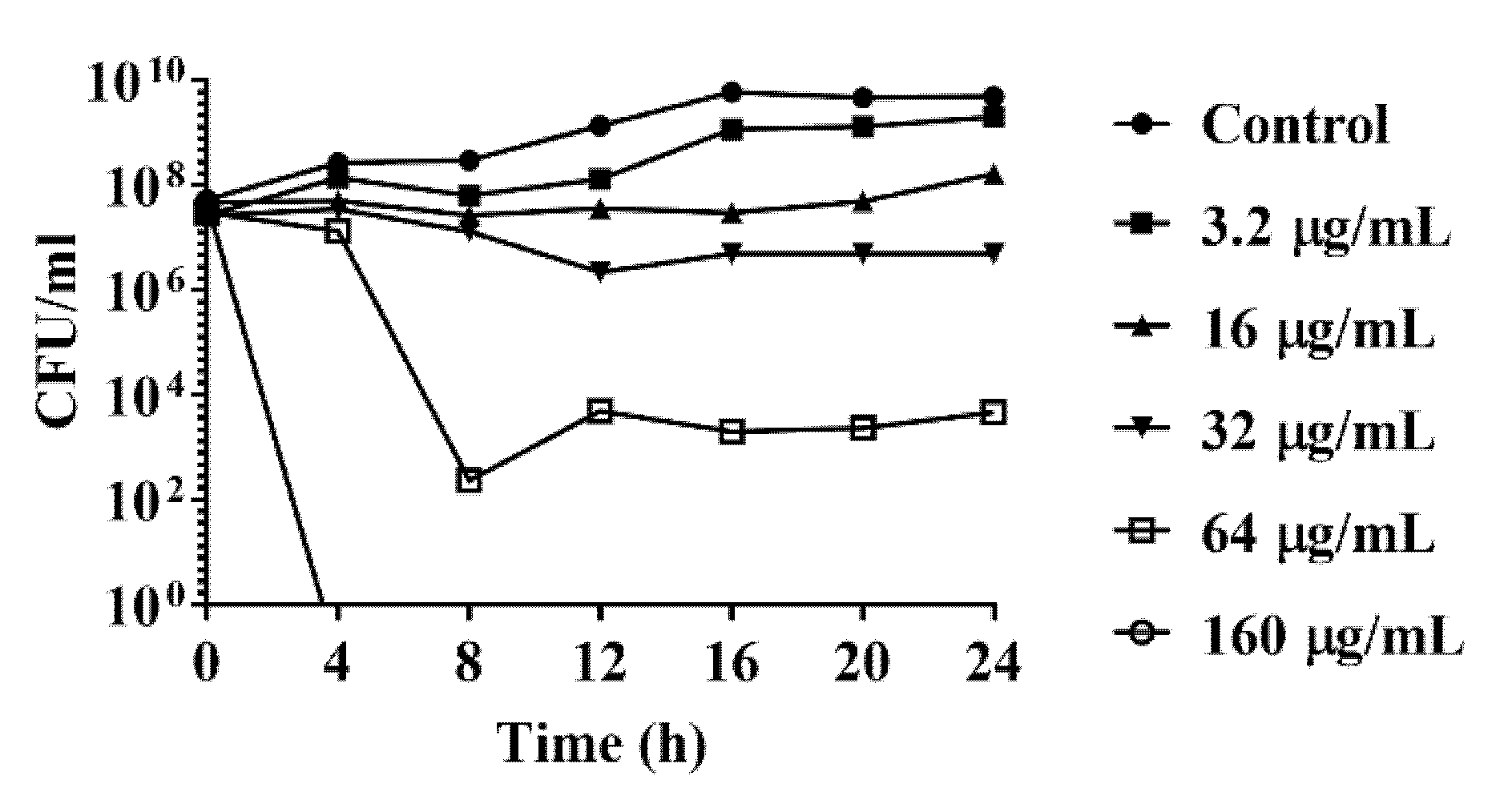
2.1. Antimicrobial Effect of DATS on Campylobacter spp.
2.2. Morphological Changes Caused by DATS
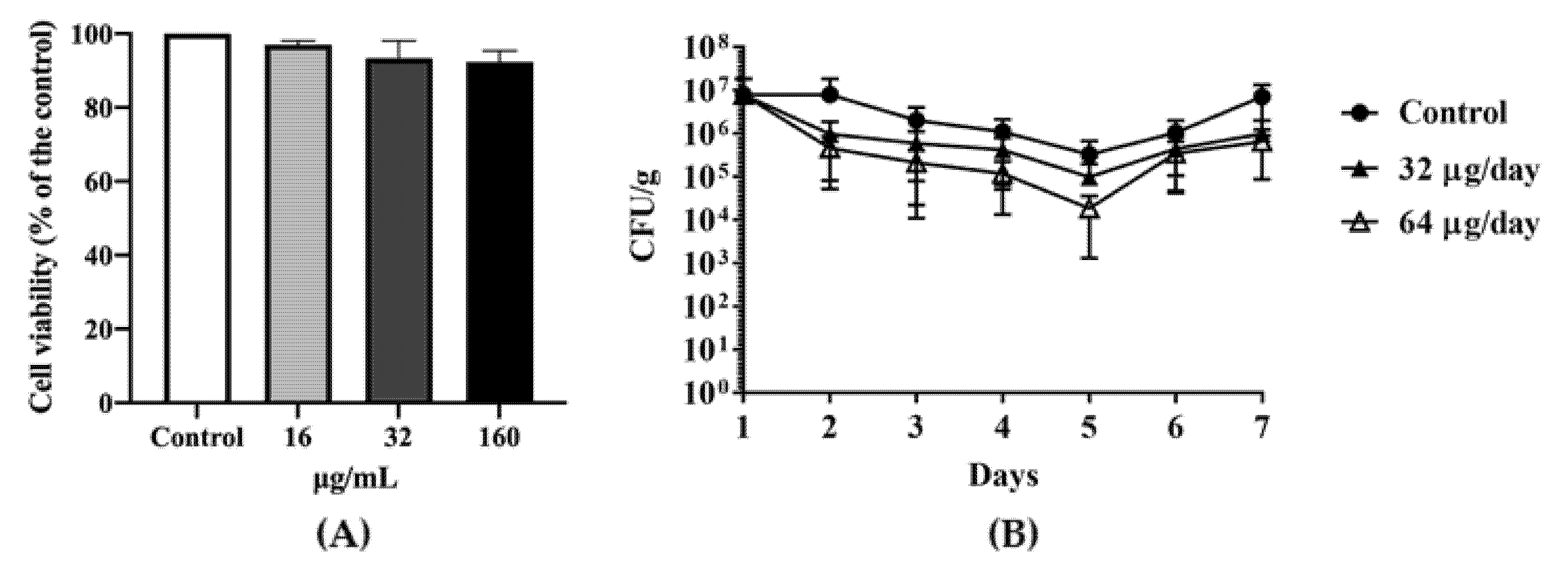
2.3. Antimicrobial Effect of DATS In Vivo
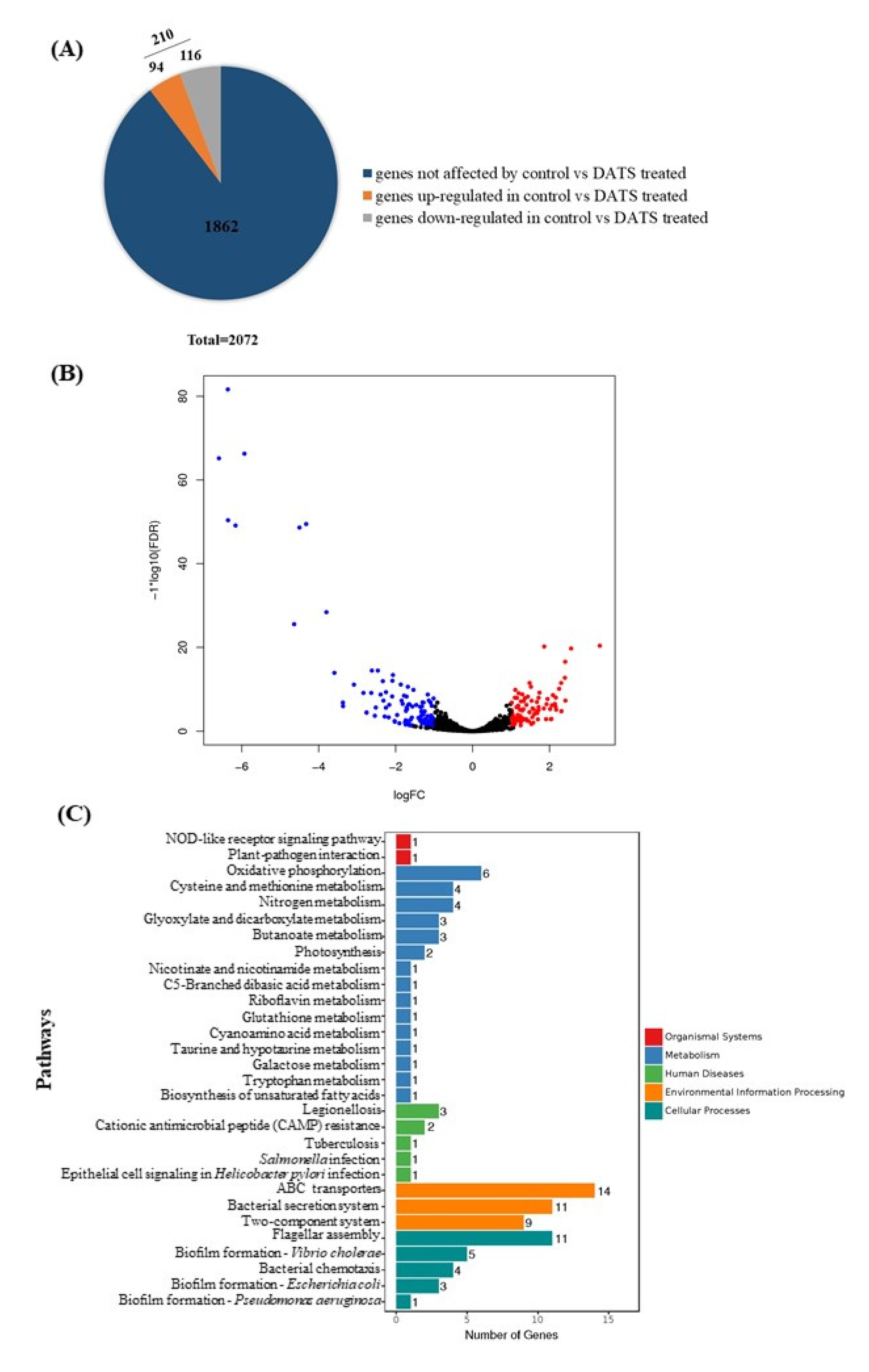
2.4. Transcriptomic Analysis
2.5. The Effect of DATS on the Transmembrane Transporter
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Methods
4.2. Minimum Inhibitory Concentration (MIC) Analysis
4.3. Time Kill Analysis
4.4. Scanning Electron Microscopy
4.5. MTT Cytotoxicity Assay of DATS
4.6. Animal Experiments
4.7. RNA-seq Analysis
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT–PCR)
4.9. Construction of C. jejuni Mutant Strain
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, e05077. [Google Scholar]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar]
- Rasschaert, G.; De Zutter, L.; Herman, L.; Heyndrickx, M. Campylobacter contamination of broilers: The role of transport and slaughterhouse. Int. J. Food Microbiol. 2020, 322, 108564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, M.; Yang, W.; Fang, Y.; Wang, G.; Hou, F. A seventeen-year observation of the antimicrobial susceptibility of clinical Campylobacter jejuni and the molecular mechanisms of erythromycin-resistant isolates in Beijing, China. Int. J. Infect. Dis. 2016, 42, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Malik, U.; Armstrong, D.; Ashworth, M.; Dregan, A.; L’Esperance, V.; McDonnell, L.; Molokhia, M.; White, P. Association between prior antibiotic therapy and subsequent risk of community-acquired infections: A systematic review. J. Antimicrob. Chemother. 2018, 73, 287–296. [Google Scholar] [CrossRef]
- Chekki, R.Z.; Snoussi, A.; Hamrouni, I.; Bouzouita, N. Chemical composition, antibacterial and antioxidant activities of Tunisian garlic (Allium sativum) essential oil and ethanol extract. Mediterr. J. Chem. 2014, 3, 947–956. [Google Scholar] [CrossRef]
- Nile, S.H.; Nile, A.S.; Keum, Y.S.; Sharma, K. Utilization of quercetin and quercetin glycosides from onion (Allium cepa L.) solid waste as an antioxidant, urease and xanthine oxidase inhibitors. Food Chem. 2017, 235, 119–126. [Google Scholar] [CrossRef]
- Prakash, D.; Singh, B.N.; Upadhyay, G. Antioxidant and free radical scavenging activities of phenols from onion (Allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]
- Santas, J.; Carbo, R.; Gordon, M.; Almajano, M. Comparison of the antioxidant activity of two Spanish onion varieties. Food Chem. 2008, 107, 1210–1216. [Google Scholar] [CrossRef]
- Marchese, A.; Barbieri, R.; Sanches-Silva, A.; Daglia, M.; Nabavi, S.F.; Jafari, N.J.; Izadi, M.; Ajami, M.; Nabavi, S.M. Antifungal and antibacterial activities of allicin: A review. Trends Food Sci. Technol. 2016, 52, 49–56. [Google Scholar] [CrossRef]
- Putnik, P.; Gabric, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Bursac Kovacevic, D. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef]
- Jeremic, J.N.; Jakovljevic, V.L.; Zivkovic, V.I.; Srejovic, I.M.; Bradic, J.V.; Bolevich, S.; Nikolic Turnic, T.R.; Mitrovic, S.L.; Jovicic, N.U.; Tyagi, S.C.; et al. The cardioprotective effects of diallyl trisulfide on diabetic rats with ex vivo induced ischemia/reperfusion injury. Mol. Cell Biochem. 2019, 460, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Casella, S.; Leonardi, M.; Melai, B.; Fratini, F.; Pistelli, L. The role of diallyl sulfides and dipropyl sulfides in the in vitro antimicrobial activity of the essential oil of garlic, Allium sativum L., and leek, Allium porrum L. Phytother. Res. 2013, 27, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.M.; Yin, M.C. In vitro antimicrobial activity of four diallyl sulphides occurring naturally in garlic and Chinese leek oils. J. Med. Microbiol. 2001, 50, 646–649. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Qiu, Z.; Liu, Q.; Huang, Y.; Li, D.; Shen, X.; Fan, K.; Xi, J.; Gu, Y.; Tang, Y.; et al. Converting organosulfur compounds to inorganic polysulfides against resistant bacterial infections. Nat. Commun. 2018, 9, 3713. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Huh, J.E.; Kyung, S.H.; Kyung, K.H. Antimicrobial activity of alk (en) yl sulfides found in essential oils of garlic and onion. Food Sci. Biotechnol. 2004, 13, 235–239. [Google Scholar]
- Ross, Z.; O’Gara, E.A.; Hill, D.J.; Sleightholme, H.; Maslin, D.J. Antimicrobial properties of garlic oil against human enteric bacteria: Evaluation of methodologies and comparisons with garlic oil sulfides and garlic powder. Appl. Environ. Microbiol. 2001, 67, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Bjarnsholt, T.; Skindersoe, M.E.; Hentzer, M.; Kristoffersen, P.; Köte, M.; Nielsen, J.; Eberl, L.; Givskov, M. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J. Bacteriol. 2005, 187, 1799–1814. [Google Scholar] [CrossRef]
- Li, W.R.; Ma, Y.K.; Xie, X.B.; Shi, Q.S.; Wen, X.; Sun, T.L.; Peng, H. Diallyl disulfide from garlic oil inhibits Pseudomonas aeruginosa quorum sensing systems and corresponding virulence factors. Front. Microbiol. 2018, 9, 3222. [Google Scholar] [CrossRef]
- Persson, T.; Hansen, T.H.; Rasmussen, T.B.; Skinderso, M.E.; Givskov, M.; Nielsen, J. Rational design and synthesis of new quorum-sensing inhibitors derived from acylated homoserine lactones and natural products from garlic. Org. Biomol. Chem. 2005, 3, 253–262. [Google Scholar] [CrossRef]
- Cady, N.C.; McKean, K.A.; Behnke, J.; Kubec, R.; Mosier, A.P.; Kasper, S.H.; Burz, D.S.; Musah, R.A. Inhibition of biofilm formation, quorum sensing and infection in Pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS ONE 2012, 7, e38492. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Ma, X.; Xu, Z. Effects of garlic and diallyl trisulfide on the growth, photosynthesis, and alkaline phosphatase activity of the toxic cyanobacterium Microcystis aeruginosa. Environ. Sci. Pollut. R. 2016, 23, 5712–5720. [Google Scholar] [CrossRef]
- Tang, Y.; Jiang, Q.; Tang, H.; Wang, Z.; Yin, Y.; Ren, F.; Kong, L.; Jiao, X.; Huang, J. Characterization and prevalence of Campylobacter spp. from broiler chicken rearing period to the slaughtering process in Eastern China. Front. Vet. Sci. 2020, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, J.K.; Felso, P.; Makszin, L.; Papai, Z.; Horvath, G.; Abraham, H.; Palkovics, T.; Boszormenyi, A.; Emody, L.; Schneider, G. Antimicrobial and virulence-modulating effects of clove essential oil on the foodborne pathogen Campylobacter jejuni. Appl. Environ. Microbiol. 2016, 82, 6158–6166. [Google Scholar] [CrossRef] [PubMed]
- Lanzotti, V.; Scala, F.; Bonanomi, G. Compounds from Allium species with cytotoxic and antimicrobial activity. Phytochem. Rev. 2014, 13, 769–791. [Google Scholar] [CrossRef]
- Najjaa, H.; Neffati, M.; Zouari, S.; Ammar, E. Essential oil composition and antibacterial activity of different extracts of Allium roseum L., a North African endemic species. C. R. Chim. 2007, 10, 820–826. [Google Scholar] [CrossRef]
- Hermans, D.; Van Deun, K.; Messens, W.; Martel, A.; Van Immerseel, F.; Haesebrouck, F.; Rasschaert, G.; Heyndrickx, M.; Pasmans, F. Campylobacter control in poultry by current intervention measures ineffective: Urgent need for intensified fundamental research. Vet. Microbiol. 2011, 152, 219–228. [Google Scholar] [CrossRef]
- Garcia-Diez, J.; Alheiro, J.; Pinto, A.L.; Soares, L.; Falco, V.; Fraqueza, M.J.; Patarata, L. Influence of food characteristics and food additives on the antimicrobial effect of garlic and oregano essential oils. Foods 2017, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.M.; George, A.M. The ABC transporter structure and mechanism: Perspectives on recent research. Cell Mol. Life Sci. 2004, 61, 682–699. [Google Scholar] [CrossRef] [PubMed]
- Noinaj, N.; Guillier, M.; Barnard, T.J.; Buchanan, S.K. TonB-dependent transporters: Regulation, structure, and function. Annu. Rev. Microbiol. 2010, 64, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Cerasi, M.; Liu, J.Z.; Ammendola, S.; Poe, A.J.; Petrarca, P.; Pesciaroli, M.; Pasquali, P.; Raffatellu, M.; Battistoni, A. The ZupT transporter plays an important role in zinc homeostasis and contributes to Salmonella enterica virulence. Metallomics 2014, 6, 845–853. [Google Scholar] [CrossRef]
- Xue, R.; Feng, J.; Ma, L.; Liu, C.; Xian, M.; Konkel, M.E.; Wang, S.; Lu, X. Whole transcriptome sequencing analysis of the synergistic antimicrobial effect of metal oxide nanoparticles and ajoene on Campylobacter jejuni. Front. Microbiol. 2018, 9, 2074. [Google Scholar] [CrossRef]
- CLSI. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; CLSI Guideline M45; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Silvan, J.M.; Zorraquin-Pena, I.; Gonzalez de Llano, D.; Moreno-Arribas, M.V.; Martinez-Rodriguez, A.J. Antibacterial activity of glutathione-stabilized silver nanoparticles against Campylobacter multidrug-resistant strains. Front. Microbiol. 2018, 9, 458. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, J.A.; Jackel, C.; Alter, T.; Janzcyk, P.; Stingl, K.; Knuver, M.T.; Hertwig, S. Reduction of Campylobacter jejuni in broiler chicken by successive application of group II and group III phages. PLoS ONE 2014, 9, e114785. [Google Scholar] [CrossRef]
- Love, M.I.; Anders, S.; Kim, V.; Huber, W. RNA-Seq workflow: Gene-level exploratory analysis and differential expression. F1000 2015, 4, 1070. [Google Scholar] [CrossRef]
- Li, X.; Ren, F.; Cai, G.; Huang, P.; Chai, Q.; Gundogdu, O.; Jiao, X.; Huang, J. Investigating the role of FlhF identifies novel interactions with genes involved in flagellar synthesis in Campylobacter jejuni. Front. Microbiol. 2020, 11, 460. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Host | MICs (μg/mL) | Source |
|---|---|---|---|
| 81-176 | Human | 32 | National Collection of Type Culture |
| YZU01384 | Chicken | 64 | [23] |
| YZU01388 | Chicken | 8 | [23] |
| YZU01391 | Chicken | 32 | [23] |
| YZU01394 | Chicken | 8 | [23] |
| YZU01364 | Chicken | 1 | [23] |
| YZU01367 | Chicken | 32 | [23] |
| YZU01372 | Chicken | 32 | [23] |
| YZU01375 | Chicken | 8 | [23] |
| YZU01378 | Chicken | 32 | [23] |
| YZU01381 | Chicken | 4 | [23] |
| YZU01385 | Chicken | 4 | [23] |
| YZU01389 | Chicken | 2 | [23] |
| YZU01368 | Chicken | 1 | [23] |
| YZU01371 | Chicken | 8 | [23] |
| Gene_ID | Fold Change | p Value | Biological Function |
|---|---|---|---|
| CJJ81176_RS07815 | −3.799 | 1.98 × 10−31 | TonB-system energizer ExbB |
| CJJ81176_RS07825 | −4.637 | 1.71 × 10−28 | Energy transducer TonB |
| CJJ81176_RS07990 | −2.315 | 1.77 × 10−9 | ABC transporter ATP-binding protein |
| CJJ81176_RS08975 | −1.600 | 9.165 × 10−6 | Major facilitator superfamily transporter |
| CJJ81176_RS01275 | −1.681 | 4.55 × 10−13 | Zinc transporter ZupT |
| CJJ81176_RS04670 | −1.134 | 5.1 × 10−4 | Membrane protein insertion efficiency factor YidD |
| CJJ81176_RS05850 | −1.091 | 5.94 × 10−9 | F0F1 ATP synthase subunit A |
| CJJ81176_RS06510 | −2.251 | 1.61 × 10−7 | ABC transporter ATP-binding protein |
| CJJ81176_RS06500 | −3.083 | 1.31 × 10−13 | ABC transporter permease |
| Strain | MIC (μg/mL) |
|---|---|
| 81-176 | 32 |
| 81-176∆RS07815 | 16 |
| 81-176∆RS07825 | 16 |
| 81-176∆RS07990 | 8 |
| 81-176∆RS05850 | 16 |
| 81-176∆RS06500 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Li, F.; Gu, D.; Wang, W.; Huang, J.; Jiao, X. Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter jejuni. Antibiotics 2021, 10, 246. https://doi.org/10.3390/antibiotics10030246
Tang Y, Li F, Gu D, Wang W, Huang J, Jiao X. Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter jejuni. Antibiotics. 2021; 10(3):246. https://doi.org/10.3390/antibiotics10030246
Chicago/Turabian StyleTang, Yuanyue, Fengming Li, Dan Gu, Wenyan Wang, Jinlin Huang, and Xinan Jiao. 2021. "Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter jejuni" Antibiotics 10, no. 3: 246. https://doi.org/10.3390/antibiotics10030246
APA StyleTang, Y., Li, F., Gu, D., Wang, W., Huang, J., & Jiao, X. (2021). Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter jejuni. Antibiotics, 10(3), 246. https://doi.org/10.3390/antibiotics10030246