Cellular Mechanical Phenotypes of Drought-Resistant and Drought-Sensitive Rice Species Distinguished by Double-Resonator Piezoelectric Cytometry Biosensors


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Reagents and Instruments
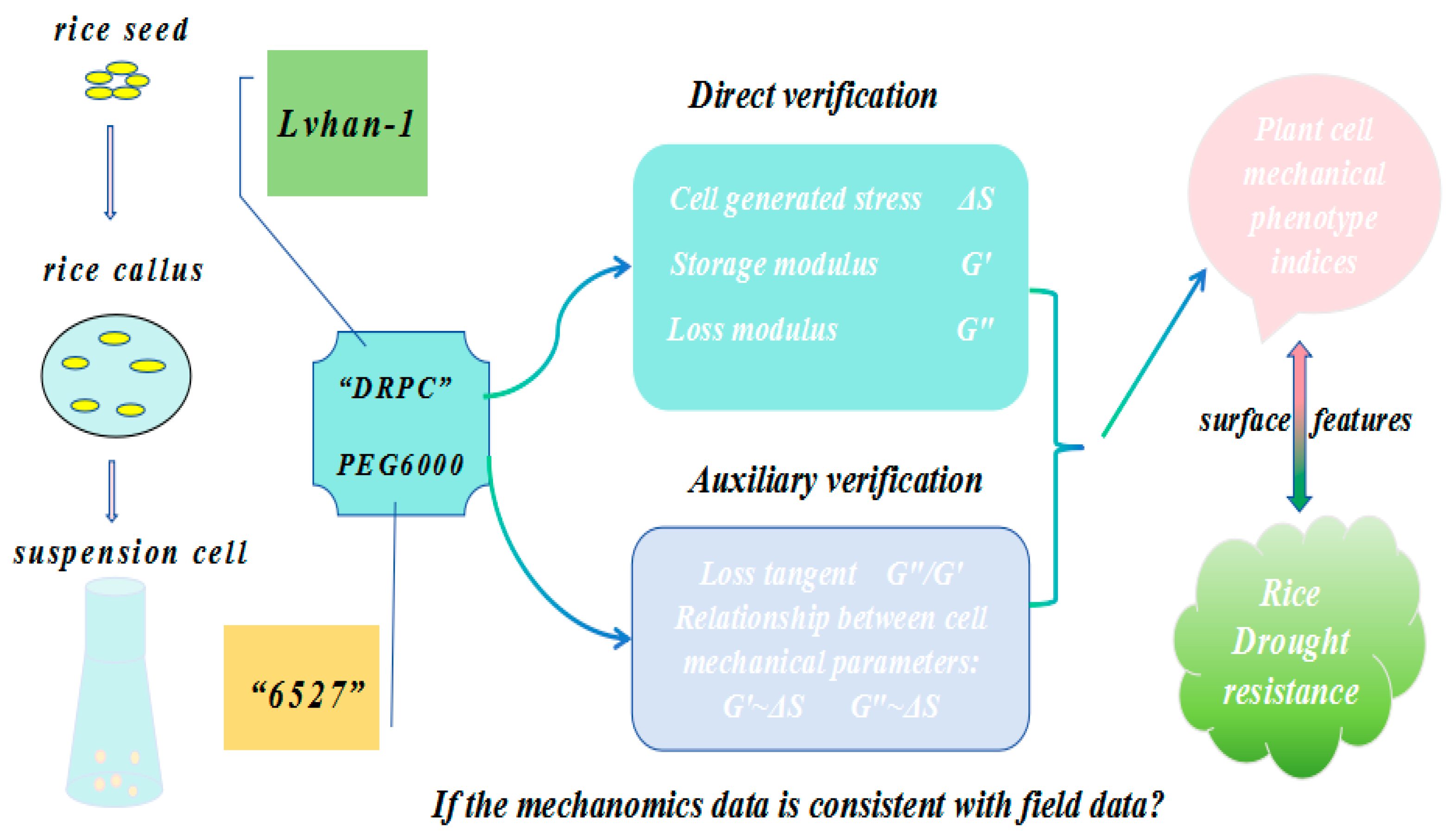
2.3. Experimental Method
2.3.1. Acquisition of Rice Cells
2.3.2. Cleaning and Surface Modification
2.3.3. Monitoring of the Changes in Surface Stress and Viscoelasticity Generated by Rice Cells Under PEG6000 Stress
2.3.4. DRPC Principle and Calculation Formulas for Cellular Mechanical Parameters
2.3.5. Data Analysis
3. Results
3.1. Cell Adhesion Morphology
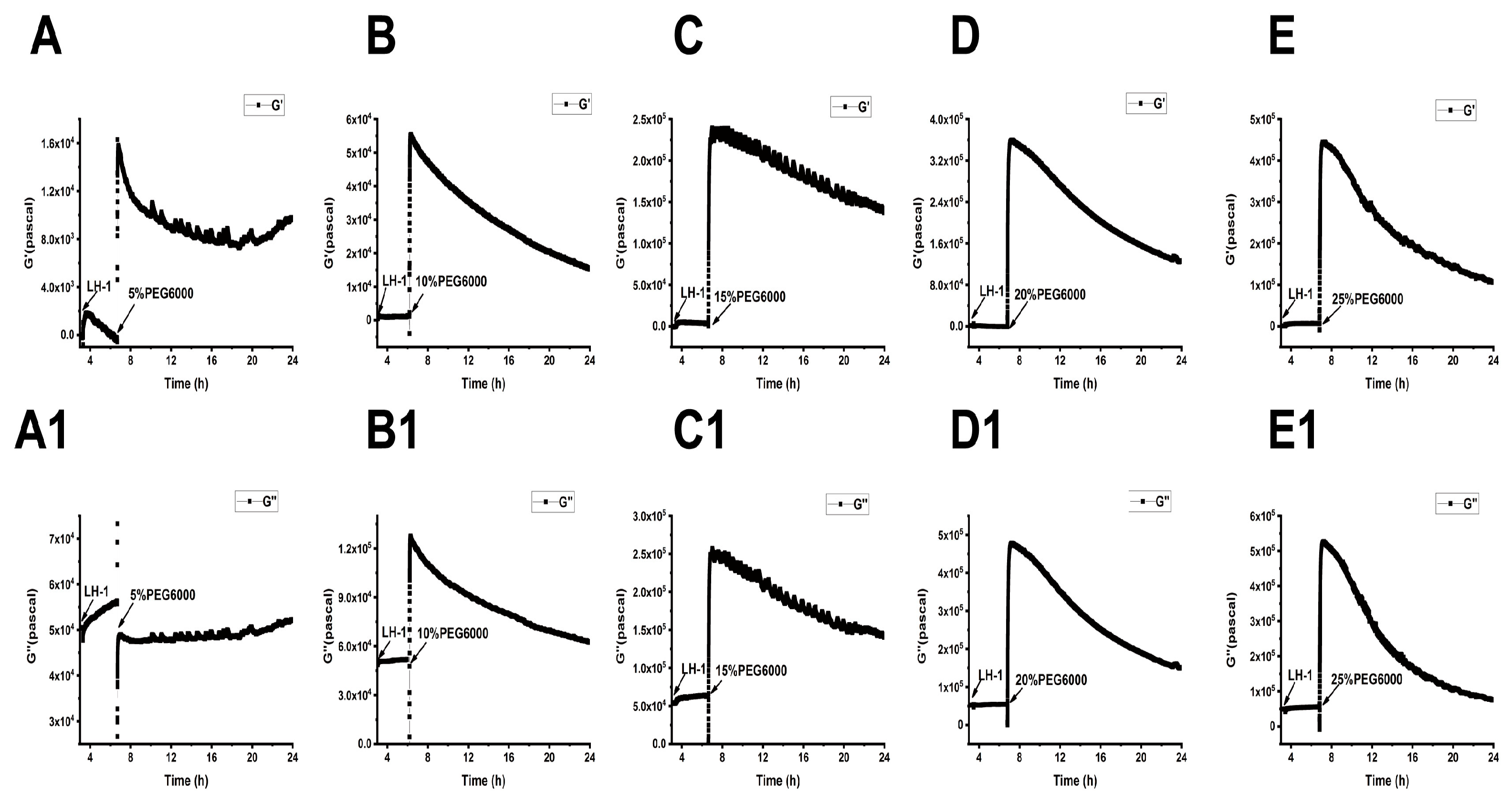
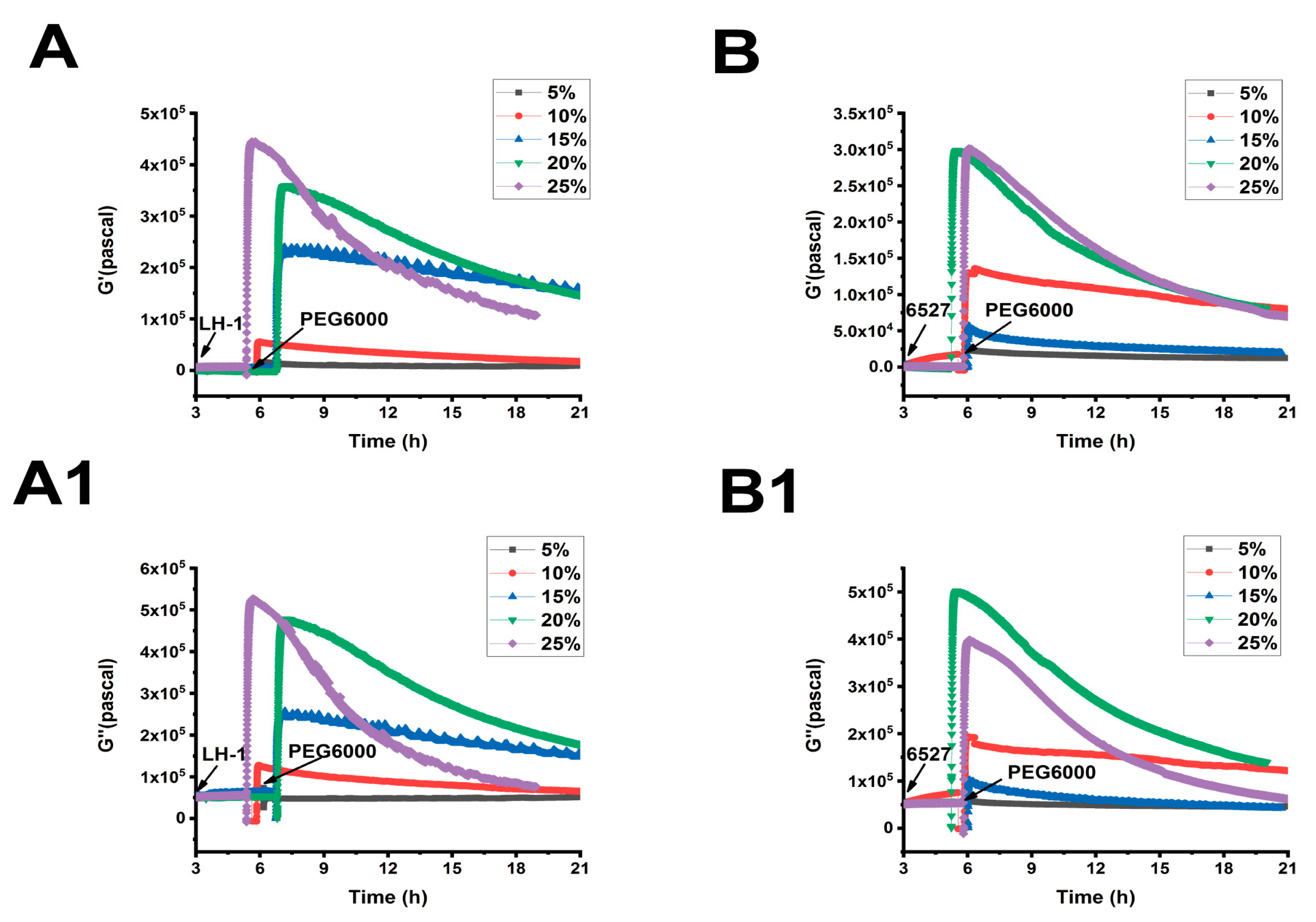
3.2. Lvhan No. 1 and 6527 Cells-Generated Surface Stresses Under PEG6000 Stress
3.3. Changes in Viscoelastic Moduli of Lvhan No. 1 and 6527 Cells Under PEG6000 Stress
4. Discussion
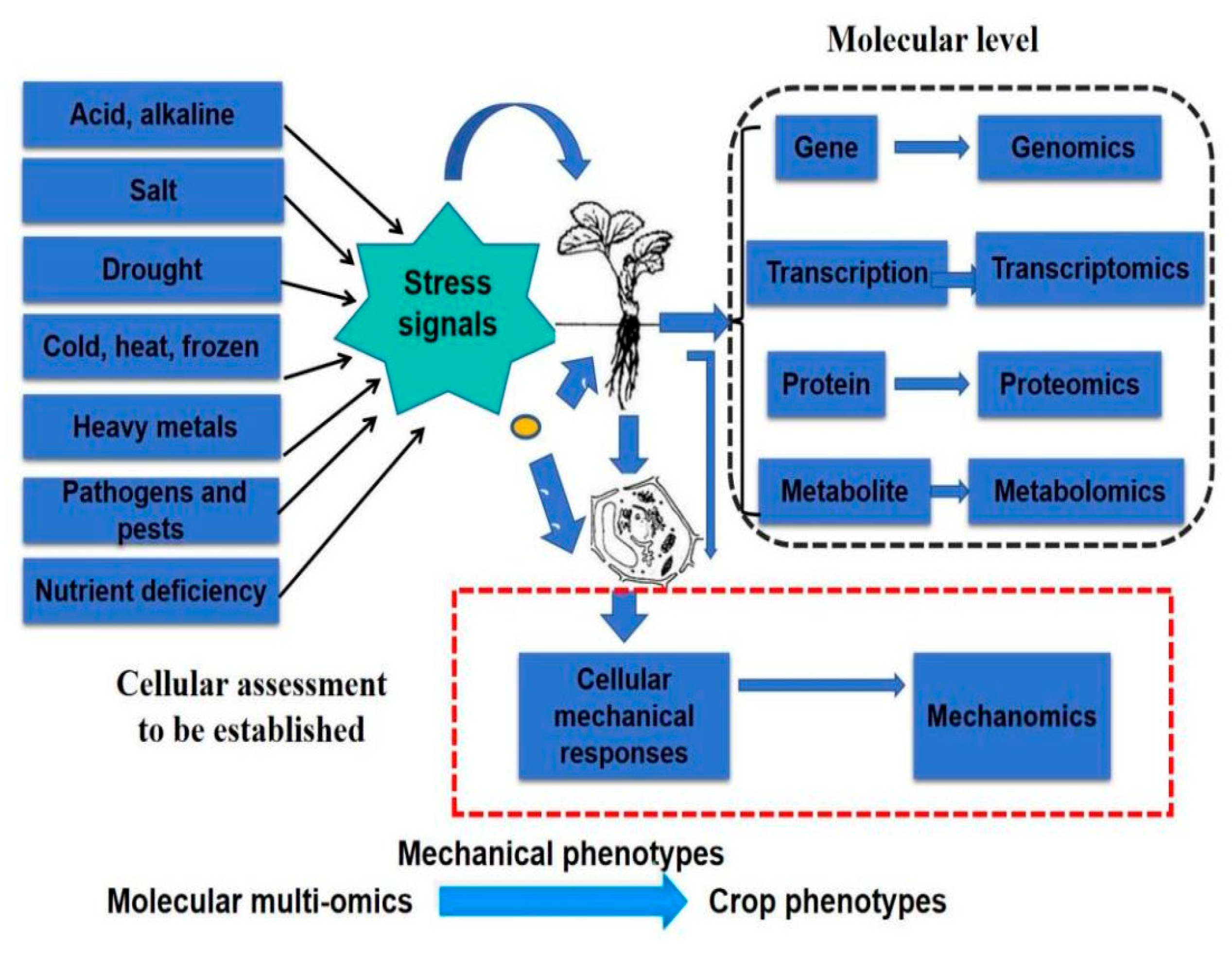
4.1. Evaluation of Crop Resistance at the Cellular Level
4.2. Drought Resistance Evaluation Indexes
4.3. Limitations and Future Developments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhanu, A.N.; Bhandari, H.R.; Singh, M.N.; Srivastava, K. Introduction to Phenomics and its Application in Physiological Breeding. Adv. Plant Physiol. 2017, 17, 200. [Google Scholar]
- Houle, D.; Govindaraju, D.R.; Omholt, S. Phenomics: The next challenge. Nat. Rev. Genet. 2010, 11, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Bassi, F.M.; Sanchez-Garcia, M.; Ortiz, R. What plant breeding may (and may not) look like in 2050? Plant Genome 2024, 17, e20368. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Tao, H.; Xu, S.; Tian, Y.; Li, Z.; Ge, Y.; Zhang, J.; Yang, Y.; Zhou, G.; Deng, X.; Zhang, Z.; et al. Proximal and remote sensing in plant phenomics: 20 years of progress, challenges, and perspectives. Plant Commun. 2022, 3, 100344. [Google Scholar] [CrossRef]
- Verslues, P.E.; Bailey-Serres, J.; Brodersen, C.; Buckley, T.N.; Conti, L.; Christmann, A.; Dinneny, J.R.; Grill, E.; Hayes, S.; Heckman, R.W.; et al. Burning questions for a warming and changing world: 15 unknowns in plant abiotic stress. Plant Cell 2023, 35, 67–108. [Google Scholar] [CrossRef]
- Song, P.; Wang, J.; Guo, X.; Yang, W.; Zhao, C. High-throughput phenotyping: Breaking through the bottleneck in future crop breeding. Crop J. 2021, 9, 633–645. [Google Scholar] [CrossRef]
- Lobos, G.A.; Estrada, F.; Del Pozo, A.; Romero-Bravo, S.; Astudillo, C.A.; Mora-Poblete, F. Challenges for a massive implementation of phenomics in plant breeding programs. In High-Throughput Plant Phenotyping: Methods and Protocols; Springer: New York, NY, USA, 2022; pp. 135–157. [Google Scholar]
- Han, Y.; Qi, W.; Chen, J.; Li, Z.; Su, X.; Yeo, F.K.S. Omics-driven crop improvement for stress tolerance, Volume II. Front. Plant Sci. 2024, 15, 151461217. [Google Scholar] [CrossRef]
- Hoseini, M.; Arzani, A. Epigenetic adaptation to drought and salinity in crop plants. J. Plant Mol. Breed. 2023, 11, 1–16. [Google Scholar]
- Großkinsky, D.K.; Svensgaard, J.; Christensen, S.; Roitsch, T. Plant phenomics and the need for physiological phenotyping across scales to narrow the genotype-to-phenotype knowledge gap. J. Exp. Bot. 2015, 66, 5429–5440. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, S.; Du, J.; Huang, G.; Shi, J.; Lu, X.; Wang, J.; Yang, W.; Guo, X.; Zhao, C. Plant microphenotype: From innovative imaging to computational analysis. Plant Biotechnol. J. 2024, 22, 802–818. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Gangurde, S.S.; Sandhu, K.S.; Lv, Y. Omics-assisted crop improvement under abiotic stress conditions. Plant Stress 2024, 14, 100626. [Google Scholar] [CrossRef]
- Zhou, T.; Huang, J.; Xiong, L.; Shen, H.; Huang, F.; Li, W.; Peng, H.; Su, Z.; Pan, W.; Zhao, J.; et al. Real-Time Quantification of Cell Mechanics and Functions by Double Resonator Piezoelectric Cytometry-Theory and Study of Cellular Adhesion of HUVECs. Adv. Mater. Interfaces 2023, 10, 2300048. [Google Scholar] [CrossRef]
- Wang, Y.; Cortes, E.; Huang, R.; Wan, J.; Zhao, J.; Hinz, B.; Damoiseaux, R.; Pushkarsky, I. FLECS technology for high-throughput screening of hypercontractile cellular phenotypes in fibrosis: A function-first approach to anti-fibrotic drug discovery. SLAS Discov. 2024, 29, 100138. [Google Scholar] [CrossRef]
- Zhou, Z.; Peng, H.; Li, J.; Chen, Z.; Huo, J.; Zhou, T. Real-time monitoring of the contractile properties of H9C2 cardiomyocytes by double resonator piezoelectric cytometry. Anal. Methods 2023, 15, 2839–2852. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, T.; Zhang, Y.; Kong, Z.; Pan, W.; Tan, C. Comparing the Mechanical Properties of Rice Cells and Protoplasts under PEG6000 Drought Stress Using Double Resonator Piezoelectric Cytometry. Biosensors 2024, 14, 303. [Google Scholar] [CrossRef] [PubMed]
- González Ortega-Villaizán, A.; King, E.; Patel, M.K.; Rodríguez-Dobreva, E.; González-Teuber, M.; Ramos, P.; Vicente-Carbajosal, J.; Benitol, B.; Pollmann, S. Identification of a drought stress response module in tomato plants commonly induced by fungal endophytes that confer increased drought tolerance. Plant Mol. Biol. 2025, 115, 7. [Google Scholar] [CrossRef]
- Ren, W.; Shi, Z.; Zhao, Y.; Zhang, Q.; Zhou, M.; Cheng, C.; Liu, M.; Zhao, B.; Guo, Y.; Due, H.; et al. Transcriptional analysis of maize elite inbred line Jing24 and the function of ZmMAPKKK21 in the response to drought stress. Agric. Commun. 2024, 2, 100063. [Google Scholar] [CrossRef]
- Gu, R.; He, H.; Chen, H.; Tian, J. Study on hierarchical regulation of crop irrigation threshold under severe drought conditions. Agric. Water Manag. 2025, 307, 109239. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, S.K.; Muhammad, I.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef]
- Lou, Q.; Chen, Y.; Wang, X.; Zhang, Y.; Gao, T.; Shi, J.; Yan, M.; Feng, F.; Xu, K.; Lin, F. Phenomics-assisted genetic dissection and molecular design of drought resistance in rice. Plant Commun. 2025, 6, 101218. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Wang, S.; Zhang, S.; Zhang, D.; Yang, Q.; Dong, J.; Wang, W.; Feng, C. Breeding and Application of New Rice Variety Luhan No. 1 with Water Saving and Drought Tolerance. J. Anhui Agric. Sci. 2007, 35, 8151. [Google Scholar]
- Ahanger, M.A.; Siddique, K.H.; Ahmad, P. Understanding drought tolerance in plants. Physiol. Plant. 2021, 172, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z. Disputes on hydraulic indexes of plant leaves. Bot. Res. 2023, 12, 56–59. [Google Scholar]
- Guizani, A.; Askri, H.; Amenta, M.L.; Defez, R.; Babay, E.; Bianco, C.; Rapana, N.; Mariella Finetti-Sialer, M.; Gharbi, F. Drought responsiveness in six wheat genotypes: Identification of stress resistance indicators. Front. Plant Sci. 2023, 14, 1232583. [Google Scholar] [CrossRef] [PubMed]
- Bidhendi, A.J.; Geitmann, A. Methods to quantify primary plant cell wall mechanics. J. Exp. Bot. 2019, 70, 3615–3648. [Google Scholar] [CrossRef]
- Fricke, W. Turgor pressure. In eLS; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Lintilhac, P.M.; Wei, C.; Tanguay, J.J.; Outwater, J.O. Ball tonometry: A rapid, non-destructive method for measuring cell turgor pressure in thin-walled plant cells. J. Plant Growth Regul. 2000, 19, 90–97. [Google Scholar] [CrossRef]
- Beauzamy, L.; Derr, J.; Boudaoud, A. Quantifying hydrostatic pressure in plant cells by using indentation with an atomic force microscope. Biophys. J. 2015, 108, 2448–2456. [Google Scholar] [CrossRef]
- Marx, K.A. Quartz crystal microbalance: A useful tool for studying thin polymer films and complex biomolecular systems at the solution− surface interface. Biomacromolecules 2003, 4, 1099–1120. [Google Scholar] [CrossRef]
- Chen, J.Y.; Penn, L.S.; Xi, J. Quartz crystal microbalance: Sensing cell-substrate adhesion and beyond. Biosens. Bioelectron. 2018, 99, 593–602. [Google Scholar] [CrossRef]
- Deng, J.; Zhou, T.; Hu, J.; Chen, Z.; Xie, J. Viscoelastic Changes and Cold Tolerance Comparison of Arabidopsis and Tobacco Cells Under Low Temperature Stress. Chin. Agric. Sci. Bull. 2017, 33, 19–23. [Google Scholar]
- Yang, H.; Zhou, T.; Pan, W.; Xiao, Y. Application of Double Resonator Piezoelectric Cytometry in Salt Tolerance Evaluation of Rice (Oryza sativa) Varieties. J. Agric. Biotechnol. 2024, 32, 911–925. [Google Scholar]
- Gao, Y.; Lu, M.; Zhao, H.; Yang, X.J.; Wang, H. Short-medium term preservation of rice suspension cells. Acta Agron. Sin. 2013, 39, 2039–2045. [Google Scholar] [CrossRef]
- EerNisse, E.P. Simultaneous thin-film stress and mass-change measurements using quartz resonators. J. Appl. Phys. 1972, 43, 1330–1337. [Google Scholar] [CrossRef]
- Haswell, E.S.; Verslues, P.E. The ongoing search for the molecular basis of plant osmosensing. J. Gen. Physiol. 2015, 145, 389–394. [Google Scholar] [CrossRef]
- Wang, N.; Naruse, K.; Stamenović, D.; Fredberg, J.J.; Mijailovich, S.M.; Tolić-Nørrelykke, I.M.; Polte, T.; Mannix, R.; Ingber, D.E. Mechanical behavior in living cells consistent with the tensegrity model. Proc. Natl. Acad. Sci. USA 2001, 98, 7765–7770. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Alemu, A.; Åstrand, J.; Montesinos-López, O.A.; Sánchez, J.I.Y.; Fernández-Gónzalez, J.; Tadesse, W.; Vetukuri, R.R.; Carlsson, A.S.; Ceplitis, A.; Crossa, J.; et al. Genomic selection in plant breeding: Key factors shaping two decades of progress. Mol. Plant 2024, 17, 552–578. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Atzori, G.; Fabbri, A.; Daccache, A.; Killi, D.; Carli, A.; Montesano, V.; Conte, A.; Balestrini, R.; et al. Plant Physiological Analysis to Overcome Limitations to Plant Phenotyping. Plants 2023, 12, 4015. [Google Scholar] [CrossRef]
- Mazri, M.A.; Koufan, M.; Rham, I.; Radi, H.; Belkoura, I. Use of Tissue Culture Methods to Improve Stress Tolerance in Plants. In New Frontiers in Plant-Environment Interactions: Innovative Technologies and Developments; Springer Nature: Cham, Switzerland, 2023; pp. 425–460. [Google Scholar]
- Jian, L.C.; Wang, H. Plant Cell Biology Under Stress; Science Press: Beijing, China, 2009. [Google Scholar]
- Colin, L.; Martin-Arevalillo, R.; Bovio, S.; Bauer, A.; Vernoux, T.; Caillaud, M.C.; Landrein, B.; Jaillais, Y. Imaging the living plant cell: From probes to quantification. Plant Cell 2022, 34, 247–272. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, X.; Gao, H.; He, T.; Lv, Z.; Zhangzhong, L. When crops meet machine vision: A review and development framework for a low-cost non-destructive online monitoring technology in agricultural production. Agric. Commun. 2024, 2, 100029. [Google Scholar] [CrossRef]
- Kang, H.; Hassan, M.A.; Kang, J.; Luo, Y.; Zhang, H.; Zeng, Y.; Fu, G.; Qin, R.; Xu, D.; Wang, S. Development of rice water-saving and drought resistance quantitative evaluation system of wide water ecological range based on quantitative gradient water control. Front. Plant Sci. 2025, 16, 1548074. [Google Scholar] [CrossRef] [PubMed]
- Esfahanian, E.; Nejadhashemi, A.P.; Abouali, M.; Adhikari, U.; Zhang, Z.; Daneshvar, F.; Herman, M.R. Development and evaluation of a comprehensive drought index. J. Environ. Manag. 2017, 185, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Dahu, N.; Hongning, T.; Qian, Z.; Yueming, Y.; Yiru, L.; Shimei, W. Drought stress in rice: Morpho-physiological and molecular responses and marker-assisted breeding. Front. Plant Sci. 2023, 14, 1215371. [Google Scholar] [CrossRef]
- Schönbeck, L.; Arteaga, M.; Mirza, H.; Coleman, M.; Mitchell, D.; Huang, X.; Ortiz, H.; Santiago, L.S. Plant physiological indicators for optimizing conservation outcomes. Conserv. Physiol. 2023, 11, coad073. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhang, J.; Zhou, L.; Xie, B.; Zhou, J.; Li, C. Comparative evaluation of drought indices for monitoring drought based on remote sensing data. Environ. Sci. Pollut. Res. 2021, 28, 20408–20425. [Google Scholar] [CrossRef]
- Bhandari, U.; Gajurel, A.; Khadka, B.; Thapa, I.; Chand, I.; Bhatta, D.; Poudel, A.; Pandey, M.; Shrestha, S.; Shrestha, J. Morpho-physiological and biochemical response of rice (Oryza sativa L.) to drought stress: A review. Heliyon 2023, 9, e13744. [Google Scholar] [CrossRef]
- Kasprowicz, A.; Smolarkiewicz, M.; Wierzchowiecka, M.; Michalak, M.; Wojtaszek, P. Introduction: Tensegral world of plants. In Mechanical Integration of Plant Cells and Plants; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–25. [Google Scholar]
- Shen, H.; Zhou, T.; Hu, J. A high-throughput QCM chip configuration for the study of living cells and cell-drug interactions. Anal. Bioanal. Chem. 2017, 409, 6463–6473. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters of Cellular Mechanical Phenotypes | Drought Resistance Significance | Possible Relationship with Cellular Structures |
|---|---|---|
| Surface stress ΔS generated by cells | (1) Under normal isotonic conditions, cells are in force balance (mechanical homeostasis), with ΔS near zero (ΔS ≈ 0); under PEG6000 drought stress, cells produce contraction stress, ΔS < 0. Under the same concentration of PEG6000 stress, the more negative ΔS is, the worse the drought resistance is. (2) The higher the PEG6000 concentration at which ΔS produces irreversible plastic changes, the better the drought resistance is. (3) Under high concentration PEG6000 stress, cells produce transient tensile stress (ΔS > 0) and have good drought resistance. | When the PEG6000 concentration exceeds a certain value, ΔS changes monotonically over time and shows no recovery trend, indicating that irreversible plastic stress has occurred in the cell structure. The higher the PEG6000 concentration that produces irreversible plastic stress, the better the drought resistance. If cells produce tensile stress (ΔS > 0) under high concentration PEG6000 stress, it indicates that the structure and function of the Hechtian strands are normal and the drought resistance is good; if ΔS < 0, it indicates that the structure of the Hechtian strands is very weak or has undergone irreversible damage or rupture, and the drought resistance is poor. |
| Cell viscoelastic function indexes G′, G″, G″/G′ | (1) Under drought stress, the storage modulus G′ and loss modulus G″ of cells increase. Similarly, under PEG6000 concentration stress, the greater the increase in G′ and G″ (the higher G′ and G″), the better the cell drought resistance. (2) In the entire PEG6000 concentration range, do the maximum values of G′ and G″ increase monotonically with the increase in PEG6000 concentration? If so, the cells show a good linear viscoelastic range and have good cell drought resistance; if not, under high concentrations of PEG6000, the cells cannot maintain a good linear viscoelastic range due to the destruction of cell structural integrity. (3) The larger the G″/G′, the softer the cell, and the smaller the G″/G′, the harder the cell. In the same PEG6000 concentration range, the larger the G″/G′ range, that is, the better the regulation function of the cell to soften and harden, the wider the regulation range, and the better the cell drought resistance. | The cell storage modulus G′ response under drought stress reflects the change in cell wall hardness. G″/G′ reflects the softness and hardness of the cell. The width of the concentration range of G′ and G″ increases monotonically with the increase in PEG6000 concentration, so the larger the adjustable viscoelastic range of the cell, and the better the integrity of the cell structure. |
| Cell force and viscoelasticity correlation curves G′~ΔS, G″~ΔS | At high PEG6000 concentrations, are there linear G′~ΔS, G″~ΔS relationship curves that conform to the cellular tensegrity model? Yes: the cell wall–plasma membrane–cytoskeleton continuum has good structural integrity and good drought resistance. | At high PEG6000 concentrations, the linear G′~ΔS and G″~ΔS relationships that conform to the cellular tensegrity model indicate that the cell wall–plasma membrane–cytoskeleton continuum has good structural integrity. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, D.; Zhou, T.; Pan, W.; Wang, S.; Hassan, M.A. Cellular Mechanical Phenotypes of Drought-Resistant and Drought-Sensitive Rice Species Distinguished by Double-Resonator Piezoelectric Cytometry Biosensors. Biosensors 2025, 15, 334. https://doi.org/10.3390/bios15060334
Tang D, Zhou T, Pan W, Wang S, Hassan MA. Cellular Mechanical Phenotypes of Drought-Resistant and Drought-Sensitive Rice Species Distinguished by Double-Resonator Piezoelectric Cytometry Biosensors. Biosensors. 2025; 15(6):334. https://doi.org/10.3390/bios15060334
Chicago/Turabian StyleTang, Ding, Tiean Zhou, Weisong Pan, Shimei Wang, and Muhammad Ahmad Hassan. 2025. "Cellular Mechanical Phenotypes of Drought-Resistant and Drought-Sensitive Rice Species Distinguished by Double-Resonator Piezoelectric Cytometry Biosensors" Biosensors 15, no. 6: 334. https://doi.org/10.3390/bios15060334
APA StyleTang, D., Zhou, T., Pan, W., Wang, S., & Hassan, M. A. (2025). Cellular Mechanical Phenotypes of Drought-Resistant and Drought-Sensitive Rice Species Distinguished by Double-Resonator Piezoelectric Cytometry Biosensors. Biosensors, 15(6), 334. https://doi.org/10.3390/bios15060334