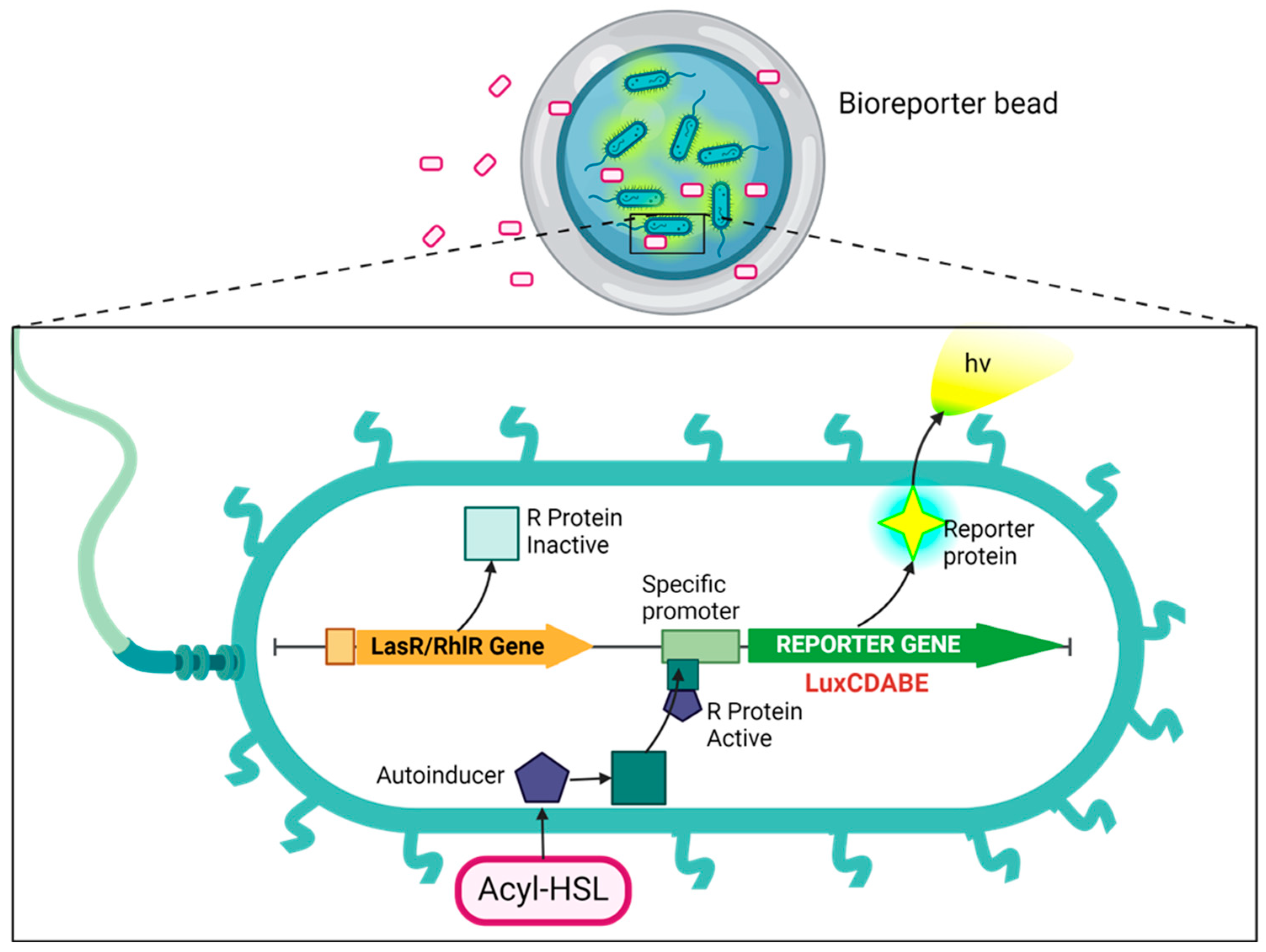
Microbead-Encapsulated Luminescent Bioreporter Screening of P. aeruginosa via Its Secreted Quorum-Sensing Molecules




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
Bacteria Strains
2.2. Synthesis of Microcapsules
2.2.1. Bacteria Strain Growth Conditions
2.2.2. Encapsulation of Bacteria in Single-/Multiple-Layer Microcapsules
2.3. Structural Characterization
2.3.1. X-ray Photoelectron Spectroscopy (XPS) Analysis
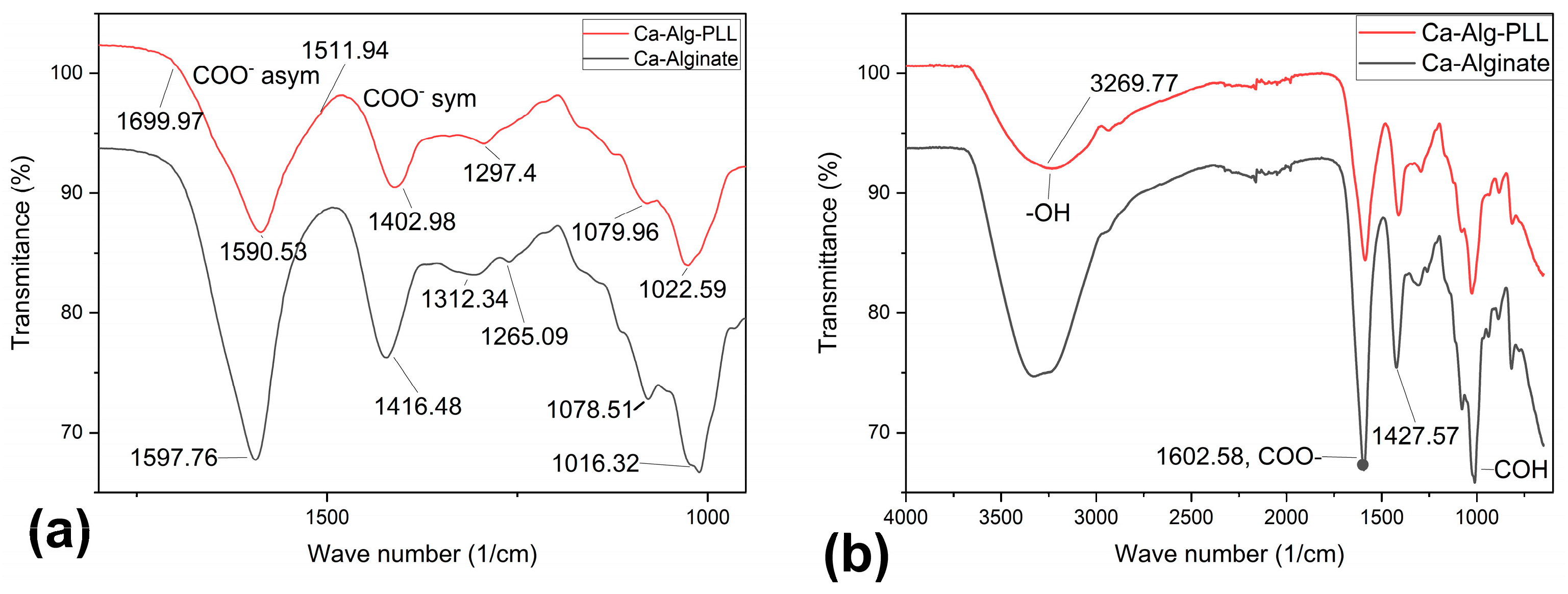
2.3.2. Attenuated Total Reflectance-Fourier Transform Infrared (ATR-FTIR) Analyses
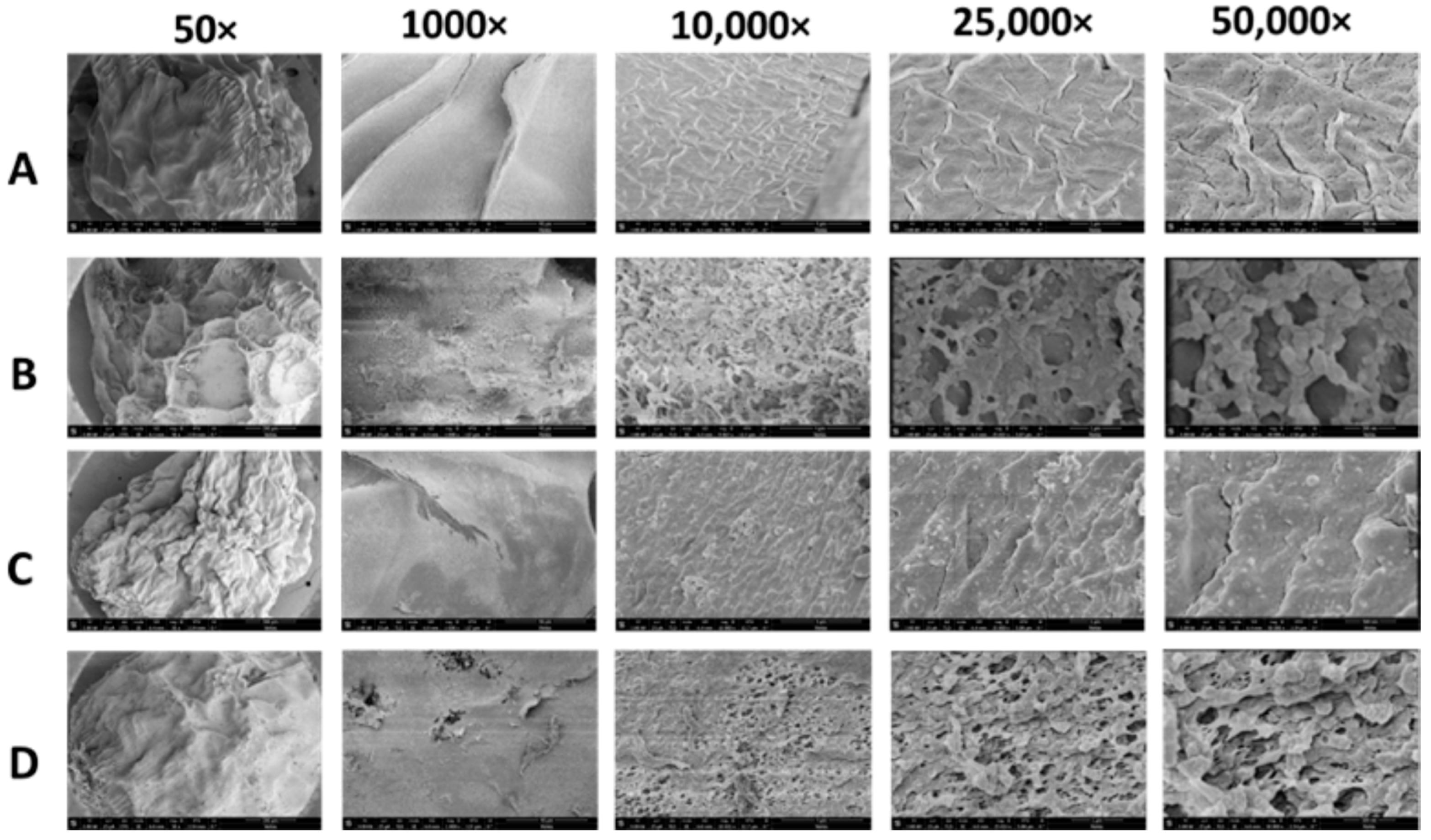
2.3.3. Scanning Electron Microscopy (SEM)
2.4. Bioluminescent Assay
2.4.1. Viability Assay of the Immobilized Bacterial Strains
2.4.2. The Detection of Synthetic AHLs (3-oxo-C12-HSL and C4-HSL)
2.4.3. Detection of AHLs in the Supernatant of Wild Cultures of PAO1 Strains
2.4.4. The Detection of AHLs in the Biofilm Effluent Obtained in the Flow-Cell Experiment
2.4.5. Quorum-Sensing Inhibition Assay
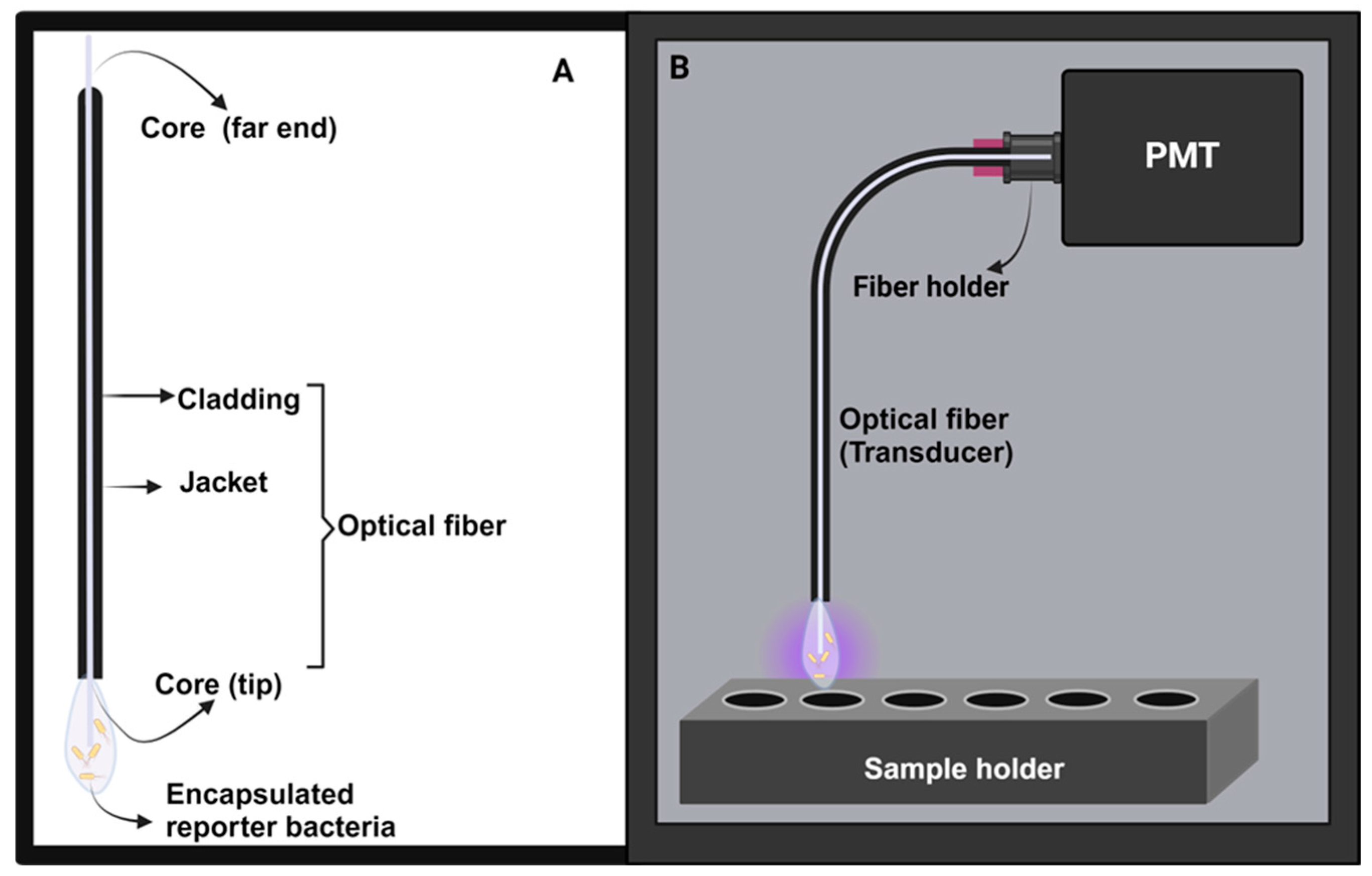
2.5. Development of Optical Biosensors
2.5.1. Instrument Setup
2.5.2. Experimental Probe Setup
2.5.3. Preparation of the Optical-Fiber Surface and Bioluminescent Measurements
2.6. Statistical Analysis
3. Results
3.1. Encapsulation and Optimization
3.1.1. Optimization of Microbead Formations
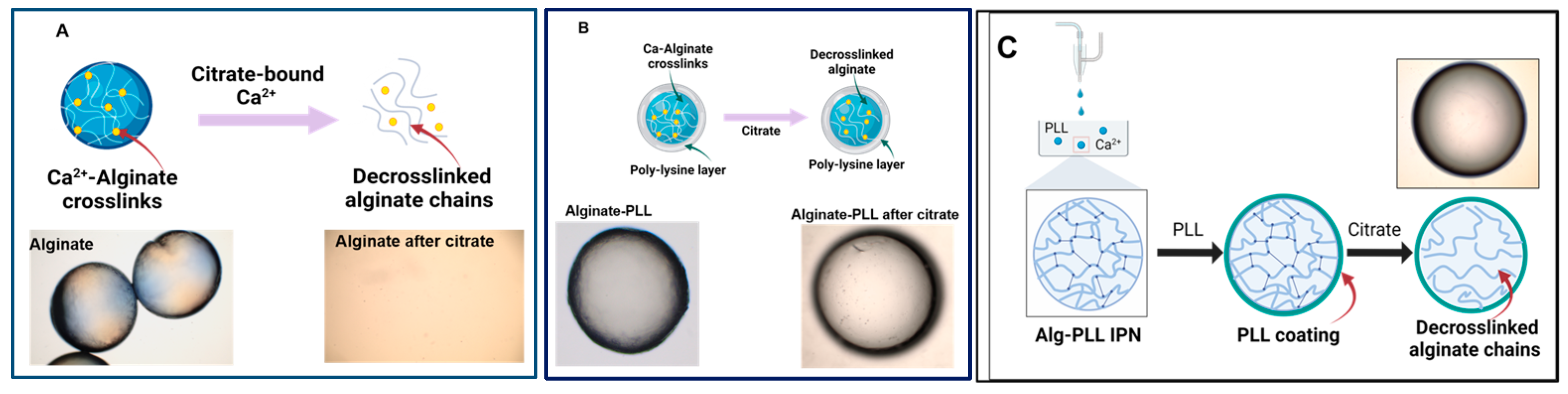
3.1.2. Effect of Encapsulation Methods
3.1.3. ATR-FTIR Data
3.1.4. XPS Analyses
3.1.5. Scanning Electron Microscopy (SEM)
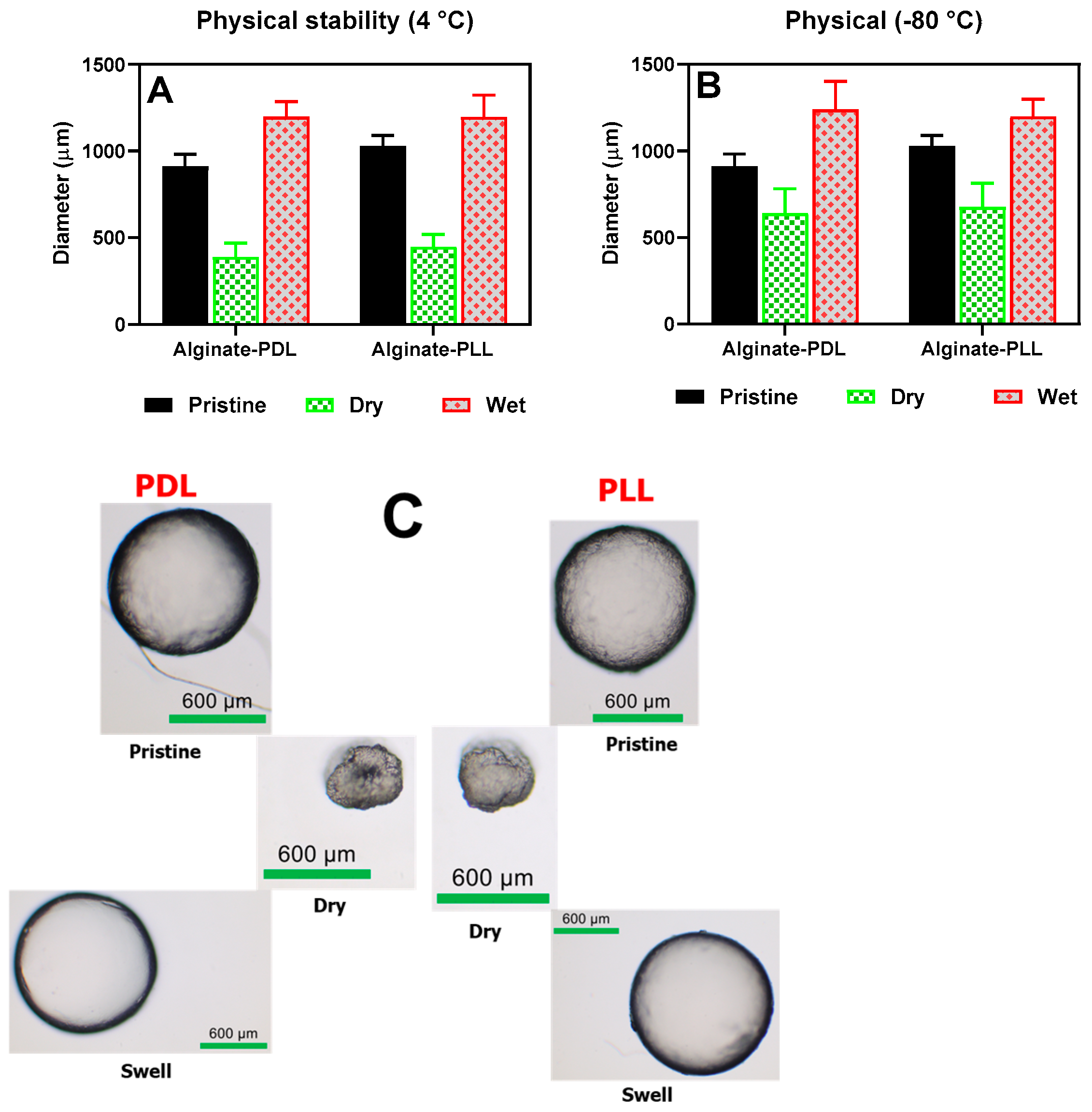
3.1.6. Microbead Stability and Swelling
3.2. Biological Activity Tests
3.2.1. Viability and Bioluminescence
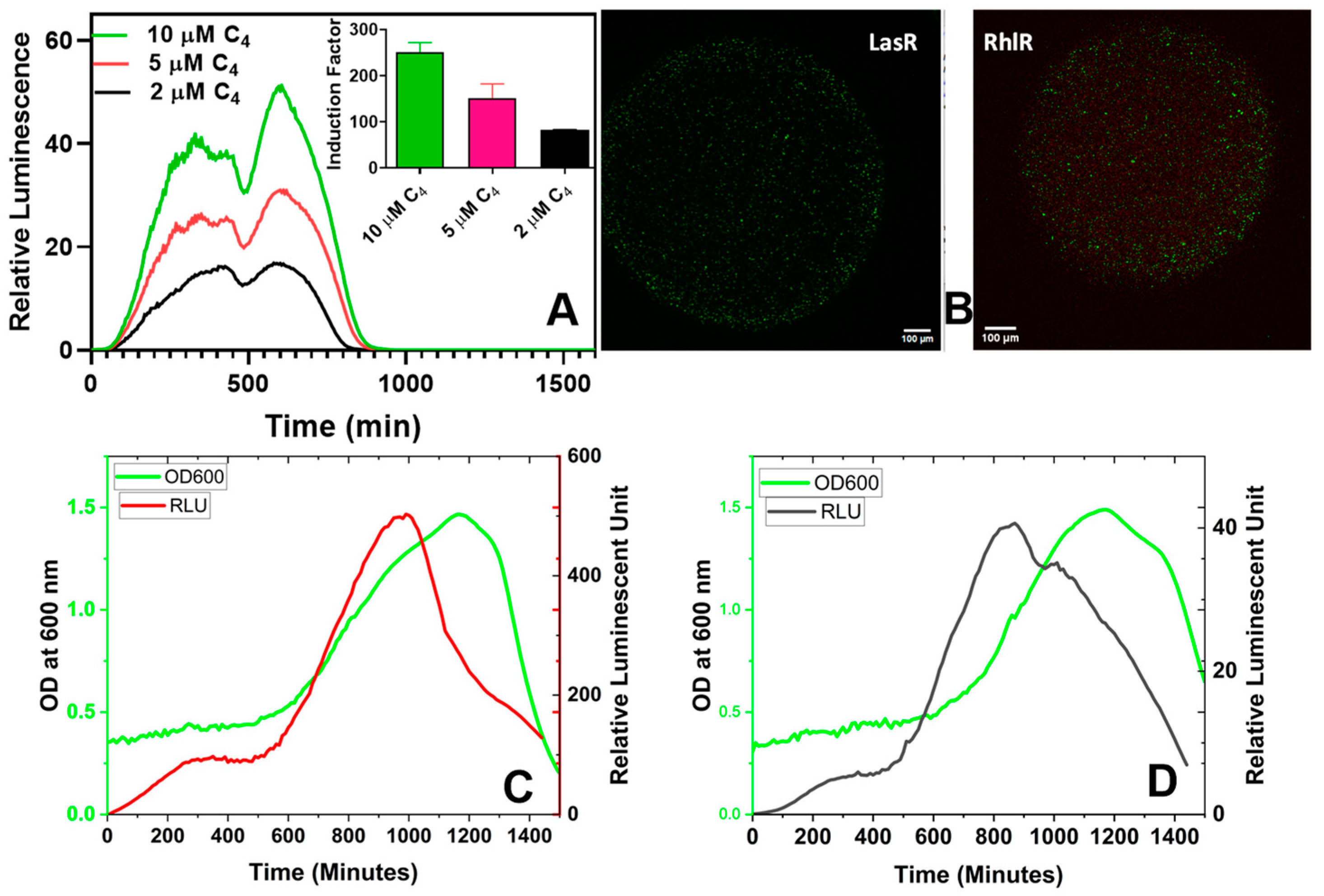
3.2.2. Bioluminescent Response of Alginate Reinforced with L- and D-Isomers of Poly-lysine
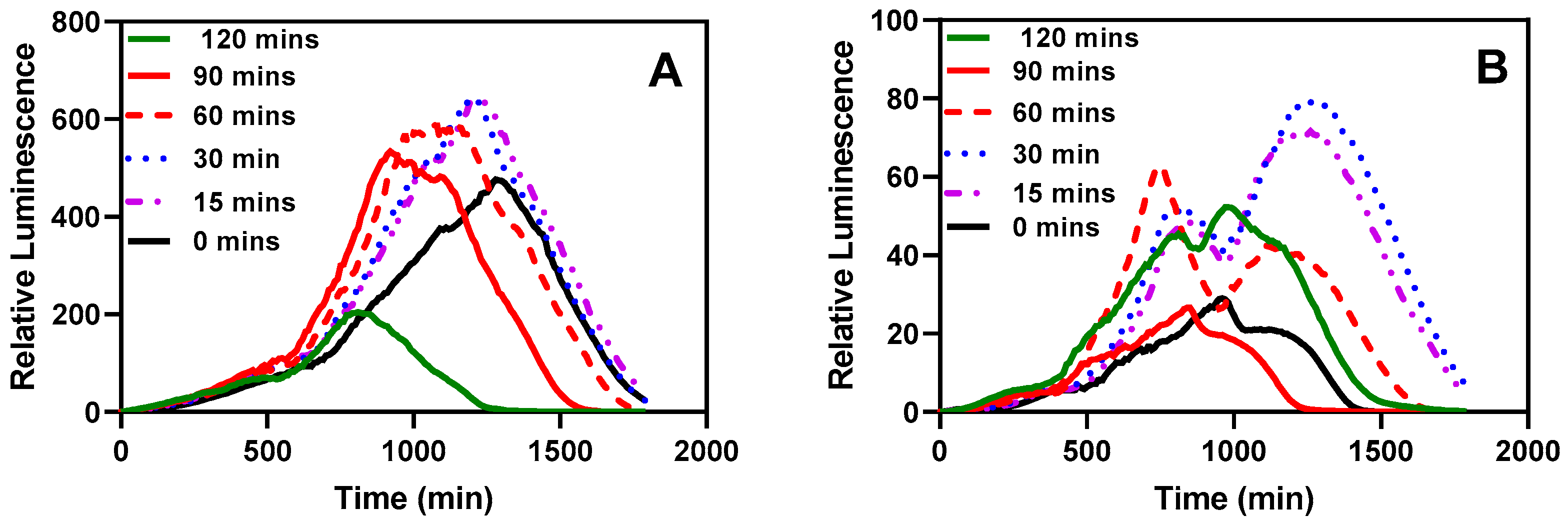
3.2.3. Effect of Activation
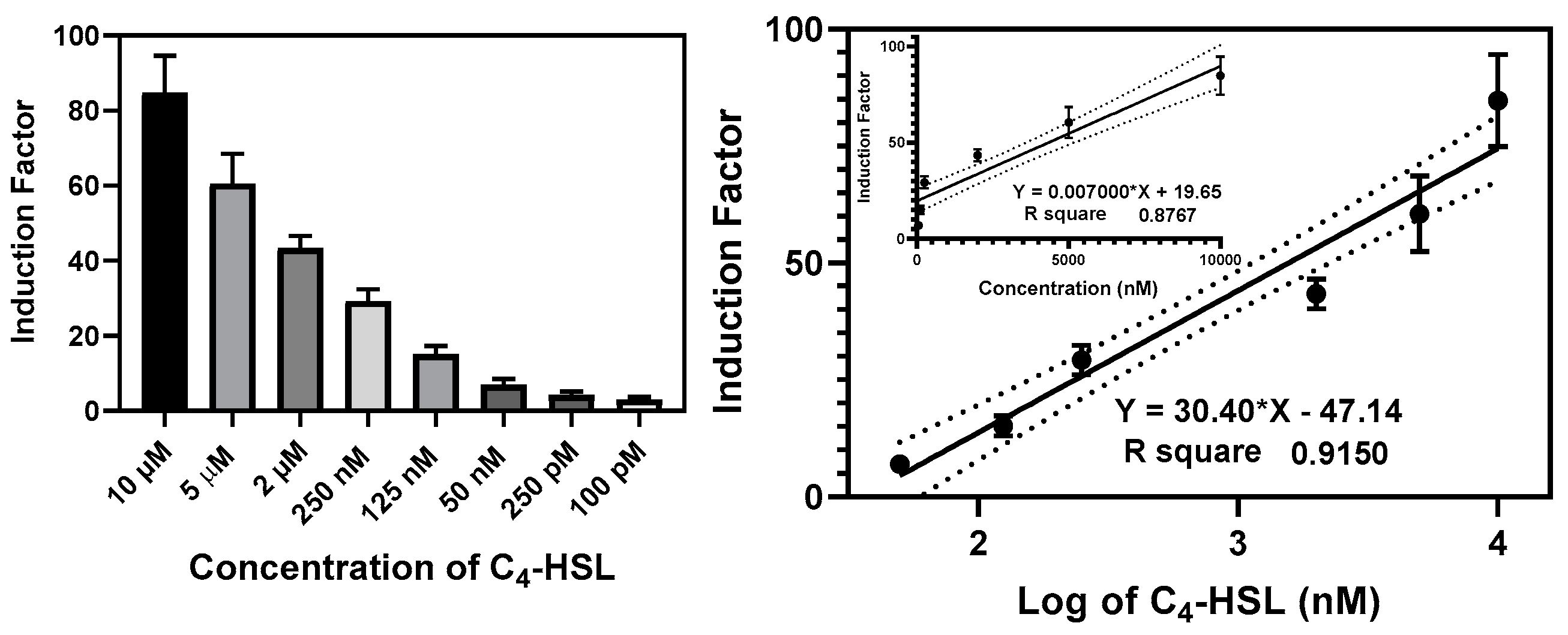
3.2.4. The Construction of the Calibration Curves
3.3. Selectivity and Inhibition Studies
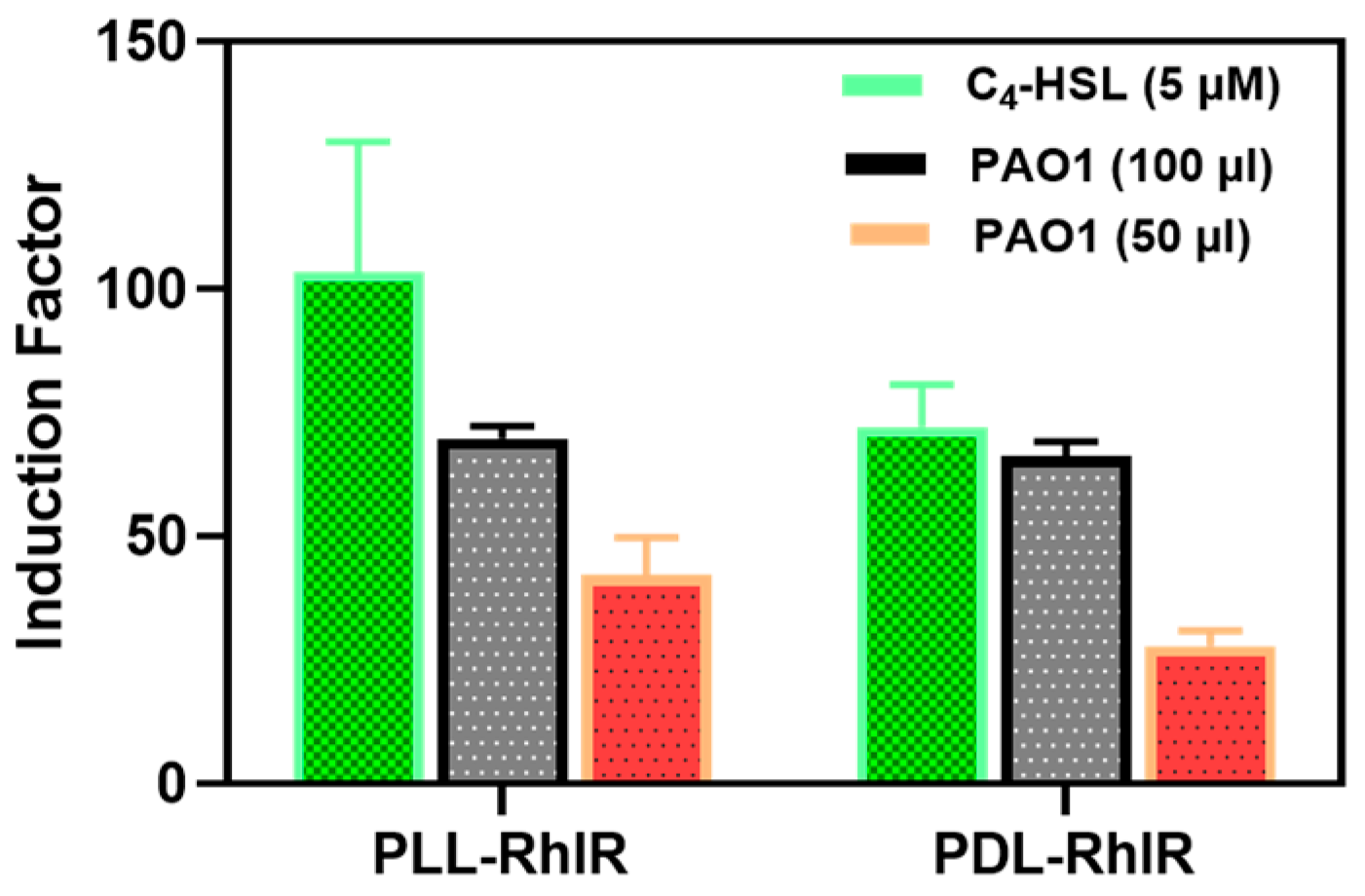
3.3.1. Selectivity of the Bioreporters to the Synthetic and Secreted QS Molecules
3.3.2. Determination of AIs in the Flow-Cell Biofilm Effluent
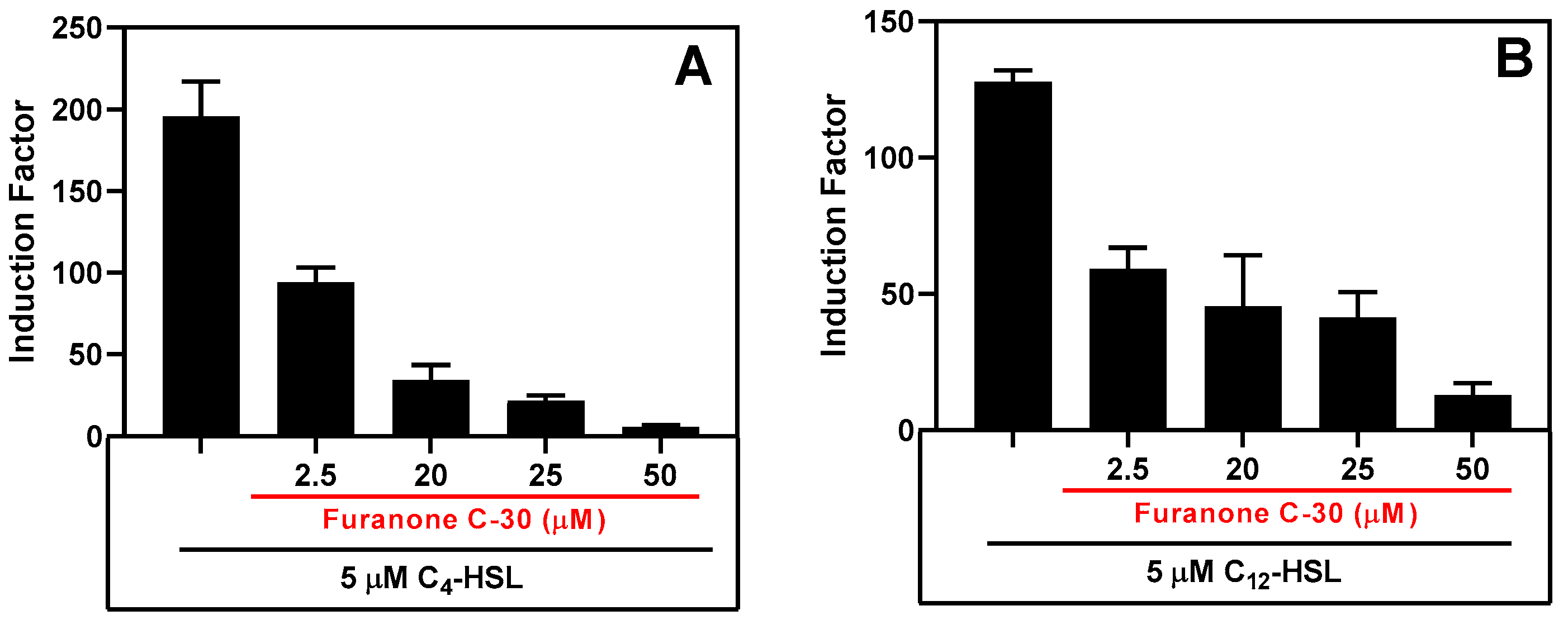
3.3.3. Inhibition Studies
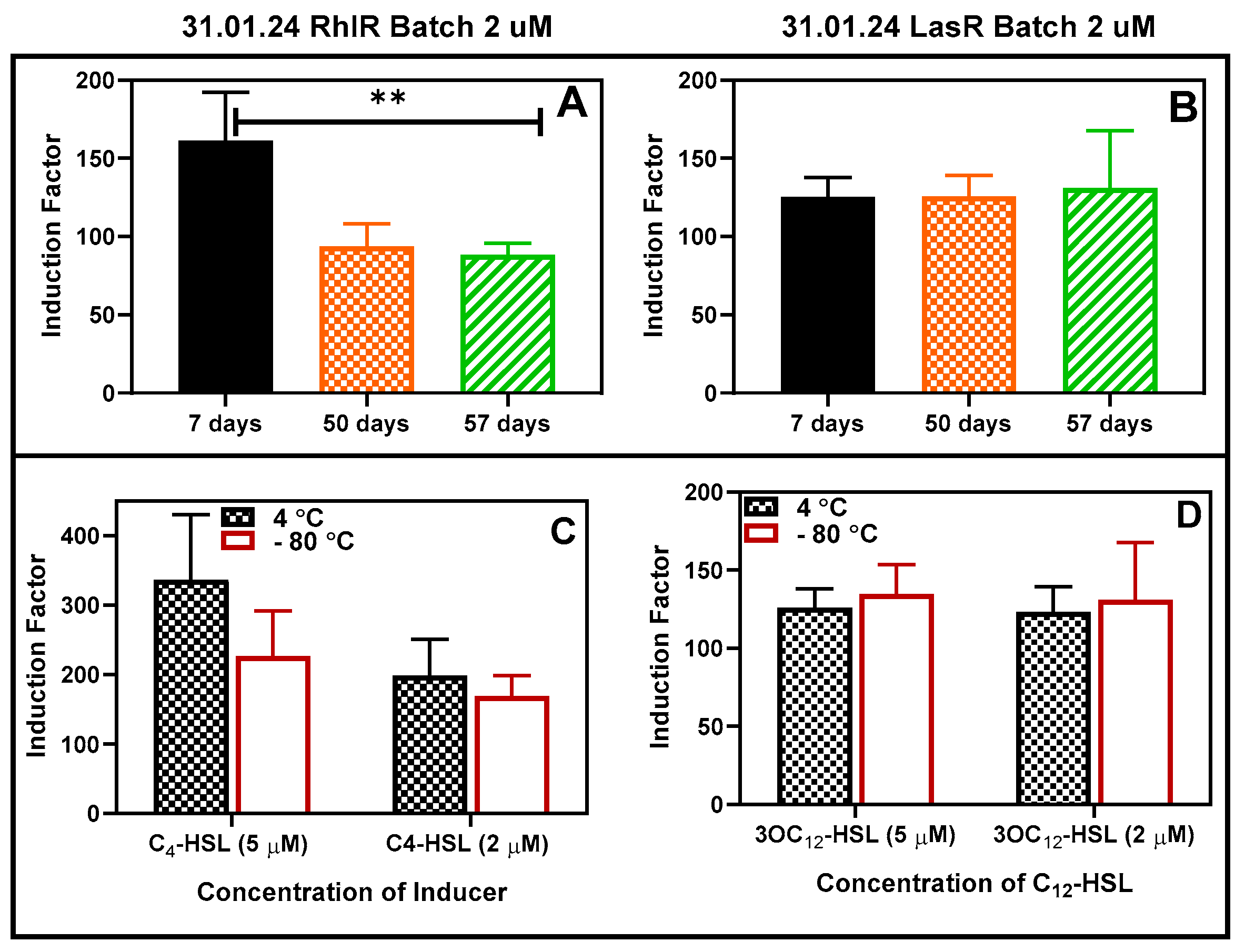
3.4. Storability Assessments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Delden, C.; Iglewski, B.H. Cell-to-cell signaling and Pseudomonas aeruginosa infections. Emerg. Infect. Dis. 1998, 4, 551. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung infections associated with cystic fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Kazemian, H.; Ghafourian, S.; Heidari, H.; Amiri, P.; Yamchi, J.K.; Shavalipour, A.; Houri, H.; Maleki, A.; Sadeghifard, N. Antibacterial, anti-swarming and anti-biofilm formation activities of Chamaemelum nobile against Pseudomonas aeruginosa. Rev. Soc. Bras. Med. Trop. 2015, 48, 432–436. [Google Scholar] [CrossRef]
- Winson, M.K.; Camara, M.; Latifi, A.; Foglino, M.; Chhabra, S.R.; Daykin, M.; Bally, M.; Chapon, V.; Salmond, G.; Bycroft, B.W. Multiple N-acyl-L-homoserine lactone signal molecules regulate production of virulence determinants and secondary metabolites in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 9427–9431. [Google Scholar] [CrossRef]
- Middleton, B.; Rodgers, H.C.; Cámara, M.; Knox, A.J.; Williams, P.; Hardman, A. Direct detection of N-acylhomoserine lactones in cystic fibrosis sputum. FEMS Microbiol. Lett. 2002, 207, 1–7. [Google Scholar] [CrossRef]
- Erickson, D.L.; Endersby, R.; Kirkham, A.; Stuber, K.; Vollman, D.D.; Rabin, H.R.; Mitchell, I.; Storey, D.G. Pseudomonas aeruginosa quorum-sensing systems may control virulence factor expression in the lungs of patients with cystic fibrosis. Infect. Immun. 2002, 70, 1783–1790. [Google Scholar] [CrossRef]
- Favre-Bonté, S.; Pache, J.-C.; Robert, J.; Blanc, D.; Pechère, J.-C.; van Delden, C. Detection of Pseudomonas aeruginosa cell-to-cell signals in lung tissue of cystic fibrosis patients. Microb. Pathog. 2002, 32, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.E.; Visser, M.B.; Schwab, U.; Sokol, P.A. Identification of N-acylhomoserine lactones in mucopurulent respiratory secretions from cystic fibrosis patients. FEMS Microbiol. Lett. 2005, 244, 297–304. [Google Scholar] [CrossRef]
- Barr, H.L.; Halliday, N.; Cámara, M.; Barrett, D.A.; Williams, P.; Forrester, D.L.; Simms, R.; Smyth, A.R.; Honeybourne, D.; Whitehouse, J.L. Pseudomonas aeruginosa quorum sensing molecules correlate with clinical status in cystic fibrosis. Eur. Respir. J. 2015, 46, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Horáček, O.; Portillo, A.E.; Dhaubhadel, U.; Sung, Y.-S.; Readel, E.R.; Kučera, R.; Armstrong, D.W. Comprehensive chiral GC-MS/MS and LC-MS/MS methods for identification and determination of N-acyl homoserine lactones. Talanta 2023, 253, 123957. [Google Scholar] [CrossRef]
- Kumari, A.; Pasini, P.; Daunert, S. Detection of bacterial quorum sensing N-acyl homoserine lactones in clinical samples. Anal. Bioanal. Chem. 2008, 391, 1619–1627. [Google Scholar] [CrossRef] [PubMed]
- Bazhenov, S.; Novoyatlova, U.; Scheglova, E.; Prazdnova, E.; Mazanko, M.; Kessenikh, A.; Kononchuk, O.; Gnuchikh, E.; Liu, Y.; Al Ebrahim, R.; et al. Bacterial lux-biosensors: Constructing, applications, and prospects. Biosens. Bioelectron. X 2023, 13, 100323. [Google Scholar] [CrossRef]
- Van Dyk, T.K.; Rosson, R.A. Photorhabdus luminescens luxCDABE promoter probe vectors. Biolumin. Methods Protoc. 1998, 102, 85–95. [Google Scholar]
- Daunert, S.; Barrett, G.; Feliciano, J.S.; Shetty, R.S.; Shrestha, S.; Smith-Spencer, W. Genetically engineered whole-cell sensing systems: Coupling biological recognition with reporter genes. Chem. Rev. 2000, 100, 2705–2738. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jin, X. Dual crosslinked methacrylated alginate hydrogel micron fibers and tissue constructs for cell biology. Mar. Drugs 2019, 17, 557. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.A.; Markus, V.; Kristollari, K.; Marks, R.S. Alginate-Based Applications in Biotechnology with a Special Mention to Biosensors; IntechOpen: London, UK, 2023. [Google Scholar]
- Mehrotra, T.; Dev, S.; Banerjee, A.; Chatterjee, A.; Singh, R.; Aggarwal, S.J. Use of immobilized bacteria for environmental bioremediation: A review. J. Environ. Chem. Eng. 2021, 9, 105920. [Google Scholar] [CrossRef]
- Budianto, E.; Saepudin, E.; Nasir, M. The encapsulation of Lactobacillus casei probiotic bacteria based on sodium alginate and chitosan. IOP Conf. Ser. Earth Environ. Sci. 2020, 483, 012043. [Google Scholar]
- Strand, B.L.; Mørch, Y.A.; Espevik, T.; Skjåk-Bræk, G. Visualization of alginate–poly-L-lysine–alginate microcapsules by confocal laser scanning microscopy. Biotechnol. Bioeng. 2003, 82, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Nothias, J.-M.; Scavone, A.; Garfinkel, M.; Millis, J.M. Biocompatibility investigation of polyethylene glycol and alginate-poly-L-lysine for islet encapsulation. Asaio J. 2010, 56, 241–245. [Google Scholar] [CrossRef]
- Eltzov, E.; Pavluchkov, V.; Burstin, M.; Marks, R.S. Creation of a fiber optic based biosensor for air toxicity monitoring. Sens. Actuators B Chem. 2011, 155, 859–867. [Google Scholar] [CrossRef]
- Polyak, B.; Bassis, E.; Novodvorets, A.; Belkin, S.; Marks, R.S. Bioluminescent whole cell optical fiber sensor to genotoxicants: System optimization. Sens. Actuators B Chem. 2001, 74, 18–26. [Google Scholar] [CrossRef]
- Marks, R.S.; Bassis, E.; Bychenko, A.; Levine, M.M. Chemiluminescent optical fiber immunosensor for detecting cholera antitoxin. Opt. Eng. 1997, 36, 3258–3264. [Google Scholar] [CrossRef]
- Eltzov, E.; Marks, R.S. Fiber-optic based cell sensors. In Whole Cell Sensing Systems I; Springer: Berlin, Germany, 2010; pp. 131–154. [Google Scholar]
- Ganin, H.; Tang, X.; Meijler, M.M. Inhibition of Pseudomonas aeruginosa quorum sensing by AI-2 analogs. Bioorganic Med. Chem. Lett. 2009, 19, 3941–3944. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Surette, M.G. Environmental regulation of Pseudomonas aeruginosa PAO1 Las and Rhl quorum-sensing systems. J. Bacteriol. 2007, 189, 4827–4836. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Rath, M.; Tsao, C.-Y.; Bentley, W.E.; Raghavan, S.R. Single-step synthesis of alginate microgels enveloped with a covalent polymeric shell: A simple way to protect encapsulated cells. ACS Appl. Mater. Interfaces 2021, 13, 18432–18442. [Google Scholar] [CrossRef] [PubMed]
- Gugerli, R.; Cantana, E.; Heinzen, C.; Stockar, U.V.; Marison, I.W. Quantitative study of the production and properties of alginate/poly-L-lysine microcapsules. J. Microencapsul. 2002, 19, 571–590. [Google Scholar] [CrossRef] [PubMed]
- Golberg, K.; Markus, V.; Kagan, B.-E.; Barzanizan, S.; Yaniv, K.; Teralı, K.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A.J.P. Anti-Virulence Activity of 3, 3′-Diindolylmethane (DIM): A Bioactive Cruciferous Phytochemical with Accelerated Wound Healing Benefits. Pharmaceutics 2022, 14, 967. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.; Smith, A.M.; Gbureck, U.; Shelton, R.; Grover, L. Encapsulation of fibroblasts causes accelerated alginate hydrogel degradation. Acta Biomater. 2010, 6, 3649–3656. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.K.; Dusseault, J.; Polizu, S.; Ménard, M.; Hallé, J.-P. Physicochemical model of alginate–poly-l-lysine microcapsules defined at the micrometric/nanometric scale using ATR-FTIR, XPS, and ToF-SIMS. Biomaterials 2005, 26, 6950–6961. [Google Scholar] [CrossRef]
- Jin, F.; Liao, S.; Li, W.; Jiang, C.; Wei, Q.; Xia, X.; Wang, Q. Amphiphilic sodium alginate-polylysine hydrogel with high antibacterial efficiency in a wide pH range. Carbohydr. Polym. 2023, 299, 120195. [Google Scholar] [CrossRef]
- de Vos, P.; Hoogmoed, C.G.; Busscher, H.J.J. Chemistry and biocompatibility of alginate-PLL capsules for immunoprotection of mammalian cells. J. Biomed. Mater. Res. 2002, 60, 252–259. [Google Scholar] [CrossRef]
- French, T.; So, P.T.C.; Weaver, D.J., Jr.; Coelho-Sampaio, T.; Gratton, E.; Voss, E.W., Jr.; Carrero, J. Two-photon fluorescence lifetime imaging microscopy of macrophage-mediated antigen processing. J. Microsc. 1997, 185, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Creighton, T.E. Proteins: Structures and Molecular Properties; Macmillan: New York, NY, USA, 1993. [Google Scholar]
- Maurer, P.H. Antigenicity of polypeptides (poly alpha amino acids) XIII. Immunological studies with synthetic polymers containing only D-or D-and L-α-amino acids. J. Exp. Med. 1965, 121, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Sela, M. Immunological studies with synthetic polypeptides. Adv. Immunol. 1966, 5, 29–129. [Google Scholar] [PubMed]
- Gu, M.B.; Mitchell, R.J.; Kim, B.C. Whole-cell-based biosensors for environmental biomonitoring and application. In Biomanufacturing; Springer: Berlin, Germany, 2004; pp. 269–305. [Google Scholar]
- de Vos, P.; van Hoogmoed, C.G.; van Zanten, J.; Netter, S.; Strubbe, J.H.; Busscher, H.J. Long-term biocompatibility, chemistry, and function of microencapsulated pancreatic islets. Biomaterials 2003, 24, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Medina, G.; Juárez, K.; Valderrama, B.; Soberón-Chávez, G. Mechanism of Pseudomonas aeruginosa RhlR transcriptional regulation of the rhlAB promoter. Bacteriology 2003, 185, 5976–5983. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jørgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 1998, 163, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Chugani, S.A.; Whiteley, M.; Lee, K.M.; D’Argenio, D.; Manoil, C.; Greenberg, E.P. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2001, 98, 2752–2757. [Google Scholar] [CrossRef]
- Markus, V.; Golberg, K.; Teralı, K.; Ozer, N.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A. Assessing the molecular targets and mode of action of furanone C-30 on Pseudomonas aeruginosa quorum sensing. Molecules 2021, 26, 1620. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paul, A.A.; Kadosh, Y.S.; Kushmaro, A.; Marks, R.S. Microbead-Encapsulated Luminescent Bioreporter Screening of P. aeruginosa via Its Secreted Quorum-Sensing Molecules. Biosensors 2024, 14, 383. https://doi.org/10.3390/bios14080383
Paul AA, Kadosh YS, Kushmaro A, Marks RS. Microbead-Encapsulated Luminescent Bioreporter Screening of P. aeruginosa via Its Secreted Quorum-Sensing Molecules. Biosensors. 2024; 14(8):383. https://doi.org/10.3390/bios14080383
Chicago/Turabian StylePaul, Abraham Abbey, Yael Schlichter Kadosh, Ariel Kushmaro, and Robert S. Marks. 2024. "Microbead-Encapsulated Luminescent Bioreporter Screening of P. aeruginosa via Its Secreted Quorum-Sensing Molecules" Biosensors 14, no. 8: 383. https://doi.org/10.3390/bios14080383
APA StylePaul, A. A., Kadosh, Y. S., Kushmaro, A., & Marks, R. S. (2024). Microbead-Encapsulated Luminescent Bioreporter Screening of P. aeruginosa via Its Secreted Quorum-Sensing Molecules. Biosensors, 14(8), 383. https://doi.org/10.3390/bios14080383