Tween 80 Improves the Acid-Fast Bacilli Quantification in the Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA)

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
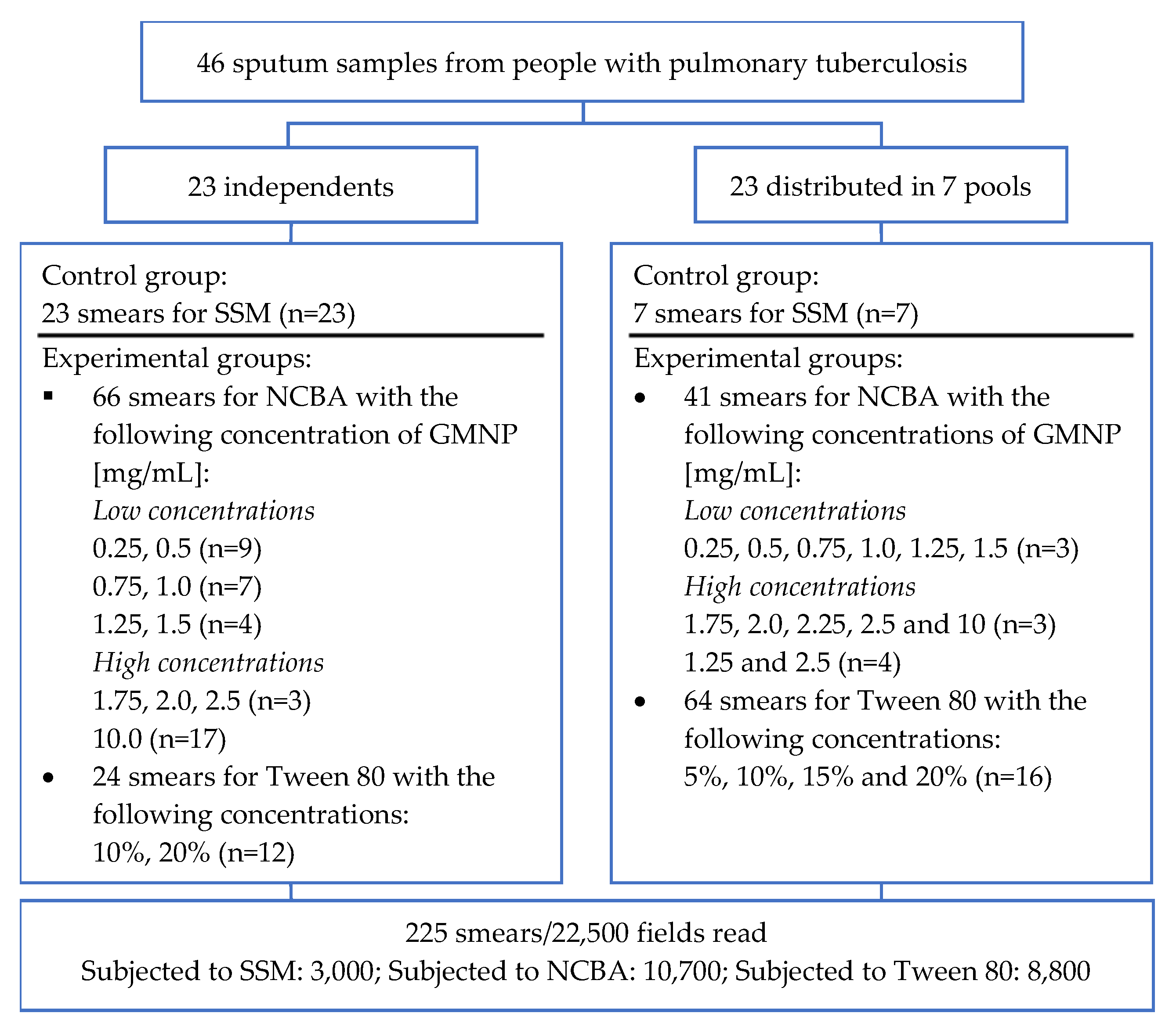
2.2. Clinical Samples
2.3. Processing of Sputum Samples
2.3.1. Sputum Smear Microscopy (SSM)
2.3.2. Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA)
2.3.3. Evaluation of the Tween 80 Effect in the Dispersion of GMNP-AFB Complexes
2.4. AFB Quantification of Acid-Fast Bacilli on Smears
2.5. Statistical Analysis
3. Results
3.1. Analysis of AFB Detection in SSM and NCBA
3.2. Low GMPN Concentrations Improve AFB Quantification
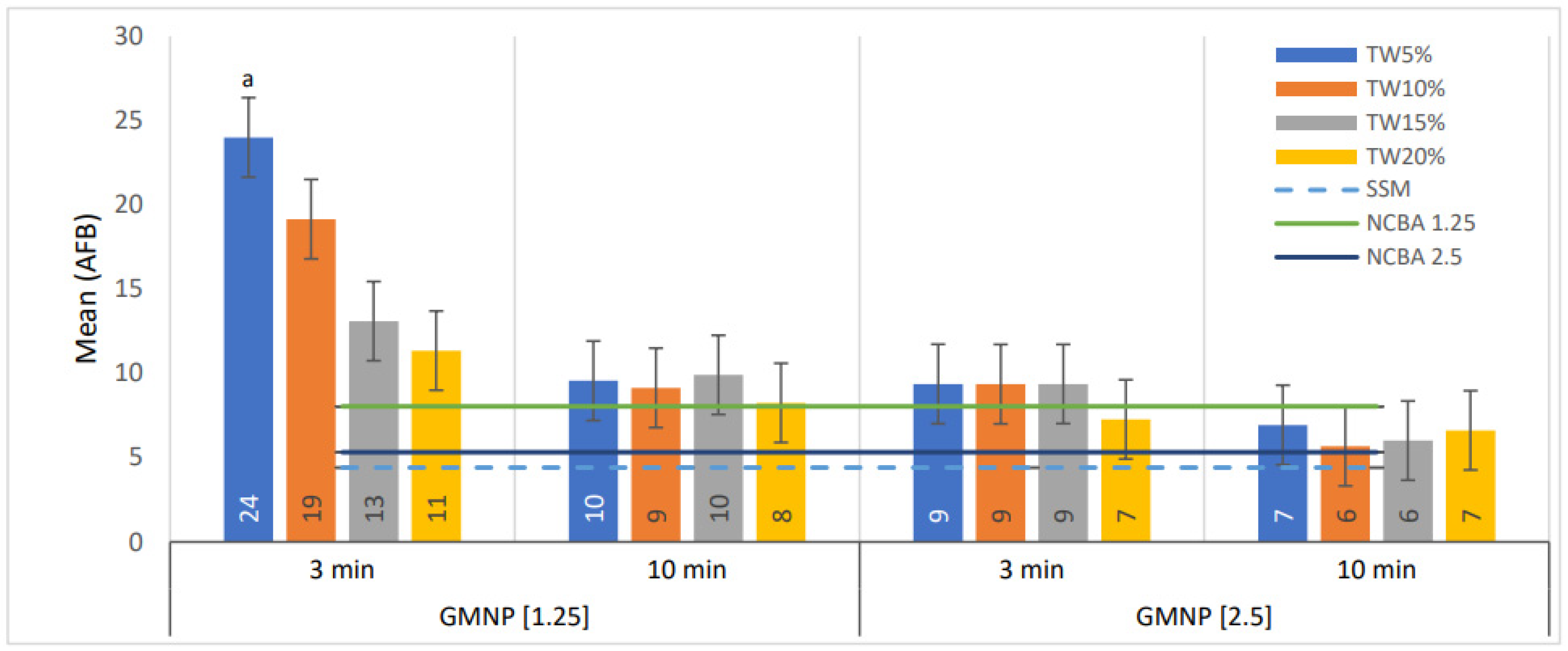
3.3. Tween 80 at Low Concentration Improves the Quantification of AFB in NCBA
4. Discussion
Novelty of the Study and Its Main Application
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report 2020; WHO: Geneva, Switzerland, 2020; ISBN 978-92-4-001313-1.
- WHO. Global Strategy for Tuberculosis Research and Innovation; WHO: Geneva, Switzerland, 2020; ISBN 9789240010017.
- WHO. Global Tuberculosis Report 2021; WHO: Geneva, Switzerland, 2021; ISBN 9780415475976.
- Visca, D.; Ong, C.W.M.; Tiberi, S.; Centis, R.; D’Ambrosio, L.; Chen, B.; Mueller, J.; Mueller, P.; Duarte, R.; Dalcolmo, M.; et al. Tuberculosis and COVID-19 interaction: A review of biological, clinical and public health effects. Pulmonology 2021, 27, 151–165. [Google Scholar] [CrossRef]
- Akhtar, M.; Bretzel, G.; Boulahbal, F.; Dawson, D.; Fattorini, L.; Feldmann, K.; Frieden, T.; Havelková, M.; de Kantor, I.N.; Kim, S.J.; et al. Technical Guide: Sputum Examination for Tuberculosis by Direct Microscopy in Low Income Countries; International Union against Tuberculosis and Lung Disease: Paris, France, 2000; pp. 7–15. [Google Scholar]
- Aït-Khaled, N.; Alarcón, E.; Armengol, R.; Bissell, K.; Boillot, F.; Caminero, J.A.; Chen-Yuan, C.; Clevenbergh, P.; Dlodlo, R.; Enarson, D.A.; et al. Management of Tuberculosis: A Guide to the Essentials of Good Practice; International Union Against Tuberculosis and Lung Disease: Paris, France, 2010; ISBN 9782914365383. [Google Scholar]
- Toman, K. Tuberculosis Case Finding and Chemotherapy. Questions and Answers; World Health Organization: Geneva, Switzerland, 1979.
- Luna-López, F.L. Sesión 1 del 2do. Foro Virtual Internacional de Tuberculosis. Situación Epidemiológica de Tuberculosis en México. Available online: https://www.youtube.com/watch?v=ygjMLDOjImo&list=PLmJwdHM6MQIZ37BHW2elLElvffEed3s_S&index=10&t=4313s (accessed on 14 October 2021).
- WHO. Tuberculosis: Key Facts. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 11 November 2021).
- WHO. Global Tuberculosis Report 2019; WHO: Geneva, Switzerland, 2019; ISBN 978-92-4-156571-4.
- Desikan, P. Sputum smear microscopy in tuberculosis: Is it still relevant? Indian J. Med. Res. 2013, 137, 442–444. [Google Scholar]
- Caulfield, A.J.; Wengenack, N.L. Diagnosis of active tuberculosis disease: From microscopy to molecular techniques. J. Clin. Tuberc. Other Mycobact. Dis. 2016, 4, 33–43. [Google Scholar] [CrossRef]
- Pfyffer, G.E. Mycobacterium: General Characteristics, Laboratory Detection, and Staining Procedures. In Manual of Clinical Microbiology; Jorgensen, J.H., Carroll, K.C., Funke, G., Pfaller, M.A., Landry, M.L., Richter, S.S., Warnock, D.W., Eds.; ASM Press: Washington, DC, USA, 2015; pp. 536–569. [Google Scholar]
- Dorman, S.E.; Schumacher, S.G.; Alland, D.; Nabeta, P.; Armstrong, D.T.; King, B.; Hall, S.L.; Chakravorty, S.; Cirillo, D.M.; Tukvadze, N.; et al. Xpert MTB/RIF Ultra for detection of Mycobacterium tuberculosis and rifampicin resistance: A prospective multicentre diagnostic accuracy study. Lancet Infect. Dis. 2018, 18, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, S.; Simmons, A.M.; Rowneki, M.; Parmar, H.; Cao, Y.; Ryan, J.; Banada, P.P.; Deshpande, S.; Shenai, S.; Gall, A.; et al. The new Xpert MTB/RIF Ultra: Improving detection of Mycobacterium tuberculosis and resistance to rifampin in an assay suitable for point-of-care testing. MBio 2017, 8, e00812–e00817. [Google Scholar] [CrossRef] [Green Version]
- Rasool, G.; Khan, A.M.; Mohy-Ud-Din, R.; Riaz, M. Detection of Mycobacterium tuberculosis in AFB smear-negative sputum specimens through MTB culture and GeneXpert® MTB/RIF assay. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419827174. [Google Scholar] [CrossRef] [Green Version]
- WHO. Meeting Report of a Technical Expert Consultation: Non-Inferiority Analysis of Xpert MTB/RIF Ultra Compared to Xpert MTB/RIF. Available online: https://apps.who.int/iris/handle/10665/254792 (accessed on 19 March 2020).
- FIND; McGill International TB Centre. UNITAID Tuberculosis Diagnostics Market in Select High-Burden Countries: Current Market and Future Opportunities for Novel Diagnostics. Available online: https://www.finddx.org/newsroom/tb-diagnostics-market-in-select-high-burden-countries-current-market-future-opportunities-for-novel-diagnostics/ (accessed on 17 July 2021).
- UNITAID. Diagnostics Technology Landscape, 5th ed.; WHO: Geneva, Switzerland, 2017; pp. 1–90.
- Gordillo-Marroquín, C.; Gómez-Velasco, A.; Sánchez-Pérez, H.J.; Pryg, K.; Shinners, J.; Murray, N.; Muñoz-Jiménez, S.G.; Bencomo-Alerm, A.; Gómez-Bustamante, A.; Jonapá-Gómez, L.; et al. Magnetic nanoparticle-based biosensing assay quantitatively enhances acid-fast bacilli count in paucibacillary pulmonary tuberculosis. Biosensors 2018, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Shrestha, S.; Pote, N.; Alocilja, E.C. Nanoparticle-based biosensing of tuberculosis, an affordable and practical alternative to current methods. Biosensors 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Briceno, R.K.; Sergent, S.R.; Benites, S.M.; Alocilja, E.C. Nanoparticle-based biosensing assay for universally accessible low-cost TB detection with comparable sensitivity as culture. Diagnostics 2019, 9, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birhanu, A.G.; Yimer, S.A.; Kalayou, S.; Riaz, T.; Zegeye, E.D.; Holm-Hansen, C.; Norheim, G.; Aseffa, A.; Abebe, M.; Tønjum, T. Ample glycosylation in membrane and cell envelope proteins may explain the phenotypic diversity and virulence in the Mycobacterium tuberculosis complex. Sci. Rep. 2019, 9, 2927. [Google Scholar] [CrossRef]
- Tonini, L.; Sadet, B.; Stella, A.; Bouyssié, D.; Nigou, J.; Burlet-Schiltz, O.; Rivière, M. Potential plasticity of the mannoprotein repertoire associated to Mycobacterium tuberculosis virulence unveiled by mass spectrometry-based glycoproteomics. Molecules 2020, 25, 2348. [Google Scholar] [CrossRef] [PubMed]
- Matta, L.L.; Alocilja, E.C. Carbohydrate ligands on magnetic nanoparticles for centrifuge-free extraction of pathogenic contaminants in pasteurized milk. J. Food Prot. 2018, 81, 1941–1949. [Google Scholar] [CrossRef]
- Lim, D.; Villame, R.G.; Quiñones, G.J.; de Vera, D.; Notorio, R.; Fernando, L.; Alocilja, E. Alocilja Magnetic Nanoparticles capture Escherichia coli O157:H7 isolates. DIAGNOSTIC Perspect. 2017, 2, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Matta, L.L.; Harrison, J.; Deol, G.S.; Alocilja, E.C. Carbohydrate-functionalized nanobiosensor for rapid extraction of pathogenic bacteria directly from complex liquids with quick detection using cyclic voltammetry. IEEE Trans. Nanotechnol. 2018, 17, 1006–1013. [Google Scholar] [CrossRef]
- Daya, K.S.; Tirumalai, P.S.; Alocilja, E. Impedance and magnetohydrodynamic measurements for label free detection and differentiation of E. Coli and S. Aureus. IEEE Trans. Nanobiosci. 2018, 17, 443–448. [Google Scholar] [CrossRef]
- Kilale, A.M.; Ngadaya, E.; Muhumuza, J.; Kagaruki, G.B.; Lema, Y.L.; Ngowi, B.J.; Mfinanga, S.G.; Hinderaker, S.G. Who has mycobacterial disease? A cross sectional study in agropastoral communities in Tanzania. PLoS ONE 2016, 11, e0153711. [Google Scholar] [CrossRef] [Green Version]
- Mertaniasih, N.; Kusumaningrum, D.; Koendhori, E.; Soedarsono; Kusmiati, T.; Dewi, D. Nontuberculous mycobacterial species and Mycobacterium tuberculosis complex coinfection in patients with pulmonary tuberculosis in Dr. Soetomo Hospital, Surabaya, Indonesia. Int. J. Mycobacteriol. 2017, 6, 9–13. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Z.; Zhang, W.; Jiang, X. Adsorbed Tween 80 is unique in its ability to improve the stability of gold nanoparticles in solutions of biomolecules. Nanoscale 2010, 2, 2114–2119. [Google Scholar] [CrossRef] [PubMed]
- Zahid Hussain, M.; Khan, R.; Ali, R.; Khan, Y. Optical properties of laser ablated ZnO nanoparticles prepared with Tween-80. Mater. Lett. 2014, 122, 147–150. [Google Scholar] [CrossRef]
- Li, H.J.; Zhang, A.Q.; Sui, L.; Qian, D.J.; Chen, M. Hyaluronan/Tween 80-assisted synthesis of silver nanoparticles for biological application. J. Nanoparticle Res. 2015, 17, 111. [Google Scholar] [CrossRef]
- Bekhit, M.; Abu El-naga, M.N.; Sokary, R.; Fahim, R.A.; El-Sawy, N.M. Radiation-induced synthesis of tween 80 stabilized silver nanoparticles for antibacterial applications. J. Environ. Sci. Heal. Part A Toxic Hazardous Subst. Environ. Eng. 2020, 55, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Khan, Y.; Durrani, S.K.; Siddique, M.; Mehmood, M. Hydrothermal synthesis of alpha Fe2O3 nanoparticles capped by Tween-80. Mater. Lett. 2011, 65, 2224–2227. [Google Scholar] [CrossRef]
- Xiao, Y.; Wiesner, M.R. Nanoparticle Aggregation and Deposition in Porous Media. In Engineered Nanoparticles and the Environment: Biophysicochemical Processes and Toxicity; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 224–244. ISBN 9781119275855. [Google Scholar]
- Lin, C.Y.; Yu, C.J.; Lin, Y.H.; Tseng, W.L. Colorimetric sensing of silver (I) and mercury (II) ions based on an assembly of tween 20-stabilized gold nanoparticles. Anal. Chem. 2010, 82, 6830–6837. [Google Scholar] [CrossRef]
- Van-Seuningen, I.; Houdret, N.; Hayem, A.; Davril, M. Strong ionic interactions between mucins and two basic proteins, mucus proteinase inhibitor and lysozyme, in human bronchial secretions. Int. J. Biochem. 1992, 24, 303–311. [Google Scholar] [CrossRef]
- Peng, Y.-P.; Chen, T.-Y.; Wu, C.-Y.; Chang, Y.-C.; Chen, K.-F. Dispersant-modified iron nanoparticles for mobility enhancement and TCE degradation: A comparison study. Environ. Sci. Pollut. Res. 2019, 26, 34157–34166. [Google Scholar] [CrossRef] [PubMed]
- Pietersen, R.D.; du Preez, I.; Loots, D.T.; van Reenen, M.; Beukes, D.; Leisching, G.; Baker, B. Tween 80 induces a carbon flux rerouting in Mycobacterium tuberculosis. J. Microbiol. Methods 2019, 170, 105795. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mahrous, E.A.; Lee, R.E.; Vestling, M.M.; Takayama, K. Novel Polyoxyethylene-Containing Glycolipids Are Synthesized in Corynebacterium matruchotii and Mycobacterium smegmatis Cultured in the Presence of Tween 80. J. Lipids 2011, 2011, 676535. [Google Scholar] [CrossRef] [Green Version]
- Julián, E.; Cama, M.; Martínez, P.; Luquin, M. An ELISA for five glycolipids from the cell wall of Mycobacterium tuberculosis: Tween 20 interference in the assay. J. Immunol. Methods 2001, 251, 21–30. [Google Scholar] [CrossRef]
- Stoops, J.K.; Arora, R.; Armitage, L.; Wanger, A.; Song, L.; Blackburn, M.R.; Krueger, G.R.; Risin, S.A. Certain surfactants show promise in the therapy of pulmonary tuberculosis. In Vivo 2010, 24, 687–694. [Google Scholar]
- Setterington, E.B.; Cloutier, B.C.; Ochoa, J.M.; Cloutier, A.K.; Patel, P.J.; Alocilja, E.C. Rapid, sensitive, and specific immunomagnetic separation of foodborne pathogens. Int. J. Food Saf. Nutr. Public Health 2011, 4, 83–100. [Google Scholar] [CrossRef]
- Laue, M. Negative Staining Electron Microscopy Tips & Trick for the Diagnosis of Pathogens. Available online: https://analyticalscience.wiley.com/do/10.1002/imaging.5585 (accessed on 19 September 2020).
- OPS. Manual Para el Diagnóstico Bacteriológico de La Tuberulosis. Normas y Guía Técnica. Parte II Cultivo; Washington, DC, USA, 2008; pp. 1–114. Available online: https://iris.paho.org/handle/10665.2/18616 (accessed on 19 September 2020).
- NATA. Guidelines for the Validation and Verification of Quantitative and Qualitative Test Methods. Available online: http://www.demarcheiso17025.com/document/Guidelines for the validation and verification of quantitative and qualitative test methods.pdf (accessed on 13 July 2018).
- Wayne, D.W. Analisis de varianza. In Bioestadística: Base para El Análisis de Las Ciencias de La Salud; Editorial Limusa S.A. de C.V.: Mexico City, Mexico, 2005; pp. 295–399. ISBN 978-9681861643. [Google Scholar]
- Wayne, D.W. Estadística no paramétrica y de libre distribución. In Bioestadistica: Base para El Analisis de Las Ciencias de La Salud; Editorial Limusa S.A. de C.V.: Mexico City, Mexico, 2005; pp. 658–736. ISBN 978-9681861643. [Google Scholar]
- Kvítek, L.; Panáček, A.; Soukupová, J.; Kolář, M.; Večeřová, R.; Prucek, R.; Holecová, M.; Zbořil, R. Effect of surfactants and polymers on stability and antibacterial activity of silver nanoparticles (NPs). J. Phys. Chem. C 2008, 112, 5825–5834. [Google Scholar] [CrossRef]
- Saleh, N.; Kim, H.J.; Phenrat, T.; Matyjaszewski, K.; Tilton, R.D.; Lowry, G.V. Ionic strength and composition affect the mobility of surface-modified Fe0 nanoparticles in water-saturated sand columns. Environ. Sci. Technol. 2008, 42, 3349–3355. [Google Scholar] [CrossRef] [PubMed]
- Toutain-Kidd, C.M.; Kadivar, S.C.; Bramante, C.T.; Bobin, S.A.; Zegans, M.E. Polysorbate 80 inhibition of Pseudomonas aeruginosa biofilm formation and its cleavage by the secreted lipase LipA. Antimicrob. Agents Chemother. 2009, 53, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, M.T.; Huynh, S. Effect of the surfactant Tween 80 on the detachment and dispersal of Salmonella enterica serovar Thompson single cells and aggregates from Cilantro leaves as revealed by image analysis. Appl. Environ. Microbiol. 2014, 80, 5037–5042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, Y.; Mashima, K.; Miyazaki, M.; Hara, S.; Takata, T.; Kamimura, H.; Takagi, S.; Jimi, S. Inhibitory effects of polysorbate 80 on MRSA biofilm formed on different substrates including dermal tissue. Sci. Rep. 2019, 9, 3128. [Google Scholar] [CrossRef]
- Phenrat, T.; Kim, H.J.; Fagerlund, F.; Illangasekare, T.; Tilton, R.D.; Lowry, G.V. Particle size distribution, concentration, and magnetic attraction affect transport of polymer-modified Fe0 nanoparticles in sand columns. Environ. Sci. Technol. 2009, 43, 5079–5085. [Google Scholar] [CrossRef]
- Leisching, G.; Pietersen, R.-D.; Wiid, I.; Baker, B. Virulence, biochemistry, morphology and host-interacting properties of detergent-free cultured mycobacteria: An update. Tuberculosis 2016, 100, 53–60. [Google Scholar] [CrossRef]
- Masaki, S.; Sugimori, G.; Okamoto, A.; Imose, J.; Hayashi, Y. Effect of Tween 80 on the Growth of Mycobacterium avium Complex. Microbiol. Immunol. 1990, 34, 653–663. [Google Scholar] [CrossRef]
- Masaki, S.; Sugimori, G.; Okamoto, A.; Imose, J.; Hayashi, Y. Effect of Tween 80 on formation of the superficial L1 layer of the Mycobacterium avium-Mycobacterium intracellulare complex. J. Clin. Microbiol. 1991, 29, 1453–1456. [Google Scholar] [CrossRef] [Green Version]
- Domenech, P.; Reed, M.B. Rapid and spontaneous loss of phthiocerol dimycocerosate (PDIM) from Mycobacterium tuberculosis grown in vitro: Implications for virulence studies. Microbiology 2009, 155, 3532–3543. [Google Scholar] [CrossRef] [Green Version]
- Hunter, R.L.; Olsen, M.R.; Jagannath, C.; Actor, J.K. Multiple roles of cord factor in the pathogenesis of primary, secondary, and cavitary tuberculosis, including a revised description of the pathology of secondary disease. Ann. Clin. Lab. Sci. 2006, 36, 371–386. [Google Scholar]
- Bowdish, D.M.E.; Sakamoto, K.; Kim, M.J.; Kroos, M.; Mukhopadhyay, S.; Leifer, C.A.; Tryggvason, K.; Gordon, S.; Russell, D.G. MARCO, TLR2, and CD14 are required for macrophage cytokine responses to mycobacterial trehalose dimycolate and Mycobacterium tuberculosis. PLoS Pathog. 2009, 5, e1000474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrop, J.; Jozefowski, K.; Zimmermann, S.; Hofmann, K.; Strasser, E.; Lepenies, B.; Lang, R. Contribution of MINCLE–SYK signaling to activation of primary human APCs by Mycobacterial Cord Factor and the novel adjuvant TDB. J. Immunol. 2015, 195, 2417–2428. [Google Scholar] [CrossRef]
- Yu, J.; Tran, V.; Li, M.; Huang, X.; Niu, C.; Wang, D.; Zhu, J.; Wang, J.; Gao, Q.; Liu, J. Both phthiocerol dimycocerosates and phenolic glycolipids are required for virulence of Mycobacterium marinum. Infect. Immun. 2012, 80, 1381–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, C.; Winter, N.; Privert, E.; Bordat, Y.; Neyrolles, O.; Avé, P.; Huerre, M.; Gicquel, B.; Jackson, M. Production of phthiocerol dimycocerosates protects Mycobacterium tuberculosis from the cidal activity of reactive nitrogen intermediates produced by macrophages and modulates the early immune response to infection. Cell. Microbiol. 2004, 6, 277–287. [Google Scholar] [CrossRef]
- Mekonen, A.; Ayele, Y.; Berhan, Y.; Woldeyohannes, D.; Erku, W.; Sisay, S. Factors which contributed for low quality sputum smears for the detection of acid fast bacilli (AFB) at selected health centers in Ethiopia: A quality control perspective. PLoS ONE 2018, 13, e0198947. [Google Scholar] [CrossRef]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat. Rev. Dis. Prim. 2016, 2, 16076. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, I.N.; Figueredo, L.J.d.A.; Soares, V.M.; Vater, M.C.; Alves, S.; Carvalho, W.d.S.; Kritski, A.L.; de Miranda, S.S. Evaluation of the mean cost and activity based cost in the diagnosis of pulmonary tuberculosis in the laboratory routine of a high-complexity hospital in Brazil. Front. Microbiol. 2017, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Kaso, A.; Hailu, A. Cost-effectiveness of GeneXpert Compared to Smear Microscopy for Diagnosis of Tuberculosis in Ethiopia. Res. Sq. 2020, 1–20, pre-print. [Google Scholar] [CrossRef]
- Sánchez-Pérez, H.J.; Bernal, D.; Torres, O.; Colorado, A. Tuberculosis en México en tiempos de COVID-19: Algunas reflexiones Introducción. Enferm. Emerg. 2021, 20, 160–165. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. AFB Observed | AFB Quantification | Estimated AFB Concentration/mL |
|---|---|---|
| 0 in 100 or more fields | Negative | 0 |
| 1–9 AFB/100 fields | Scanty | 30,000 (3 × 104) |
| 10–99 AFB/100 fields | 1+ | 50,000 (5 × 104) |
| 1–10 AFB/field in at least 50 fields | 2+ | 100,000 (1 × 105) |
| >10 AFB/field in at least 20 fields | 3+ | 500,000 (5 × 105) |
| Method | GMNP (mg/mL) | No. Smears | No. Fields Read a | Median (AFB) | IQR |
|---|---|---|---|---|---|
| SSM | - | 30 | 3000 | 9 | 1−44.8 |
| NCBA | 0.25 | 12 | 1200 | 22 | 14−78 |
| 0.5 | 12 | 1200 | 28 | 4−112 | |
| 0.75 | 10 | 1000 | 40 | 2−121.5 | |
| 1.0 | 10 | 1000 | 48 | 14−142 | |
| 1.25 | 11 | 1100 | 10 | 2-80 | |
| 1.5 | 7 | 700 | 34 | 4−99.5 | |
| 1.75 | 6 | 600 | 6 | 0−52 | |
| 2.0 | 6 | 600 | 4 | 0−38 | |
| 2.25 | 3 | 300 | 0 | 0−6.75 | |
| 2.5 | 10 | 1000 | 4 | 0−51 | |
| 10.0 | 10 | 2000 | 0 | 0−42 | |
| Total | 137 | 13,700 | - | - |
| GMNP (mg/mL) | No. Fields/ Smears | Fields SSM Negative Improved with NCBA to S+, 1+, 2+ and 3+ | Fields SSM S+ Improved with NCBA to 1+ and 2+ | Fields SSM 1+ Improved with NCBA to 2+ and 3+ | |||
|---|---|---|---|---|---|---|---|
| Fields * | % | Fields * | % | Fields * | % | ||
| 0.25 | 1200 | 65/127 | 51.2 | 117/306 | 38.2 | 89/457 | 19.5 |
| 0.5 | 1200 | 39/127 | 30.7 | 98/306 | 32 | 91/457 | 20 |
| 0.75 | 1000 | 32/126 | 25.4 | 39/250 | 16 | 67/314 | 21.3 |
| 1.0 | 1000 | 47/72 | 65.3 | 76/224 | 34 | 44/394 | 11.2 |
| 1.25 | 1100 | 102/202 | 50.5 | 111/437 | 25.4 | 83/306 | 27.1 |
| 1.5 | 700 | 33/72 | 45.8 | 56/224 | 25 | 62/249 | 25 |
| 1.75 | 600 | 31/72 | 43.1 | 30/206 | 14.6 | 3/167 | 1.8 |
| 2.0 | 600 | 19/72 | 26.4 | 30/206 | 14.6 | 9/167 | 5.4 |
| 2.25 | 300 | 15/67 | 22.4 | 8/124 | 6.5 | 2/56 | 3.6 |
| 2.5 | 1000 | 89/202 | 44.1 | 67/419 | 16 | 16/224 | 7.1 |
| 10.0 | 2000 | 76/546 | 13.9 | 41/566 | 7.2 | 64/604 | 10.6 |
| Total, ≤1.5 | 6200 | 318/726 | 43.8 | 497/1747 | 28.4 | 436/2177 | 20.0 |
| Total, 0.25−10 | 10,700 | 548/1685 | 32.5 | 673/3268 | 20.6 | 530/3395 | 15.6 |
| GMNP (mg/mL) | Mean of AFB in NCBA (Number of Fields) * | SD | ||||||
|---|---|---|---|---|---|---|---|---|
| SSM- | SSM <9 | SSM 1+ | SSM 2+ and 3+ | SSM- | SSM <9 | SSM 1+ | SSM 2+ and 3+ | |
| 0.25 | 7 (65) | 16 (193) | 79 (422) | 188 (306) | 24 | 30 | 134 | 209 |
| 0.5 | 3 (39) | 15 (187) | 56 (424) | 558 (310) | 9 | 33 | 56 | 540 |
| 0.75 | 2 (32) | 5 (128) | 70 (306) | 486 (310) | 4 | 10 | 54 | 734 |
| 1.0 | 7 (47) | 11 (163) | 59 (391) | 675 (310) | 11 | 14 | 43 | 846 |
| 1.25 | 6 (102) | 9 (294) | 69 (293) | 2030 (155) | 12 | 15 | 49 | 2097 |
| 1.5 | 2 (33) | 9 (155) | 73 (244) | 1740 (155) | 3 | 16 | 46 | 1580 |
| 1.75 | 3 (31) | 5 (94) | 34 (141) | 269 (149) | 5 | 9 | 33 | 231 |
| 2.0 | 3 (19) | 5 (99) | 22 (105) | 1206 (155) | 11 | 10 | 41 | 1186 |
| 2.25 | 1 (15) | 2 (36) | 19 (48) | 15 (45) | 2 | 6 | 19 | 19 |
| 2.5 | 4 (89) | 5 (184) | 44 (191) | 1891 (155) | 11 | 10 | 41 | 1605 |
| 10.0 | 1 (76) | 3 (136) | 44 (445) | 119 (278) | 3 | 13 | 43 | 136 |
| GMNP (mg/mL) | [Tween 80] | AFB Number (Mean) | No. Fields a | Percentage Difference between TW with Respect NCBA and SSM | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TW * | TW ** | NCBA | SSM | TW *-NCBA | TW *-SSM | TW **-NCBA | TW **-SSM | |||
| 1.25 | 5% | 2400 | 956 | 804 | 441 | 3200 | 199 | 445 | 19 | 117 |
| 10% | 1915 | 914 | 138 | 335 | 14 | 107 | ||||
| 15% | 1310 | 991 | 63 | 197 | 23 | 125 | ||||
| 20% | 1335 | 825 | 41 | 158 | 3 | 87 | ||||
| 2.5 | 5% | 937 | 694 | 532 | 76 | 113 | 30 | 58 | ||
| 10% | 936 | 568 | 76 | 112 | 7 | 29 | ||||
| 15% | 937 | 601 | 76 | 113 | 13 | 36 | ||||
| 20% | 727 | 662 | 37 | 65 | 24 | 50 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordillo-Marroquín, C.; Sánchez-Pérez, H.J.; Gómez-Velasco, A.; Martín, M.; Guillén-Navarro, K.; Vázquez-Marcelín, J.; Gómez-Bustamante, A.; Jonapá-Gómez, L.; Alocilja, E.C. Tween 80 Improves the Acid-Fast Bacilli Quantification in the Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA). Biosensors 2022, 12, 29. https://doi.org/10.3390/bios12010029
Gordillo-Marroquín C, Sánchez-Pérez HJ, Gómez-Velasco A, Martín M, Guillén-Navarro K, Vázquez-Marcelín J, Gómez-Bustamante A, Jonapá-Gómez L, Alocilja EC. Tween 80 Improves the Acid-Fast Bacilli Quantification in the Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA). Biosensors. 2022; 12(1):29. https://doi.org/10.3390/bios12010029
Chicago/Turabian StyleGordillo-Marroquín, Cristina, Héctor J. Sánchez-Pérez, Anaximandro Gómez-Velasco, Miguel Martín, Karina Guillén-Navarro, Janeth Vázquez-Marcelín, Adriana Gómez-Bustamante, Letisia Jonapá-Gómez, and Evangelyn C. Alocilja. 2022. "Tween 80 Improves the Acid-Fast Bacilli Quantification in the Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA)" Biosensors 12, no. 1: 29. https://doi.org/10.3390/bios12010029
APA StyleGordillo-Marroquín, C., Sánchez-Pérez, H. J., Gómez-Velasco, A., Martín, M., Guillén-Navarro, K., Vázquez-Marcelín, J., Gómez-Bustamante, A., Jonapá-Gómez, L., & Alocilja, E. C. (2022). Tween 80 Improves the Acid-Fast Bacilli Quantification in the Magnetic Nanoparticle-Based Colorimetric Biosensing Assay (NCBA). Biosensors, 12(1), 29. https://doi.org/10.3390/bios12010029