
High-Dose Exposure to Polymer-Coated Iron Oxide Nanoparticles Elicits Autophagy-Dependent Ferroptosis in Susceptible Cancer Cells
 ,
,  , ,
, ,  , , , , and
, , , , and 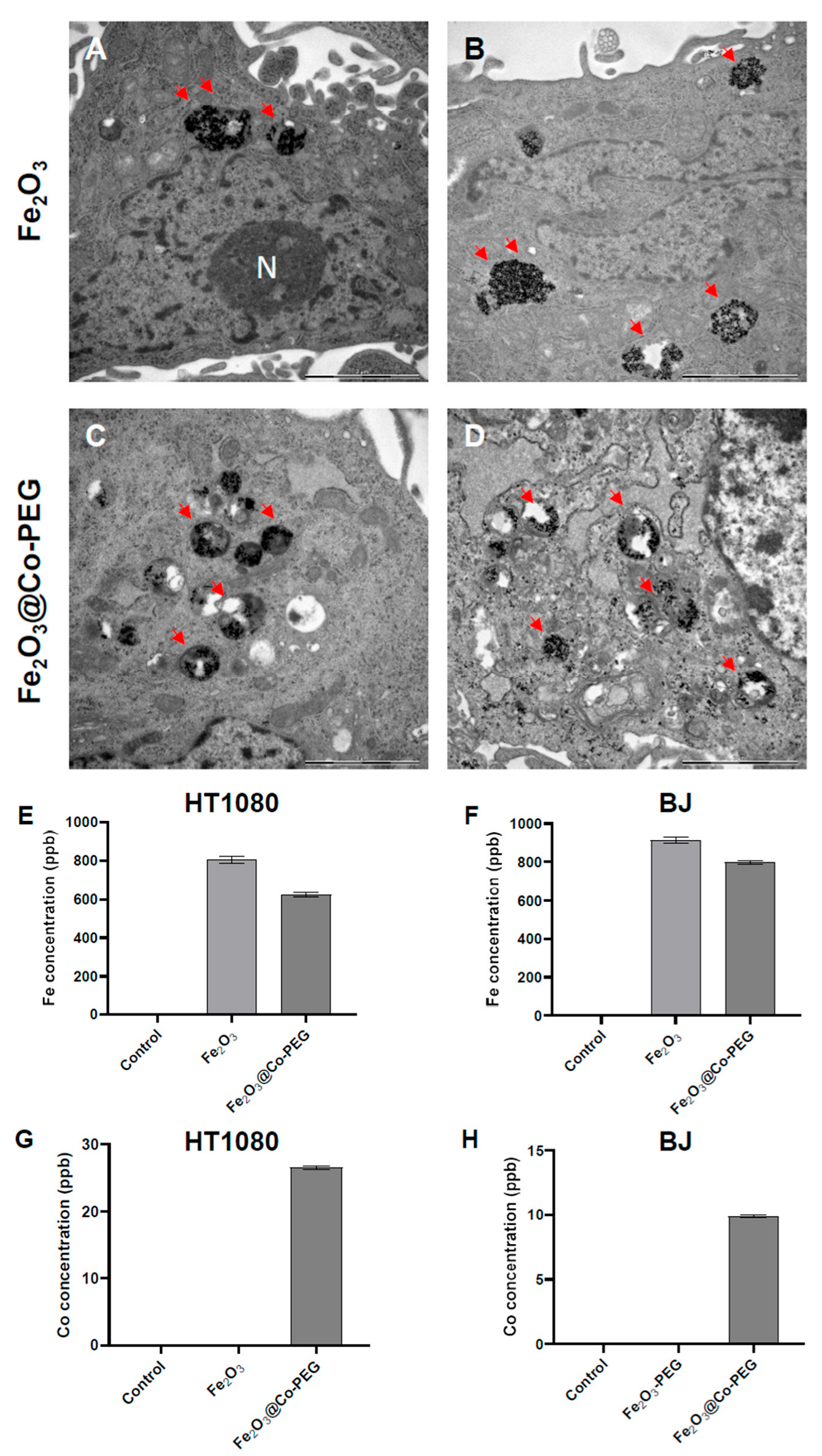{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Nanoparticles
2.2. Human Cell Lines
2.3. TEM Analysis
2.4. ICP-MS Analysis
2.5. LDH Release Assay
2.6. Metabolic Activity Assay
2.7. Lipid Peroxidation Assay
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Cellular Internalization of Polymer-Coated Fe2O3 Nanoparticles
3.2. Toxicity of Fe2O3 Nanoparticles with/without Cobalt Functionalization
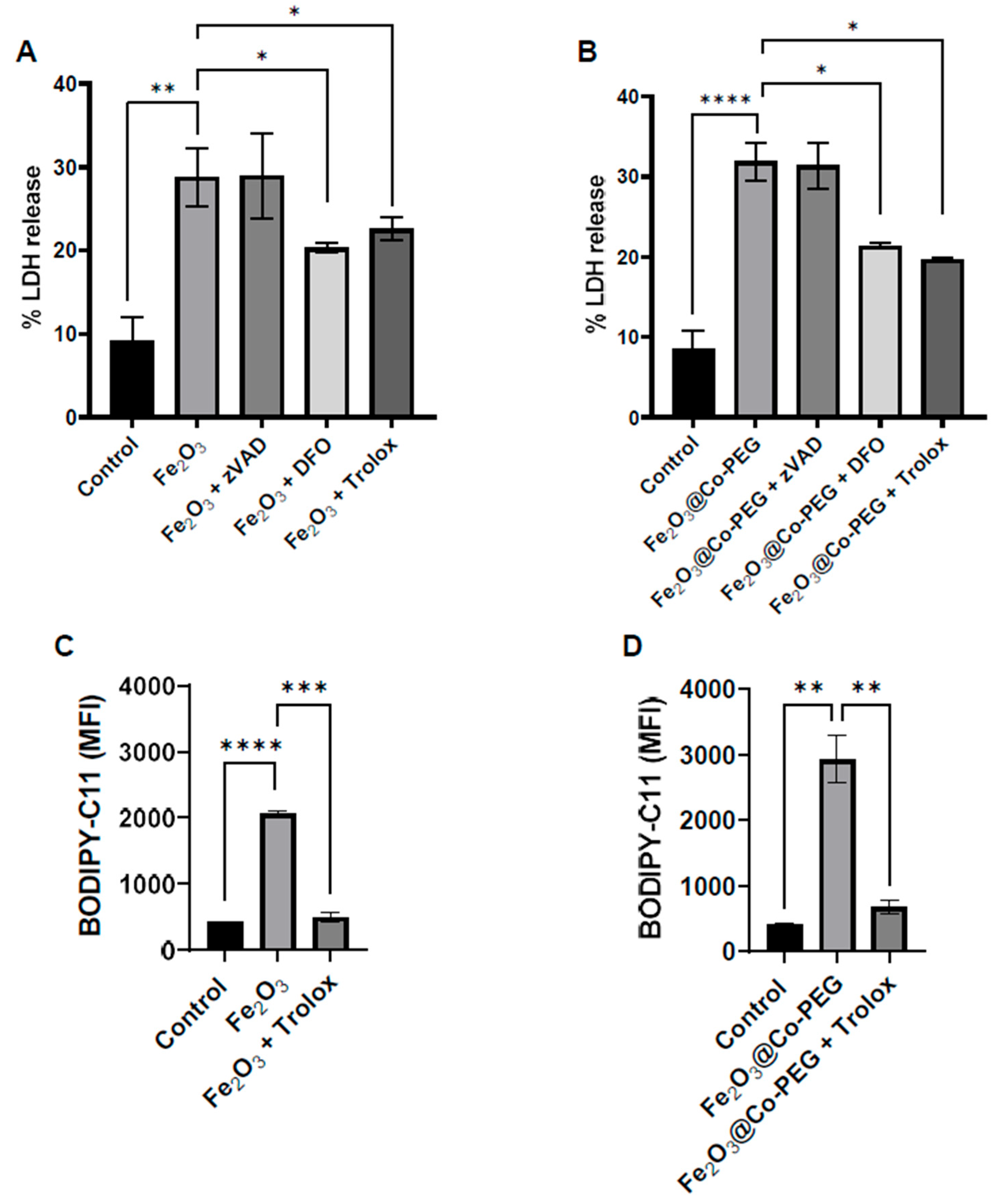
3.3. High-Dose Exposure to Fe2O3 Nanoparticles Triggers Ferroptosis
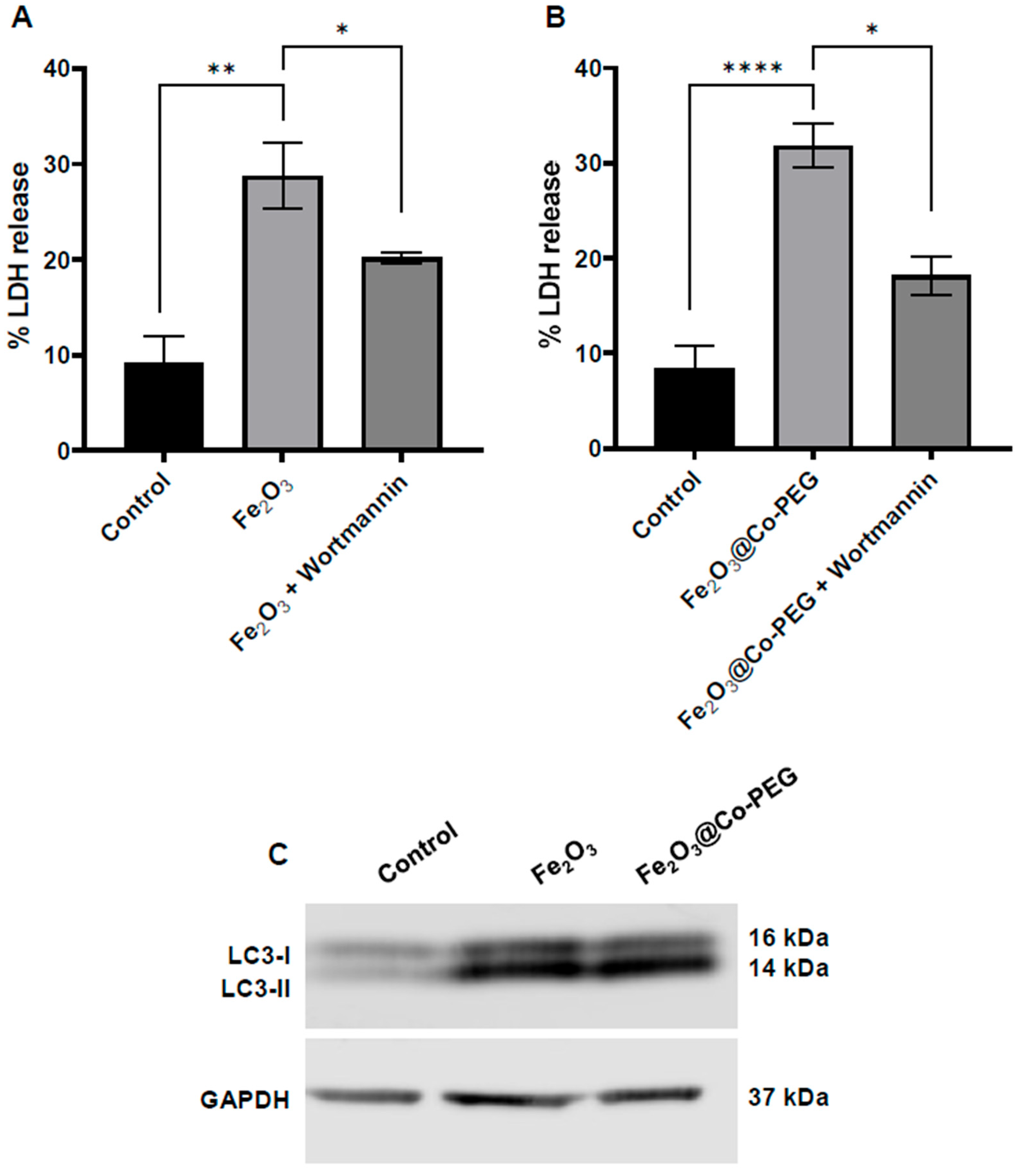
3.4. Fe2O3 Nanoparticle-Triggered Ferroptosis Is Autophagy-Dependent
3.5. No Cytotoxicity Elicited by PEG-PLGA-Modified Fe3O4 Nanoparticles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knüchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [Google Scholar] [CrossRef]
- Fadeel, B.; Garcia-Bennett, A.E. Better safe than sorry: Understanding the toxicological properties of inorganic nanoparticles manufactured for biomedical applications. Adv. Drug Deliv. Rev. 2010, 62, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Zhuang, L.; Gan, B. Targeting ferroptosis as a vulnerability in cancer. Nat. Rev. Cancer. 2022, 22, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Lomphithak, T.; Fadeel, B. Die hard: Cell death mechanisms and their implications in nanotoxicology. Toxicol. Sci. 2023, 192, 141–154. [Google Scholar] [CrossRef]
- Fernández-Acosta, R.; Iriarte-Mesa, C.; Alvarez-Alminaque, D.; Hassannia, B.; Wiernicki, B.; Díaz-García, A.M.; Vandenabeele, P.; Vanden Berghe, T.; Pardo Andreu, G.L. Novel iron oxide nanoparticles induce ferroptosis in a panel of cancer cell lines. Molecules 2022, 27, 3970. [Google Scholar] [CrossRef]
- Bae, C.; Kim, H.; Kook, Y.M.; Lee, C.; Kim, C.; Yang, C.; Park, M.H.; Piao, Y.; Koh, W.G.; Lee, K. Induction of ferroptosis using functionalized iron-based nanoparticles for anti-cancer therapy. Mater. Today Bio 2022, 17, 100457. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, H.; Zhao, Y.; Lu, L.; Zhang, J.; Cheng, W.; Song, X.; Zheng, Y.; Chen, C.; Tang, J. Time-course effect of ultrasmall superparamagnetic iron oxide nanoparticles on intracellular iron metabolism and ferroptosis activation. Nanotoxicology 2021, 15, 366–379. [Google Scholar] [CrossRef]
- Wen, J.; Chen, H.; Ren, Z.; Zhang, P.; Chen, J.; Jiang, S. Ultrasmall iron oxide nanoparticles induced ferroptosis via Beclin1/ATG5-dependent autophagy pathway. Nano Converg. 2021, 8, 10. [Google Scholar] [CrossRef]
- Zhang, X.; Kong, F.; Wang, T.; Huang, X.; Li, W.; Zhang, M.; Wen, T.; Liu, J.; Zhang, Y.; Meng, J.; et al. Iron oxide nanoparticles cause surface coating- and core chemistry-dependent endothelial cell ferroptosis. Nanotoxicology 2023, 16, 829–843. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Khan, M.A.M.; Alhadlaq, H.A.; Alshamsan, A. Cobalt iron oxide nanoparticles induce cytotoxicity and regulate the apoptotic genes through ROS in human livercells (HepG2). Colloids Surf. B Biointerfaces 2016, 148, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Abudayyak, M.; Altincekic Gurkaynak, T.; Özhan, G. In vitro toxicological assessment of cobalt ferrite nanoparticles in several mammalian cell types. Biol. Trace Elem. Res. 2017, 175, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Dutz, S.; Buske, N.; Landers, J.; Gräfe, C.; Wende, H.; Clement, J.H. Biocompatible magnetic fluids of Co-doped iron oxide nanoparticles with tunable magnetic properties. Nanomaterials 2020, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, J.; Turiel, D.; Bettmer, J.; Jakubowski, N.; Panne, U.; Rivas García, L.; Llopis, J.; Sánchez González, C.; Montes-Bayón, M. In vitro and in situ experiments to evaluate the biodistribution and cellular toxicity of ultrasmall iron oxide nanoparticles potentially used as oral iron supplements. Nanotoxicology 2020, 14, 388–403. [Google Scholar] [CrossRef] [PubMed]
- Ovejero, J.G.; Armenia, I.; Serantes, D.; Veintemillas-Verdaguer, S.; Zeballos, N.; López-Gallego, F.; Grüttner, C.; de la Fuente, J.M.; Puerto Morales, M.D.; Grazú, V. Selective magnetic nanoheating: Combining iron oxide nanoparticles for multi-hot-spot induction and sequential regulation. Nano Lett. 2021, 21, 7213–7220. [Google Scholar] [CrossRef]
- D’Elios, M.M.; Aldinucci, A.; Amoriello, R.; Benagiano, M.; Bonechi, E.; Maggi, P.; Flori, A.; Ravagli, C.; Saer, D.; Cappiello, L.; et al. Myelin-specific T cells carry and release magnetite PGLA-PEG COOH nanoparticles in the mouse central nervous system. RSC Adv. 2018, 8, 904–913. [Google Scholar] [CrossRef]
- Psimadas, D.; Baldi, G.; Ravagli, C.; Comes Franchini, M.; Locatelli, E.; Innocenti, C.; Sangregorio, C.; Loudos, G. Comparison of the magnetic, radiolabeling, hyperthermic and biodistribution properties of hybrid nanoparticles bearing CoFe2O4 and Fe3O4 metal cores. Nanotechnology 2014, 25, 025101. [Google Scholar] [CrossRef]
- Martin, S.; de Haan, L.; Miro Estruch, I.; Eder, K.M.; Marzi, A.; Schnekenburger, J.; Blosi, M.; Costa, A.; Antonello, G.; Bergamaschi, E.; et al. Pre-validation of a reporter gene assay for oxidative stress for the rapid screening of nanobiomaterials. Front. Toxicol. 2022, 4, 974429. [Google Scholar] [CrossRef]
- Keshavan, S.; Gupta, G.; Martin, S.; Fadeel, B. Multi-walled carbon nanotubes trigger lysosome-dependent cell death (pyroptosis) in macrophages but not in neutrophils. Nanotoxicology 2021, 15, 1125–1150. [Google Scholar] [CrossRef]
- Gupta, G.; Gliga, A.; Hedberg, J.; Serra, A.; Greco, D.; Odnevall Wallinder, I.; Fadeel, B. Cobalt nanoparticles trigger ferroptosis-like cell death (oxytosis) in neuronal cells: Potential implications for neurodegenerative disease. FASEB J. 2020, 34, 5262–5281. [Google Scholar] [CrossRef]
- Torii, S.; Shintoku, R.; Kubota, C.; Yaegashi, M.; Torii, R.; Sasaki, M.; Suzuki, T.; Mori, M.; Yoshimoto, Y.; Takeuchi, T.; et al. An essential role for functional lysosomes in ferroptosis of cancer cells. Biochem. J. 2016, 473, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Shawki, A.; Ganz, T.; Nemeth, E.; Mackenzie, B. Functional properties of human ferroportin, a cellular iron exporter reactive also with cobalt and zinc. Am. J. Physiol. Physiol. 2014, 306, C450–C459. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, T.; Linkermann, A.; Jouan-Lanhouet, S.; Walczak, H.; Vandenabeele, P. Regulated necrosis: The expanding network of non-apoptotic cell death pathways. Nat. Rev. Mol. Cell. Biol. 2014, 15, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis is an autophagic cell death process. Cell Res. 2016, 26, 1021–1032. [Google Scholar] [CrossRef]
- Marassi, V.; Zanoni, I.; Ortelli, S.; Giordani, S.; Reschiglian, P.; Roda, B.; Zattoni, A.; Ravagli, C.; Cappiello, L.; Baldi, G.; et al. Native study of the behaviour of magnetite nanoparticles for hyperthermia treatment during the initial moments of intravenous administration. Pharmaceutics 2022, 14, 2810. [Google Scholar] [CrossRef]
- Keshavan, S.; Andón, F.T.; Gallud, A.; Chen, W.; Reinert, K.; Tran, L.; Fadeel, B. Profiling of sub-lethal in vitro effects of multi-walled carbon nanotubes reveals changes in chemokines and chemokine receptors. Nanomaterials 2021, 11, 883. [Google Scholar] [CrossRef]
- Basit, F.; van Oppen, L.M.; Schöckel, L.; Bossenbroek, H.M.; van Emst-de Vries, S.E.; Hermeling, J.C.; Grefte, S.; Kopitz, C.; Heroult, M.; Hgm Willems, P.; et al. Mitochondrial complex I inhibition triggers a mitophagy-dependent ROS increase leading to necroptosis and ferroptosis in melanoma cells. Cell Death Dis. 2017, 8, e2716. [Google Scholar] [CrossRef]
- Dos Santos, A.F.; Inague, A.; Arini, G.S.; Terra, L.F.; Wailemann, R.A.M.; Pimentel, A.C.; Yoshinaga, M.Y.; Silva, R.R.; Severino, D.; de Almeida, D.R.Q.; et al. Distinct photo-oxidation-induced cell death pathways lead to selective killing of human breast cancer cells. Cell Death Dis. 2020, 11, 1070. [Google Scholar] [CrossRef]
- Gong, Y.N.; Guy, C.; Crawford, J.C.; Green, D.R. Biological events and molecular signaling following MLKL activation during necroptosis. Cell Cycle 2017, 16, 1748–1760. [Google Scholar] [CrossRef]
- Wegner, S.V.; Spatz, J.P. Cobalt(III) as a stable and inert mediator ion between NTA and His6-tagged proteins. Angew. Chem. Int. Ed. 2013, 52, 7593–7596. [Google Scholar] [CrossRef]
- Líbalová, H.; Costa, P.M.; Olsson, M.; Farcal, L.; Ortelli, S.; Blosi, M.; Topinka, J.; Costa, A.L.; Fadeel, B. Toxicity of surface-modified copper oxide nanoparticles in a mouse macrophage cell line: Interplay of particles, surface coating and particle dissolution. Chemosphere 2018, 196, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.M.; Cheung, P.C.K. Gallic acid triggers iron-dependent cell death with apoptotic, ferroptotic, and necroptotic features. Toxins 2019, 11, 492. [Google Scholar] [CrossRef] [PubMed]
- Maurizi, L.; Papa, A.L.; Dumont, L.; Bouyer, F.; Walker, P.; Vandroux, D.; Millot, N. Influence of surface charge and polymer coating on internalization and biodistribution of polyethylene glycol-modified iron oxide nanoparticles. J. Biomed. Nanotechnol. 2015, 11, 126–136. [Google Scholar] [CrossRef]
- Patsula, V.; Tulinska, J.; Trachtová, S.; Kuricova, M.; Liskova, A.; Spanová, A.; Ciampor, F.; Vavra, I.; Rittich, B.; Ursinyova, M.; et al. Toxicity evaluation of monodisperse PEGylated magnetic nanoparticles for nanomedicine. Nanotoxicology 2019, 13, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Sun, X.; Shi, Y.; Chen, L.; Wang, L.; Cai, H.; Han, C.; Liao, T.; Yang, C.; Zuo, Z.; et al. The valence state of iron-based nanomaterials determines the ferroptosis potential in a zebrafish model. Sci. Total Environ. 2023, 855, 158715. [Google Scholar] [CrossRef] [PubMed]
- Stoyanovsky, D.A.; Tyurina, Y.Y.; Shrivastava, I.; Bahar, I.; Tyurin, V.A.; Protchenko, O.; Jadhav, S.; Bolevich, S.B.; Kozlov, A.V.; Vladimirov, Y.A.; et al. Iron catalysis of lipid peroxidation in ferroptosis: Regulated enzymatic or random free radical reaction? Free Radic. Biol. Med. 2019, 133, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Crielaard, B.J.; Lammers, T.; Rivella, S. Targeting iron metabolism in drug discovery and delivery. Nat. Rev. Drug Discov. 2017, 16, 400–423. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem. Biol. 2008, 15, 234–245. [Google Scholar] [CrossRef]
- Khan, M.I.; Mohammad, A.; Patil, G.; Naqvi, S.A.; Chauhan, L.K.; Ahmad, I. Induction of ROS, mitochondrial damage and autophagy in lung epithelial cancer cells by iron oxide nanoparticles. Biomaterials 2012, 33, 1477–1488. [Google Scholar] [CrossRef]
- Panja, P.; Debnath, K.; Jana, N.R.; Jana, N.R. Surface chemistry- and intracellular trafficking-dependent autophagy induction by iron oxide nanoparticles. ACS Appl. Bio Mater. 2020, 3, 5974–5983. [Google Scholar] [CrossRef]
- Portilla, Y.; Mulens-Arias, V.; Paradela, A.; Ramos-Fernández, A.; Pérez-Yagüe, S.; Morales, M.P.; Barber, D.F. The surface coating of iron oxide nanoparticles drives their intracellular trafficking and degradation in endolysosomes differently depending on the cell type. Biomaterials 2022, 281, 121365. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Xie, Y.; Song, X.; Sun, X.; Lotze, M.T.; Zeh, H.J.; Kang, R.; Tang, D. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy 2016, 12, 1425–1428. [Google Scholar] [CrossRef] [PubMed]
- Mancias, J.D.; Wang, X.; Gygi, S.P.; Harper, J.W.; Kimmelman, A.C. Quantitative proteomics identifies NCOA4 as the cargo receptor mediating ferritinophagy. Nature 2014, 509, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhang, J.; Wang, B.; Xu, G.; Yang, X.; Zou, Z.; Yu, C. Ferritinophagy is involved in the zinc oxide nanoparticles-induced ferroptosis of vascular endothelial cells. Autophagy 2021, 17, 4266–4285. [Google Scholar] [CrossRef] [PubMed]
- Frtús, A.; Smolková, B.; Uzhytchak, M.; Lunova, M.; Jirsa, M.; Kubinová, S.; Dejneka, A.; Lunov, O. Analyzing the mechanisms of iron oxide nanoparticles interactions with cells: A road from failure to success in clinical applications. J. Control. Release 2020, 328, 59–77. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lomphithak, T.; Helvacioglu, S.; Armenia, I.; Keshavan, S.; Ovejero, J.G.; Baldi, G.; Ravagli, C.; Grazú, V.; Fadeel, B. High-Dose Exposure to Polymer-Coated Iron Oxide Nanoparticles Elicits Autophagy-Dependent Ferroptosis in Susceptible Cancer Cells. Nanomaterials 2023, 13, 1719. https://doi.org/10.3390/nano13111719
Lomphithak T, Helvacioglu S, Armenia I, Keshavan S, Ovejero JG, Baldi G, Ravagli C, Grazú V, Fadeel B. High-Dose Exposure to Polymer-Coated Iron Oxide Nanoparticles Elicits Autophagy-Dependent Ferroptosis in Susceptible Cancer Cells. Nanomaterials. 2023; 13(11):1719. https://doi.org/10.3390/nano13111719
Chicago/Turabian StyleLomphithak, Thanpisit, Selin Helvacioglu, Ilaria Armenia, Sandeep Keshavan, Jesús G. Ovejero, Giovanni Baldi, Costanza Ravagli, Valeria Grazú, and Bengt Fadeel. 2023. "High-Dose Exposure to Polymer-Coated Iron Oxide Nanoparticles Elicits Autophagy-Dependent Ferroptosis in Susceptible Cancer Cells" Nanomaterials 13, no. 11: 1719. https://doi.org/10.3390/nano13111719
APA StyleLomphithak, T., Helvacioglu, S., Armenia, I., Keshavan, S., Ovejero, J. G., Baldi, G., Ravagli, C., Grazú, V., & Fadeel, B. (2023). High-Dose Exposure to Polymer-Coated Iron Oxide Nanoparticles Elicits Autophagy-Dependent Ferroptosis in Susceptible Cancer Cells. Nanomaterials, 13(11), 1719. https://doi.org/10.3390/nano13111719