

A Structural Analysis of Proteinaceous Nanotube Cavities and Their Applications in Nanotechnology


Abstract

1. Introduction
2. Naturally Occurring Nanotubes and Their Functions
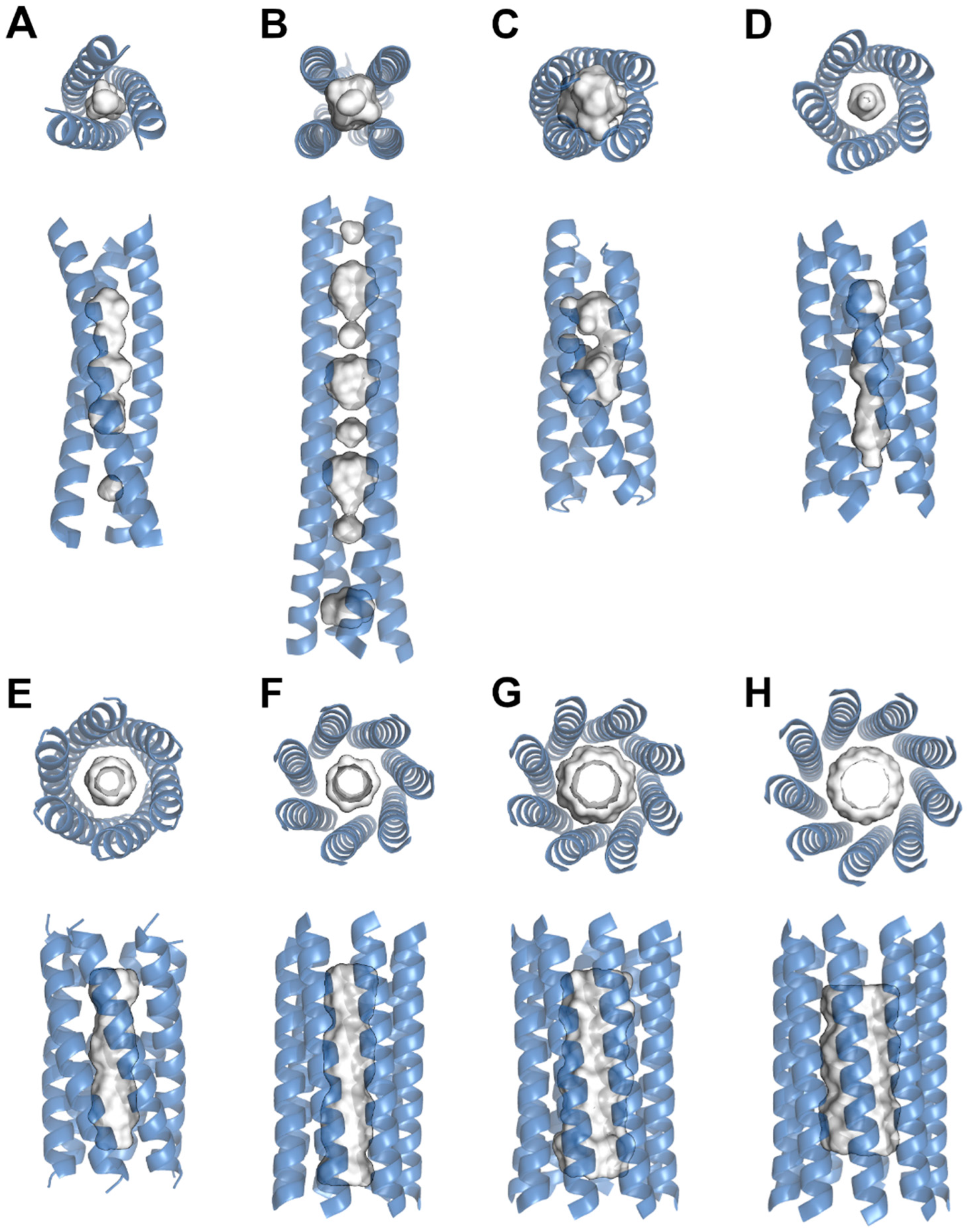
3. Structural Examination of Nanotube Cavities
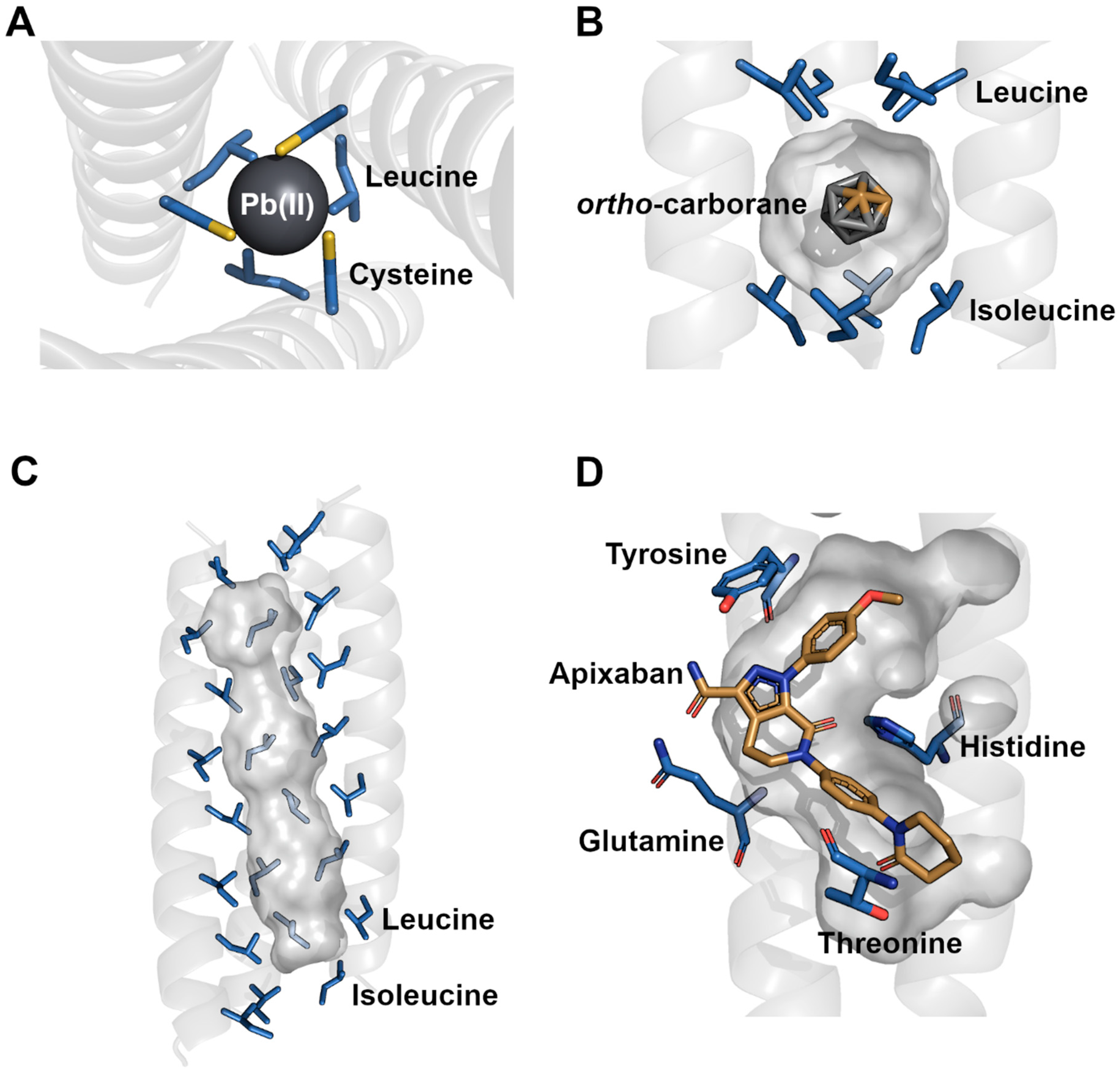
4. Nanotechnology Advancements in the Design of Nanotubes and Their Cavities
5. Applications for Nanotube Cavities
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chitranshi, M.; Pujari, A.; Ng, V.; Chen, D.; Chauhan, D.; Hudepohl, R.; Saleminik, M.; Kim, S.Y.; Kubley, A.; Shanov, V.; et al. Carbon Nanotube Sheet-Synthesis and Applications. Nanomaterials 2020, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Audette, G.F.; Yaseen, A.; Bragagnolo, N.; Bawa, R. Protein Nanotubes: From Bionanotech towards Medical Applications. Biomedicines 2019, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Hossain, M.K. Classification and Properties of Nanoparticles; Elsevier Ltd.: Amsterdam, The Netherlands, 2022; ISBN 9780128242728. [Google Scholar]
- Bati, A.S.R.; Yu, L.; Batmunkh, M.; Shapter, J.G. Recent Advances in Applications of Sorted Single-Walled Carbon Nanotubes. Adv. Funct. Mater. 2019, 29, 1902273. [Google Scholar] [CrossRef]
- Katouzian, I.; Jafari, S.M. Protein Nanotubes as State-of-the-Art Nanocarriers: Synthesis Methods, Simulation and Applications. J. Control. Release 2019, 303, 302–318. [Google Scholar] [CrossRef]
- Ren, D.; Dalmau, M.; Randall, A.; Shindel, M.M.; Baldi, P.; Wang, S.W. Biomimetic Design of Protein Nanomaterials for Hydrophobic Molecular Transport. Adv. Funct. Mater. 2012, 22, 3170–3180. [Google Scholar] [CrossRef]
- Jurado, S.; Cano-Muñoz, M.; Morel, B.; Standoli, S.; Santarossa, E.; Moog, C.; Schmidt, S.; Laumond, G.; Cámara-Artigas, A.; Conejero-Lara, F. Structural and Thermodynamic Analysis of HIV-1 Fusion Inhibition Using Small Gp41 Mimetic Proteins. J. Mol. Biol. 2019, 431, 3091–3106. [Google Scholar] [CrossRef]
- Thanasupawat, T.; Bergen, H.; Hombach-Klonisch, S.; Krcek, J.; Ghavami, S.; Del Bigio, M.R.; Krawitz, S.; Stelmack, G.; Halayko, A.; McDougall, M.; et al. Platinum (IV) Coiled Coil Nanotubes Selectively Kill Human Glioblastoma Cells. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 913–925. [Google Scholar] [CrossRef]
- Heide, F.; McDougall, M.; Harder-Viddal, C.; Roshko, R.; Davidson, D.; Wu, J.; Aprosoff, C.; Moya-Torres, A.; Lin, F.; Stetefeld, J. Boron Rich Nanotube Drug Carrier System Is Suited for Boron Neutron Capture Therapy. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Thomas, F.; Dawson, W.M.; Lang, E.J.M.; Burton, A.J.; Bartlett, G.J.; Rhys, G.G.; Mulholland, A.J.; Woolfson, D.N. De Novo-Designed α-Helical Barrels as Receptors for Small Molecules. ACS Synth. Biol. 2018, 7, 1808–1816. [Google Scholar] [CrossRef]
- Boyle, A.L.; Rabe, M.; Crone, N.S.A.; Rhys, G.G.; Soler, N.; Voskamp, P.; Pannu, N.S.; Kros, A. Selective Coordination of Three Transition Metal Ions within a Coiled-Coil Peptide Scaffold. Chem. Sci. 2019, 10, 7456–7465. [Google Scholar] [CrossRef]
- Harder-Viddal, C.; McDougall, M.; Roshko, R.M.; Stetefeld, J. Energetics of Storage and Diffusion of Water and Cyclo-Octasulfur for a Nonpolar Cavity of RHCC Tetrabrachion by Molecular Dynamics Simulations. Comput. Struct. Biotechnol. J. 2019, 17, 675–683. [Google Scholar] [CrossRef]
- McDougall, M.; Francisco, O.; Harder-Viddal, C.; Roshko, R.; Meier, M.; Stetefeld, J. Archaea S-Layer Nanotube from a “Black Smoker” in Complex with Cyclo-Octasulfur (S8) Rings. Proteins Struct. Funct. Bioinform. 2017, 85, 2209–2216. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Y.; He, Y.; Zhang, B.; Zhao, L.; Tian, S.; Wang, Q.; Chen, S.; Li, Z.; Liang, S.; et al. Intestinal-Targeted Nanotubes-in-Microgels Composite Carriers for Capsaicin Delivery and Their Effect for Alleviation of Salmonella Induced Enteritis. Biomaterials 2022, 287, 121613. [Google Scholar] [CrossRef]
- Chang, R.; Liu, B.; Wang, Q.; Zhang, J.; Yuan, F.; Zhang, H.; Chen, S.; Liang, S.; Li, Y. The Encapsulation of Lycopene with α-Lactalbumin Nanotubes to Enhance Their Anti-Oxidant Activity, Viscosity and Colloidal Stability in Dairy Drink. Food Hydrocoll. 2022, 131, 107792. [Google Scholar] [CrossRef]
- Breitwieser, A.; Sleytr, U.B.; Pum, D. A New Method for Dispersing Pristine Carbon Nanotubes Using Regularly Arranged S-Layer Proteins. Nanomaterials 2021, 11, 1346. [Google Scholar] [CrossRef]
- Mizuno, T.; Hasegawa, C.; Tanabe, Y.; Hamajima, K.; Muto, T.; Nishi, Y.; Oda, M.; Kobayashi, Y.; Tanaka, T. Organic Ligand Binding by a Hydrophobic Cavity in a Designed Tetrameric Coiled-Coil Protein. Chem.–A Eur. J. 2009, 15, 1491–1498. [Google Scholar] [CrossRef]
- Tolbert, A.E.; Ervin, C.S.; Ruckthong, L.; Paul, T.J.; Jayasinghe-Arachchige, V.M.; Neupane, K.P.; Stuckey, J.A.; Prabhakar, R.; Pecoraro, V.L. Heteromeric Three-Stranded Coiled Coils Designed Using a Pb(Ii)(Cys)3 Template Mediated Strategy. Nat. Chem. 2020, 12, 405–411. [Google Scholar] [CrossRef]
- Woolfson, D.N. A Brief History of De Novo Protein Design: Minimal, Rational, and Computational: De Novo Protein Design. J. Mol. Biol. 2021, 433, 167160. [Google Scholar] [CrossRef]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. Silk Fibroin as a Functional Biomaterial for Drug and Gene Delivery. Pharmaceutics 2019, 11, 494. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Chen, H.; Sun, J.; Feng, F. Protein Nanocages for Delivery and Release of Luminescent Ruthenium(II) Polypyridyl Complexes. ACS Appl. Mater. Interfaces 2016, 8, 22756–22761. [Google Scholar] [CrossRef]
- Cho, K.; Wang, X.; Nie, S.; Chen, Z.; Shin, D.M. Therapeutic Nanoparticles for Drug Delivery in Cancer. Clin. Cancer Res. 2008, 14, 1310–1316. [Google Scholar] [CrossRef]
- Dawson, W.M.; Martin, F.J.O.; Rhys, G.G.; Shelley, K.L.; Brady, R.L.; Woolfson, D.N. Coiled Coils 9-to-5: Rational: De Novo Design of α-Helical Barrels with Tunable Oligomeric States. Chem. Sci. 2021, 12, 6923–6928. [Google Scholar] [CrossRef]
- Hayashi, I. The C-Terminal Region of the Plasmid Partitioning Protein TubY Is a Tetramer That Can Bind Membranes and DNA. J. Biol. Chem. 2020, 295, 17770–17780. [Google Scholar] [CrossRef]
- Peters, J.; Baumeister, W.; Lupas, A. Hyperthermostable Surface Layer Protein Tetrabrachion from the Archaebacterium Staphylothermus Marinus: Evidence for the Presence of a Right-Handed Coiled Coil Derived from the Primary Structure. J. Mol. Biol. 1996, 257, 1031–1041. [Google Scholar] [CrossRef]
- Stetefeld, J.; Jenny, M.; Schulthess, T.; Landwehr, R.; Engel, J.; Kammerer, R.A. Crystal Structure of a Naturally Occurring Parallel Right-Handed Coiled Coil Tetramer. Nat. Struct. Biol. 2000, 7, 772–776. [Google Scholar] [CrossRef]
- Veith, A.; Klingl, A.; Zolghadr, B.; Lauber, K.; Mentele, R.; Lottspeich, F.; Rachel, R.; Albers, S.-V.; Kletzin, A. Acidianus, Sulfolobus and Metallosphaera Surface Layers: Structure, Composition and Gene Expression. Mol. Microbiol. 2009, 73, 58–72. [Google Scholar] [CrossRef]
- Fiedler, T.; Fabrice, T.N.; Studer, V.; Vinet, A.; Faltova, L.; Kammerer, R.A.; Steinmetz, M.O.; Sharpe, T.; Pieters, J. Homodimerization of Coronin A through the C-Terminal Coiled-Coil Domain Is Essential for Multicellular Differentiation of Dictyostelium Discoideum. FEBS Lett. 2020, 594, 2116–2127. [Google Scholar] [CrossRef]
- Parihar, P.S.; Singh, A.; Karade, S.S.; Sahasrabuddhe, A.A.; Pratap, J.V. Structural Insights into Kinetoplastid Coronin Oligomerization Domain and F-Actin Interaction. Curr. Res. Struct. Biol. 2021, 3, 268–276. [Google Scholar] [CrossRef]
- Karade, S.S.; Ansari, A.; Srivastava, V.K.; Nayak, A.R.; Pratap, J.V. Molecular and Structural Analysis of a Mechanical Transition of Helices in the L. Donovani Coronin Coiled-Coil Domain. Int. J. Biol. Macromol. 2020, 143, 785–796. [Google Scholar] [CrossRef]
- MacFarlane, A.A.; Orriss, G.; Okun, N.; Meier, M.; Klonisch, T.; Khajehpour, M.; Stetefeld, J. The Pentameric Channel of COMPcc in Complex with Different Fatty Acids. PLoS ONE 2012, 7, e48130. [Google Scholar] [CrossRef]
- Kumar, S.; Ramappa, R.; Pamidimukkala, K.; Rao, C.D.; Suguna, K. New Tetrameric Forms of the Rotavirus NSP4 with Antiparallel Helices. Arch. Virol. 2018, 163, 1531–1547. [Google Scholar] [CrossRef] [PubMed]
- Bowman, G.D.; Nodelman, I.M.; Levy, O.; Lin, S.L.; Tian, P.; Zamb, T.J.; Udem, S.A.; Venkataraghavan, B.; Schutt, C.E. Crystal Structure of the Oligomerization Domain of NSP4 from Rotavirus Reveals a Core Metal-Binding Site. J. Mol. Biol. 2000, 304, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, S.J.; Argos, P. Cavities and Packing at Protein Interfaces. Protein Sci. 1994, 3, 2194–2206. [Google Scholar] [CrossRef]
- Graveland-Bikker, J.F.; Koning, R.I.; Koerten, H.K.; Geels, R.B.J.; Heeren, R.M.A.; De Kruif, C.G. Structural Characterization of α-Lactalbumin Nanotubes. Soft Matter 2009, 5, 2020–2026. [Google Scholar] [CrossRef]
- Shokuhfar, A.; Ghaffari, A.; Ghasemi, R.H. Cavity Control of Prefoldin Nano Actuator (PNA) by Temperature and PH. Nano-Micro Lett. 2012, 4, 110–117. [Google Scholar] [CrossRef]
- Rhys, G.G.; Wood, C.W.; Lang, E.J.M.; Mulholland, A.J.; Brady, R.L.; Thomson, A.R.; Woolfson, D.N. Maintaining and Breaking Symmetry in Homomeric Coiled-Coil Assemblies. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Bergendahl, L.T.; Marsh, J.A. Functional Determinants of Protein Assembly into Homomeric Complexes. Sci. Rep. 2017, 7, 4932. [Google Scholar] [CrossRef]
- Beesley, J.L.; Woolfson, D.N. The de Novo Design of α-Helical Peptides for Supramolecular Self-Assembly. Curr. Opin. Biotechnol. 2019, 58, 175–182. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Q.; Liu, H. DEPACT and PACMatch: A Workflow of Designing De Novo Protein Pockets to Bind Small Molecules. J. Chem. Inf. Model. 2022, 62, 971–985. [Google Scholar] [CrossRef]
- Buer, B.C.; Meagher, J.L.; Stuckey, J.A.; Marsh, E.N.G. Comparison of the Structures and Stabilities of Coiled-Coil Proteins Containing Hexafluoroleucine and t -Butylalanine Provides Insight into the Stabilizing Effects of Highly Fluorinated Amino Acid Side-Chains. Protein Sci. 2012, 21, 1705–1715. [Google Scholar] [CrossRef][Green Version]
- Tomar, D.S.; Weber, V.; Pettitt, B.M.; Asthagiri, D. Importance of Hydrophilic Hydration and Intramolecular Interactions in the Thermodynamics of Helix-Coil Transition and Helix-Helix Assembly in a Deca-Alanine Peptide. J. Phys. Chem. B 2016, 120, 69–76. [Google Scholar] [CrossRef]
- Meier, M.; Stetefeld, J.; Burkhard, P. The Many Types of Interhelical Ionic Interactions in Coiled Coils—An Overview. J. Struct. Biol. 2010, 170, 192–201. [Google Scholar] [CrossRef]
- Boyken, S.E.; Chen, Z.; Groves, B.; Langan, R.A.; Oberdorfer, G.; Ford, A.; Gilmore, J.M.; Xu, C.; Dimaio, F.; Henrique Pereira, J.; et al. De Novo Design of Protein Homo-Oligomers with Modular Hydrogen-Bond Network–Mediated Specificity. Science 2016, 352, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Stern, K.L.; Smith, M.S.; Billings, W.M.; Loftus, T.J.; Conover, B.M.; Della Corte, D.; Price, J.L. Context-Dependent Stabilizing Interactions among Solvent-Exposed Residues along the Surface of a Trimeric Helix Bundle. Biochemistry 2020, 59, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Chwastyk, M.; Panek, E.A.; Malinowski, J.; Jaskólski, M.; Cieplak, M. Properties of Cavities in Biological Structures—A Survey of the Protein Data Bank. Front. Mol. Biosci. 2020, 7, 591381. [Google Scholar] [CrossRef] [PubMed]
- Rhys, G.G.; Dawson, W.M.; Beesley, J.L.; Martin, F.J.O.; Brady, R.L.; Thomson, A.R.; Woolfson, D.N. How Coiled-Coil Assemblies Accommodate Multiple Aromatic Residues. Biomacromolecules 2021, 22, 2010–2019. [Google Scholar] [CrossRef]
- Sidorova, A.; Bystrov, V.; Lutsenko, A.; Shpigun, D.; Belova, E.; Likhachev, I. Quantitative Assessment of Chirality of Protein Secondary Structures and Phenylalanine Peptide Nanotubes. Nanomaterials 2021, 11, 3299. [Google Scholar] [CrossRef]
- McDougall, M.; Francisco, O.; Harder-Viddal, C.; Roshko, R.; Heide, F.; Sidhu, S.; Khajehpour, M.; Leslie, J.; Palace, V.; Tomy, G.T.; et al. Proteinaceous Nano Container Encapsulate Polycyclic Aromatic Hydrocarbons. Sci. Rep. 2019, 9, 1058. [Google Scholar] [CrossRef]
- Harder-Viddal, C.; Heide, F.; Roshko, R.M.; Stetefeld, J. Molecular Dynamics Simulations of Ortho-Carborane Nano-Diamond Storage within the Nonpolar Channel Cavities of a Right-Handed Coiled-Coil Tetrabrachion Nanotube. Comput. Struct. Biotechnol. J. 2021, 19, 3531–3541. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Ren, P.X.; Balusu, R.; Yang, X. Transmembrane Helices Tilt, Bend, Slide, Torque, and Unwind between Functional States of Rhodopsin. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Er, S.; Laraib, U.; Arshad, R.; Sargazi, S.; Rahdar, A.; Pandey, S.; Thakur, V.K.; Díez-Pascual, A.M. Amino Acids, Peptides, and Proteins: Implications for Nanotechnological Applications in Biosensing and Drug/Gene Delivery. Nanomaterials 2021, 11, 3002. [Google Scholar] [CrossRef] [PubMed]
- Wagschal, K.; Tripet, B.; Mant, C.; Hodges, R.S.; Lavigne, P. The Role of Position a in Determining the Stability and Oligomerization State of α-Helical Coiled Coils: 20 Amino Acid Stability Coefficients in the Hydrophobic Core of Proteins. Protein Sci. 2008, 8, 2312–2329. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zheng, Q.; Deng, Y.; Cheng, C.S.; Kallenbach, N.R.; Lu, M. A Seven-Helix Coiled Coil. Proc. Natl. Acad. Sci. USA 2006, 103, 15457–15462. [Google Scholar] [CrossRef]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed Atlas of Surface Topography of Proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [PubMed]
- Polizzi, N.F.; DeGrado, W.F. A Defined Structural Unit Enables de Novo Design of Small-Molecule-Binding Proteins. Science 2020, 369, 1227–1233. [Google Scholar] [CrossRef]
- Bao, C.; Liu, B.; Li, B.; Chai, J.; Zhang, L.; Jiao, L.; Li, D.; Yu, Z.; Ren, F.; Shi, X.; et al. Enhanced Transport of Shape and Rigidity-Tuned α-Lactalbumin Nanotubes across Intestinal Mucus and Cellular Barriers. Nano Lett. 2020, 20, 1352–1361. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Liu, P.; Wang, L.; Lin, J.; Fan, C. Biomimetic DNA Nanotubes: Nanoscale Channel Design and Applications. Angew. Chem. Int. Ed. 2019, 58, 8996–9011. [Google Scholar] [CrossRef]
- Chen, Z.; Boyken, S.E.; Jia, M.; Busch, F.; Flores-Solis, D.; Bick, M.J.; Lu, P.; VanAernum, Z.L.; Sahasrabuddhe, A.; Langan, R.A.; et al. Programmable Design of Orthogonal Protein Heterodimers. Nature 2019, 565, 106–111. [Google Scholar] [CrossRef]
- Fujita, S.; Matsuura, K. Self-Assembled Artificial Viral Capsids Bearing Coiled-Coils at the Surface. Org. Biomol. Chem. 2017, 15, 5070–5077. [Google Scholar] [CrossRef]
- Villegas, J.A.; Sinha, N.J.; Teramoto, N.; Von Bargen, C.D.; Pochan, D.J.; Saven, J.G. Computational Design of Single-Peptide Nanocages with Nanoparticle Templating. Molecules 2022, 27, 1237. [Google Scholar] [CrossRef]
- Nambiar, M.; Nepal, M.; Chmielewski, J. Self-Assembling Coiled-Coil Peptide Nanotubes with Biomolecular Cargo Encapsulation. ACS Biomater. Sci. Eng. 2019, 5, 5082–5087. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Wang, F.; Wang, S.; Kreutzberger, M.A.B.; Osinski, T.; Orlova, A.; Wall, J.S.; Zuo, X.; Egelman, E.H.; Conticello, V.P. Ambidextrous Helical Nanotubes from Self-Assembly of Designed Helical Hairpin Motifs. Proc. Natl. Acad. Sci. USA 2019, 116, 14456–14464. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Adler-Abramovich, L.; Lampel, A.; Bram, Y.; Lipstman, S.; Gazit, E. Formation of Functional Super-Helical Assemblies by Constrained Single Heptad Repeat. Nat. Commun. 2015, 6, 8615. [Google Scholar] [CrossRef] [PubMed]
- Lajoie, M.J.; Boyken, S.E.; Salter, A.I.; Bruffey, J.; Rajan, A.; Langan, R.A.; Olshefsky, A.; Muhunthan, V.; Bick, M.J.; Gewe, M.; et al. Designed Protein Logic to Target Cells with Precise Combinations of Surface Antigens. Science 2020, 369, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Polizzi, N.F.; Wu, Y.; Lemmin, T.; Maxwell, A.M.; Zhang, S.Q.; Rawson, J.; Beratan, D.N.; Therien, M.J.; DeGrado, W.F. De Novo Design of a Hyperstable Non-Natural Protein-Ligand Complex with Sub-Å Accuracy. Nat. Chem. 2017, 9, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Naudin, E.A.; Albanese, K.I.; Smith, A.J.; Mylemans, B.; Baker, E.G.; Weiner, O.D.; Andrews, D.M.; Tigue, N.; Savery, N.J.; Woolfson, D.N. From Peptides to Proteins: Coiled-Coil Tetramers to Single-Chain 4-Helix Bundles. Chem. Sci. 2022, 13, 11330–11340. [Google Scholar] [CrossRef] [PubMed]
- Slope, L.N.; Daubney, O.J.; Campbell, H.; White, S.A.; Peacock, A.F.A. Location-Dependent Lanthanide Selectivity Engineered into Structurally Characterized Designed Coiled Coils. Angew. Chem. Int. Ed. 2021, 60, 24473–24477. [Google Scholar] [CrossRef] [PubMed]
- Mandeep; Shukla, P. Microbial Nanotechnology for Bioremediation of Industrial Wastewater. Front. Microbiol. 2020, 11, 590631. [Google Scholar] [CrossRef]
- Piperopoulos, E.; Calabrese, L.; Khaskhoussi, A.; Proverbio, E.; Milone, C. Thermo-Physical Characterization of Carbon Nanotube Composite Foam for Oil Recovery Applications. Nanomaterials 2020, 10, 86. [Google Scholar] [CrossRef]
- Tanaka, M.; Aoki, K.; Haniu, H.; Kamanaka, T.; Takizawa, T.; Sobajima, A.; Yoshida, K.; Okamoto, M.; Kato, H.; Saito, N. Applications of Carbon Nanotubes in Bone Regenerative Medicine. Nanomaterials 2020, 10, 659. [Google Scholar] [CrossRef]
- Dehaghani, M.Z.; Yousefi, F.; Seidi, F.; Bagheri, B.; Mashhadzadeh, A.H.; Naderi, G.; Esmaeili, A.; Abida, O.; Habibzadeh, S.; Saeb, M.R.; et al. Encapsulation of an Anticancer Drug Isatin inside a Host Nano-Vehicle SWCNT: A Molecular Dynamics Simulation. Sci. Rep. 2021, 11, 18753. [Google Scholar] [CrossRef] [PubMed]
- Zare, H.; Ahmadi, S.; Ghasemi, A.; Ghanbari, M.; Rabiee, N.; Bagherzadeh, M.; Karimi, M.; Webster, T.J.; Hamblin, M.R.; Mostafavi, E. Carbon Nanotubes: Smart Drug/Gene Delivery Carriers. Int. J. Nanomed. 2021, 16, 1681–1706. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Park, S.H.; Lee, J.W. Applications of Functionalized Carbon Nanotubes for the Therapy and Diagnosis of Cancer. Polymers 2017, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Hossain, M.I.; Hossain, M.K.; Rubel, M.H.K.; Hossain, K.M.; Mahfuz, A.M.U.B.; Anik, M.I. Recent Progress in Nanostructured Smart Drug Delivery Systems for Cancer Therapy: A Review. ACS Appl. Bio Mater. 2022, 5, 971–1012. [Google Scholar] [CrossRef] [PubMed]
- Corbo, C.; Molinaro, R.; Parodi, A.; Toledano Furman, N.E.; Salvatore, F.; Tasciotti, E. The Impact of Nanoparticle Protein Corona on Cytotoxicity, Immunotoxicity and Target Drug Delivery. Nanomedicine 2016, 11, 81–100. [Google Scholar] [CrossRef]
- Jaiswal, S.; Manhas, A.; Pandey, A.K.; Priya, S.; Sharma, S.K. Engineered Nanoparticle-Protein Interactions Influence Protein Structural Integrity and Biological Significance. Nanomaterials 2022, 12, 1214. [Google Scholar] [CrossRef]
- Montes-Fonseca, S.L.; Sánchez-Ramírez, B.; Luna-Velasco, A.; Arzate-Quintana, C.; Silva-Cazares, M.B.; González Horta, C.; Orrantia-Borunda, E. Cytotoxicity of Protein-Carbon Nanotubes on J774 Macrophages Is a Functionalization Grade-Dependent Effect. Biomed. Res. Int. 2015, 2015, 796456. [Google Scholar] [CrossRef]
- Kim, M.T.; Chen, Y.; Marhoul, J.; Jacobson, F. Statistical Modeling of the Drug Load Distribution on Trastuzumab Emtansine (Kadcyla), a Lysine-Linked Antibody Drug Conjugate. Bioconjug. Chem. 2014, 25, 1223–1232. [Google Scholar] [CrossRef]
- Miele, E.; Spinelli, G.P.; Miele, E.; Tomao, F.; Tomao, S. Albumin-Bound Formulation of Paclitaxel (Abraxane® ABI-007) in the Treatment of Breast Cancer. Int. J. Nanomed. 2009, 4, 99–105. [Google Scholar] [CrossRef]
- Eriksson, M.; Hassan, S.; Larsson, R.; Linder, S.; Ramqvist, T.; Lövborg, H.; Vikinge, T.; Figgemeier, E.; Müller, J.; Stetefeld, J.; et al. Utilization of a Right-Handed Coiled-Coil Protein from Archaebacterium Staphylothermus Marinus as a Carrier for Cisplatin. Anticancer Res. 2009, 29, 11–18. [Google Scholar]
- Scagliotti, G.V.; Park, K.; Patil, S.; Rolski, J.; Goksel, T.; Martins, R.; Gans, S.J.M.; Visseren-Grul, C.; Peterson, P. Survival without Toxicity for Cisplatin plus Pemetrexed versus Cisplatin plus Gemcitabine in Chemonaïve Patients with Advanced Non-Small Cell Lung Cancer: A Risk-Benefit Analysis of a Large Phase III Study. Eur. J. Cancer 2009, 45, 2298–2303. [Google Scholar] [CrossRef]
- Merritt, J.C.; Richbart, S.D.; Moles, E.G.; Cox, A.J.; Brown, K.C.; Miles, S.L.; Finch, P.T.; Hess, J.A.; Tirona, M.T.; Valentovic, M.A.; et al. Anti-Cancer Activity of Sustained Release Capsaicin Formulations. Pharmacol. Ther. 2022, 238, 108177. [Google Scholar] [CrossRef]
- Youssef, R.B.; Fouad, M.A.; El-Zaher, A.A. Bioanalytical Study of the Effect of Lycopene on the Pharmacokinetics of Theophylline in Rats. Pharm. Chem. J. 2020, 53, 1053–1058. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Kao, T.-W.; Wang, C.-C.; Wu, C.-J.; Zhou, Y.-C.; Chen, W.-L. Association between Polycyclic Aromatic Hydrocarbons Exposure and Bone Turnover in Adults. Eur. J. Endocrinol. 2020, 182, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-W.; Won, Y.L.; Park, D.J.; Kim, Y.S.; Jin, E.S.; Lee, S.K. Combined Toxic Effects of Polar and Nonpolar Chemicals on Human Hepatocytes (HepG2) Cells by Quantitative Property—Activity Relationship Modeling. Toxicol. Res. 2016, 32, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, K.R.; Niitsu, A.; Kong, L.; Thomson, A.R.; Sessions, R.B.; Woolfson, D.N.; Bayley, H. A Monodisperse Transmembrane α-Helical Peptide Barrel. Nat. Chem. 2017, 9, 411–419. [Google Scholar] [CrossRef]
- Bao, G. Lanthanide Complexes for Drug Delivery and Therapeutics. J. Lumin. 2020, 228, 117622. [Google Scholar] [CrossRef]
- Heide, F.; Aprosoff, C.; Peters, L.; Palace, V.; Tomy, G.; Stetefeld, J.; McDougall, M. A Novel Passive Sampling Device for Low Molecular Weight PAHs with a Proteinaceous Medium. Environ. Nanotechnol. Monit. Manag. 2022, 17, 100609. [Google Scholar] [CrossRef]
- García-Hevia, L.; Saramiforoshani, M.; Monge, J.; Iturrioz-Rodríguez, N.; Padín-González, E.; González, F.; González-Legarreta, L.; González, J.; Fanarraga, M.L. The Unpredictable Carbon Nanotube Biocorona and a Functionalization Method to Prevent Protein Biofouling. J. Nanobiotechnology 2021, 19, 129. [Google Scholar] [CrossRef]
- Martins, C.H.Z.; Côa, F.; Da Silva, G.H.; Bettini, J.; De Farias, M.A.; Portugal, R.V.; de Aragao Umbuzeiro, G.; Alves, O.L.; Martinez, D.S.T. Functionalization of Carbon Nanotubes with Bovine Plasma Biowaste by Forming a Protein Corona Enhances Copper Removal from Water and Ecotoxicity Mitigation. Environ. Sci. Nano 2022, 9, 2887–2905. [Google Scholar] [CrossRef]
- Mann, F.A.; Lv, Z.; Großhans, J.; Opazo, F.; Kruss, S. Nanobody-Conjugated Nanotubes for Targeted Near-Infrared In Vivo Imaging and Sensing. Angew. Chem. Int. Ed. 2019, 58, 11469–11473. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.A.; Horlebein, J.; Meyer, N.F.; Meyer, D.; Thomas, F.; Kruss, S. Carbon Nanotubes Encapsulated in Coiled-Coil Peptide Barrels. Chem.—A Eur. J. 2018, 24, 12241–12245. [Google Scholar] [CrossRef] [PubMed]
- Klermund, L.; Poschenrieder, S.T.; Castiglione, K. Simple Surface Functionalization of Polymersomes Using Non-Antibacterial Peptide Anchors. J. Nanobiotechnol. 2016, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zang, C.; An, G.; Shang, M.; Cui, Z.; Chen, G.; Xi, Z.; Zhou, C. Cysteine-Specific Protein Multi-Functionalization and Disulfide Bridging Using 3-Bromo-5-Methylene Pyrrolones. Nat. Commun. 2020, 11, 1015. [Google Scholar] [CrossRef]
- Farran, B.; Montenegro, R.C.; Kasa, P.; Pavitra, E.; Huh, Y.S.; Han, Y.K.; Kamal, M.A.; Nagaraju, G.P.; Rama Raju, G.S. Folate-Conjugated Nanovehicles: Strategies for Cancer Therapy. Mater. Sci. Eng. C 2020, 107, 110341. [Google Scholar] [CrossRef]
- Kwan, H.Y.; Xu, Q.; Gong, R.; Bian, Z.; Chu, C.C. Targeted Chinese Medicine Delivery by A New Family of Biodegradable Pseudo-Protein Nanoparticles for Treating Triple-Negative Breast Cancer: In Vitro and In Vivo Study. Front. Oncol. 2021, 10, 600298. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, A.; Bernardes, G.J.L.; Shimanovich, U.; Sárria, M.P.; Nogueira, E.; Preto, A.; Gomes, A.C.; Cavaco-Paulo, A. Folic Acid-Tagged Protein Nanoemulsions Loaded with CORM-2 Enhance the Survival of Mice Bearing Subcutaneous A20 Lymphoma Tumors. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1077–1083. [Google Scholar] [CrossRef]
- Murase, S.; Ishino, S.; Ishino, Y.; Tanaka, T. Control of Enzyme Reaction by a Designed Metal-Ion-Dependent α-Helical Coiled-Coil Protein. J. Biol. Inorg. Chem. 2012, 17, 791–799. [Google Scholar] [CrossRef]
- Majerle, A.; Hadzi, S.; Aupič, J.; Satler, T.; Lapenta, F.; Strmšek, Ž.; Lah, J.; Loris, R.; Jerala, R. A Nanobody Toolbox Targeting Dimeric Coiled-Coil Modules for Functionalization of Designed Protein Origami Structures. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Gil-Garcia, M.; Ventura, S. Multifunctional Antibody-Conjugated Coiled-Coil Protein Nanoparticles for Selective Cell Targeting. Acta Biomater. 2021, 131, 472–482. [Google Scholar] [CrossRef]
- Ahn, B.; Lee, S.G.; Yoon, H.R.; Lee, J.M.; Oh, H.J.; Kim, H.M.; Jung, Y. Four-Fold Channel-Nicked Human Ferritin Nanocages for Active Drug Loading and PH-Responsive Drug Release. Angew. Chem. Int. Ed. 2018, 57, 2909–2913. [Google Scholar] [CrossRef] [PubMed]
- Agrawalla, B.K.; Wang, T.; Riegger, A.; Domogalla, M.P.; Steinbrink, K.; Dörfler, T.; Chen, X.; Boldt, F.; Lamla, M.; Michaelis, J.; et al. Chemoselective Dual Labeling of Native and Recombinant Proteins. Bioconjug. Chem. 2018, 29, 29–34. [Google Scholar] [CrossRef]
- Saleh, A.M.; Wilding, K.M.; Calve, S.; Bundy, B.C.; Kinzer-Ursem, T.L. Non-Canonical Amino Acid Labeling in Proteomics and Biotechnology. J. Biol. Eng. 2019, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, D.C.; Lee, J.J.; Link, A.J.; Graumann, J.; Tirrell, D.A.; Schuman, E.M. Labeling, Detection and Identification of Newly Synthesized Proteomes with Bioorthogonal Non-Canonical Amino-Acid Tagging. Nat. Protoc. 2007, 2, 532–540. [Google Scholar] [CrossRef]
- Jorgensen, M.D.; Chmielewski, J. Reversible Crosslinked Assembly of a Trimeric Coiled-coil Peptide into a Three-dimensional Matrix for Cell Encapsulation and Release. J. Pept. Sci. 2022, 28, e3302. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | PDB ID | Design | Number of Helices | Length (Å) | Width Diameter (Å) | Number of Cavities | Solvent-Accessible Volume (Å3) |
|---|---|---|---|---|---|---|---|
| Pb(II)C3 denovo | 6MCD | de novo | 3 | 55.2 | 24.9 | 1 | 12.3 |
| Peptide HC02 | 7P3H | de novo | 3 | 58.9 | 27.4 | 1 | 9.8 |
| alpha4tbA6 | 4G4L | de novo | 4 | 39.8 | 29.6 | 1 | 76.4 |
| PS1 | 5TGW | de novo | 4 | 45.1 | 28.4 | 1 | 215 |
| Coronin CC | 6AH6 | natural | 4 | 91.3 | 27.4 | 1 | 34 |
| Coronin CC double mutant | 6ADZ | mutant | 4 | 86.4 | 27.6 | 1 | 10.4 |
| TubY-ctd | 7C7Y | natural | 4 | 102.3 | 30.9 | 1 | 74.6 |
| ABLE | 6W70 | de novo | 4 | 51.1 | 26.0 | 1 | 181.5 |
| RHCC | 7R6H | natural | 4 | 78.9 | 27.0 | 4 | 84.8, 78.4, 57.1, 55.4 |
| COMPcc | 3V2N | natural | 5 | 72.0 | 30.4 | 2 | 166.4, 143.9 |
| CC-Pent | 4PN8 | de novo | 5 | 44.7 | 30.4 | 1 | 76.5 |
| CC-Hex2 | 4PN9 | de novo | 6 | 48.6 | 32.7 | 1 | 393 |
| CC-Hept | 4PNA | de novo | 7 | 48.8 | 31.8 | 1 | 619.3 |
| CC-Type2-II | 6G67 | de novo | 8 | 48.7 | 32.3 | 1 | 1638.1 |
| CC-Type2-(GgLaId)4 | 7BIM | de novo | 9 | 44.8 | 38.2 | 1 | 2178.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heide, F.; Stetefeld, J. A Structural Analysis of Proteinaceous Nanotube Cavities and Their Applications in Nanotechnology. Nanomaterials 2022, 12, 4080. https://doi.org/10.3390/nano12224080
Heide F, Stetefeld J. A Structural Analysis of Proteinaceous Nanotube Cavities and Their Applications in Nanotechnology. Nanomaterials. 2022; 12(22):4080. https://doi.org/10.3390/nano12224080
Chicago/Turabian StyleHeide, Fabian, and Jörg Stetefeld. 2022. "A Structural Analysis of Proteinaceous Nanotube Cavities and Their Applications in Nanotechnology" Nanomaterials 12, no. 22: 4080. https://doi.org/10.3390/nano12224080
APA StyleHeide, F., & Stetefeld, J. (2022). A Structural Analysis of Proteinaceous Nanotube Cavities and Their Applications in Nanotechnology. Nanomaterials, 12(22), 4080. https://doi.org/10.3390/nano12224080