
Camptothecin Encapsulated in β-Cyclodextrin-EDTA-Fe3O4 Nanoparticles Induce Metabolic Reprogramming Repair in HT29 Cancer Cells through Epigenetic Modulation: A Bioinformatics Approach

 , , , and
, , , and
Abstract
1. Introduction
2. Results
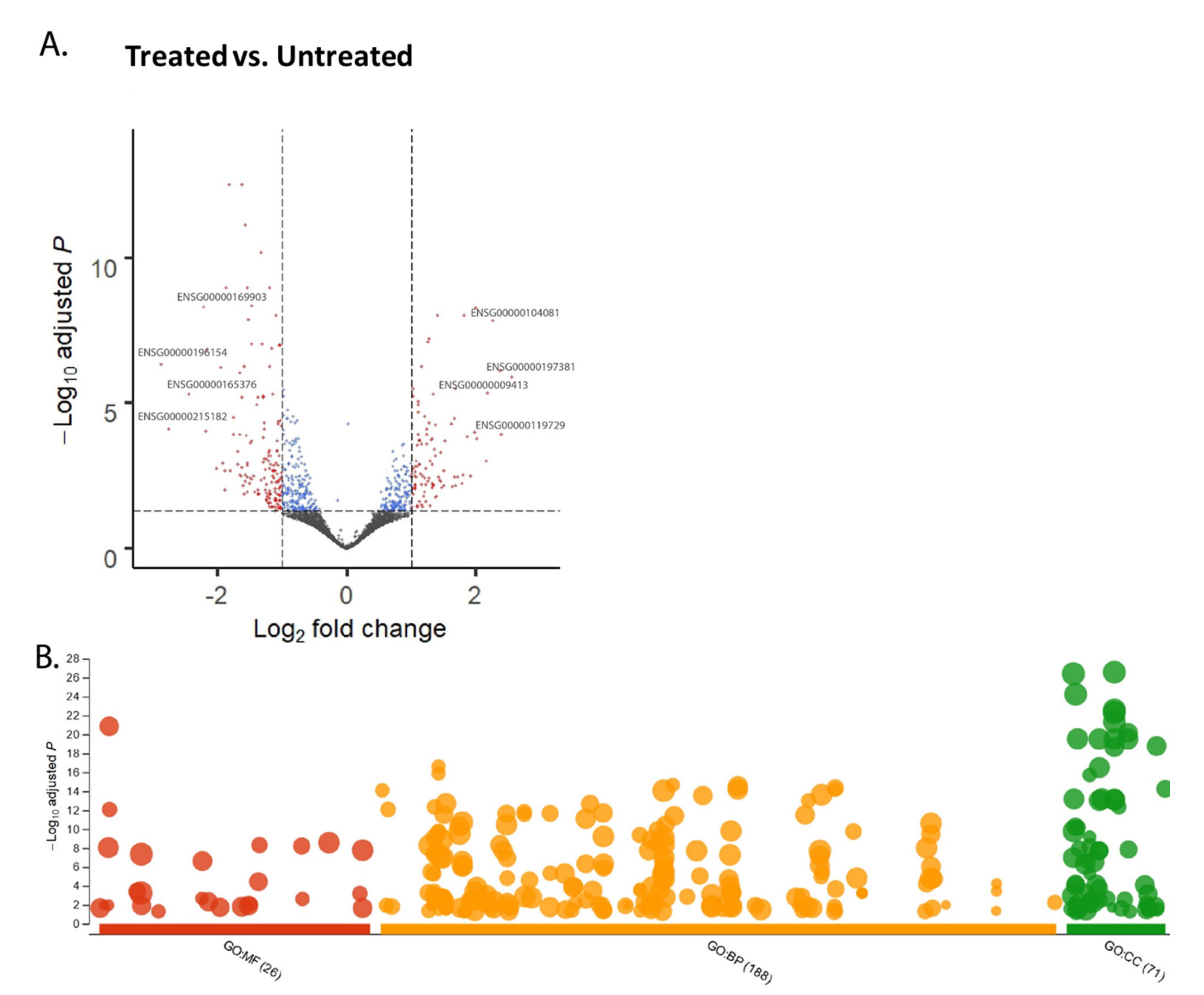
2.1. Visualization of the CPT-CEF-Treated HT29 Colon Cancer Cells through Over-Representation Analysis from RNA Sequencing Data
2.2. Identification of Genes Involved in Epigenetic Modification Differentially Regulated upon CPT-CEF Treatment of HT29 Colon Cancer Cells
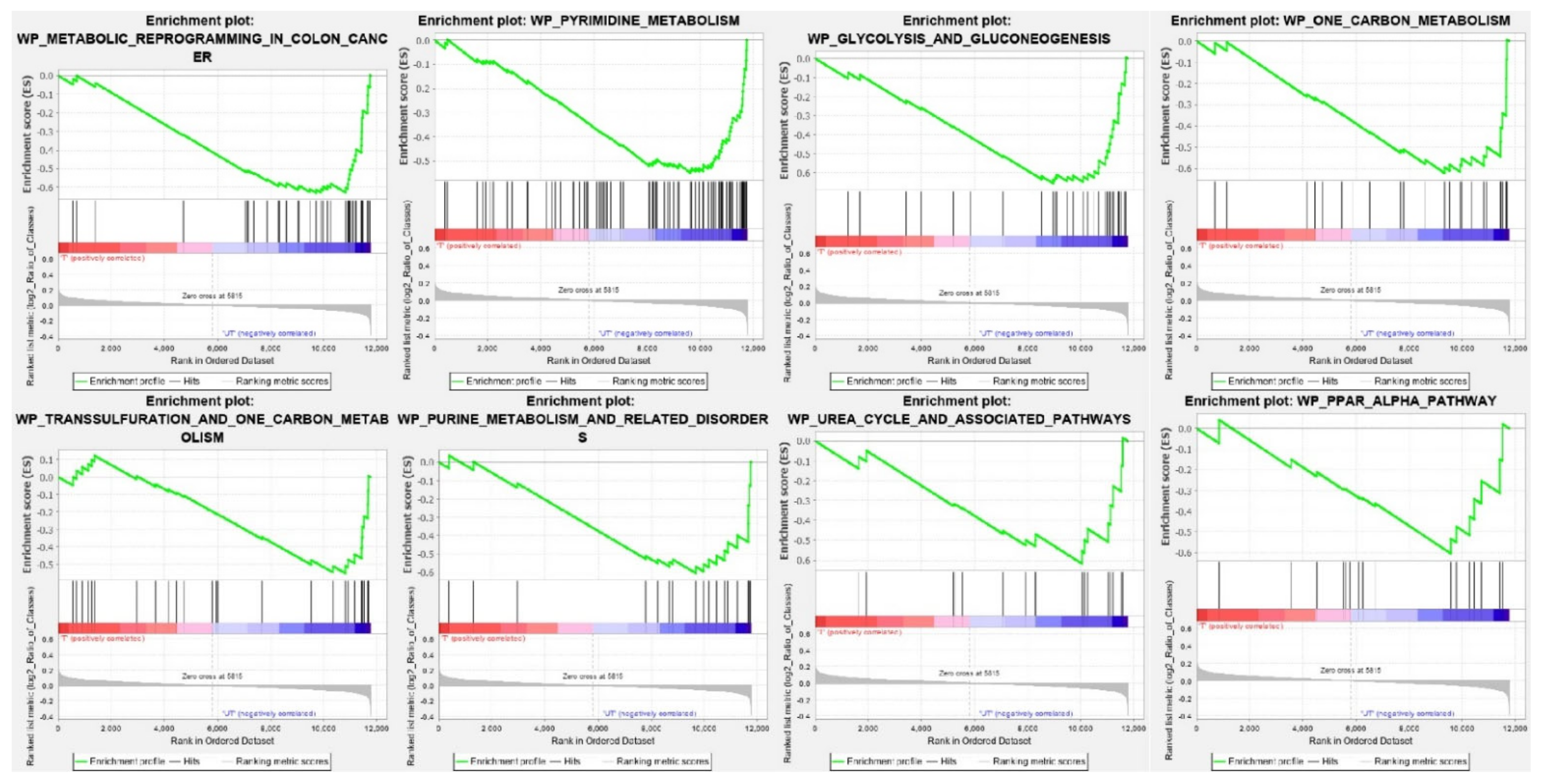
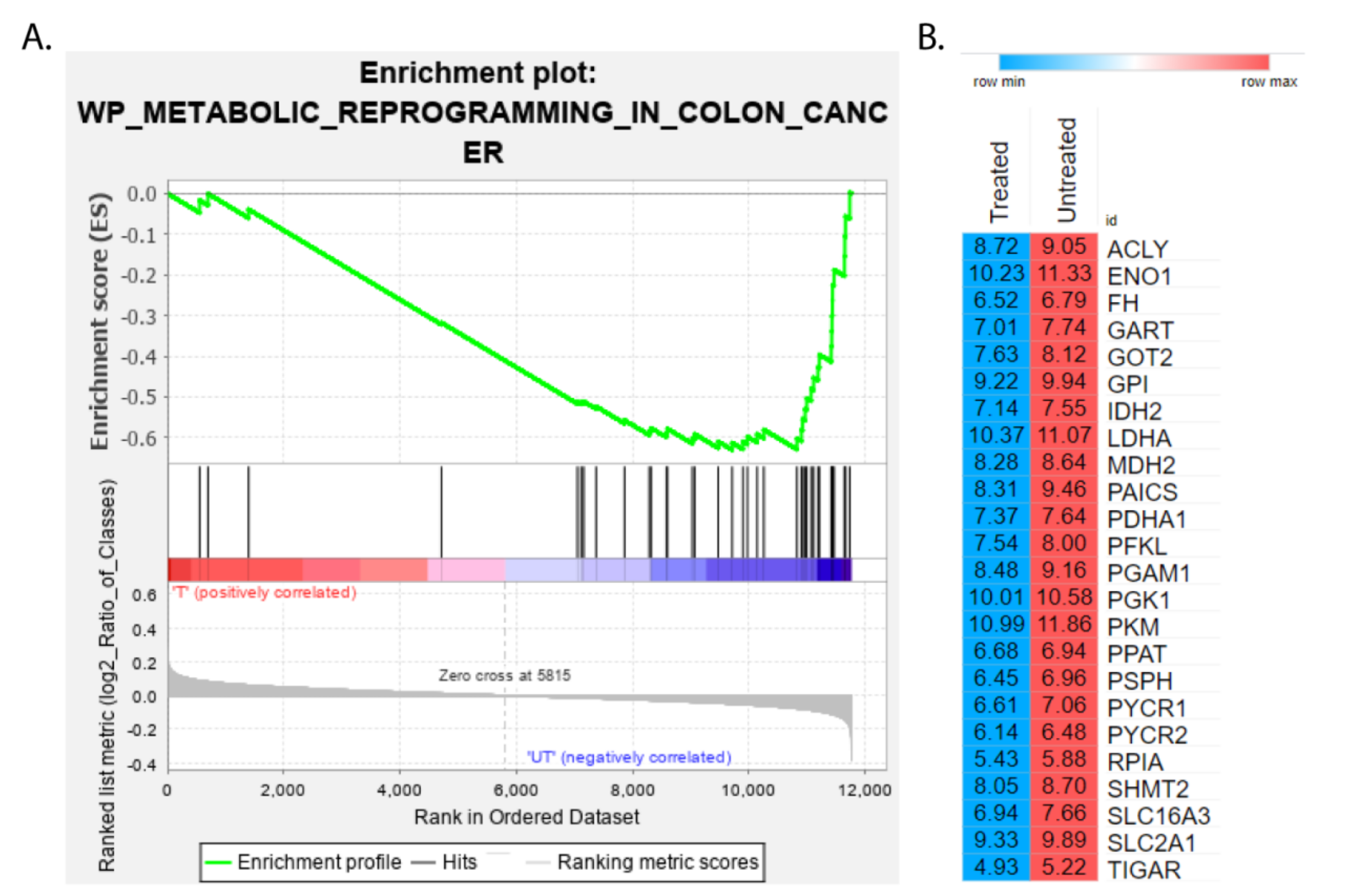
2.3. Identification of Enriched Pathways in CPT-CEF Treated HT29 Cancer Cells
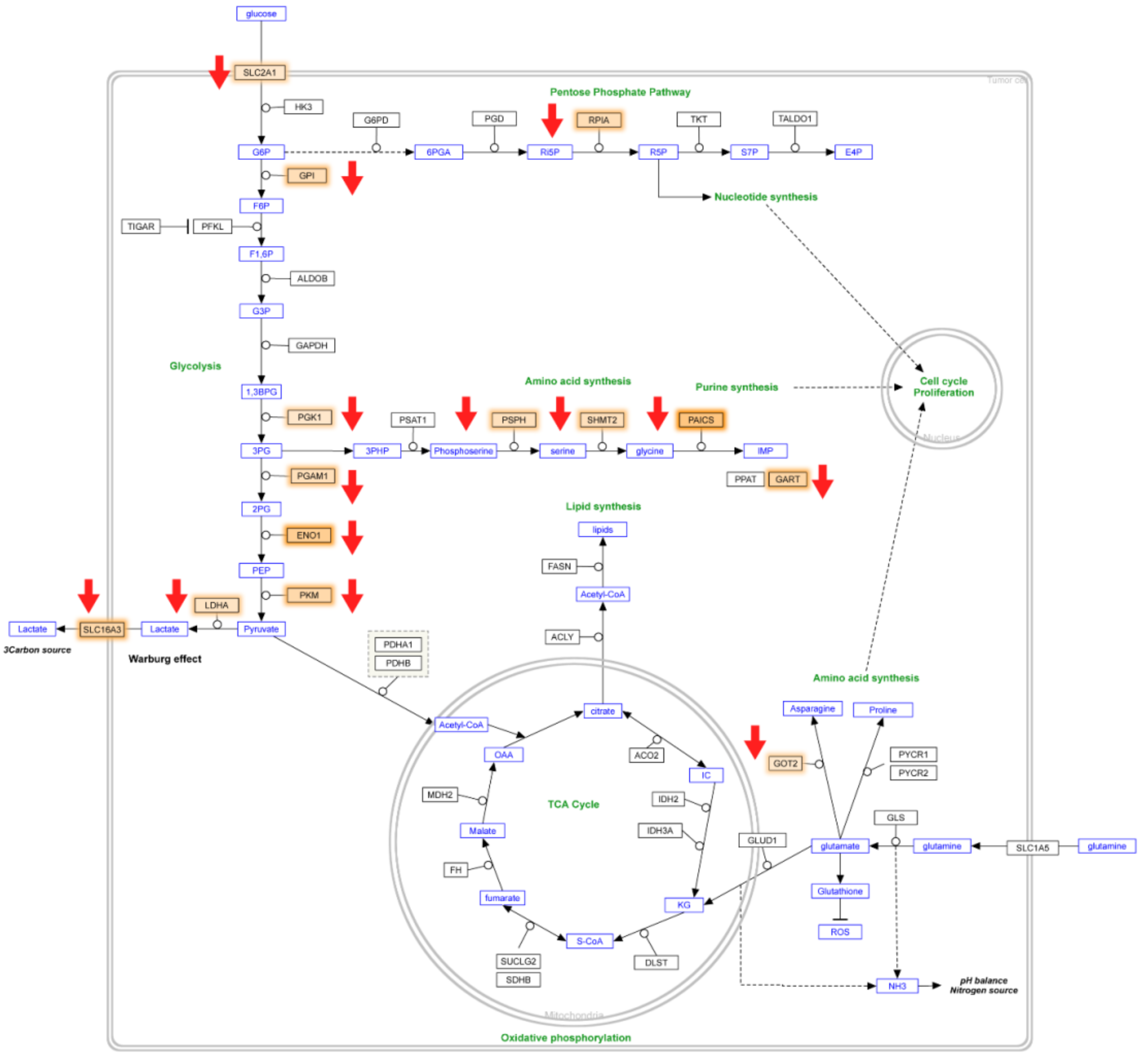
2.4. Identification of Genes with Functional Overlap between Colon Cancer Metabolism and Epigenetic Regulation
3. Discussion
4. Material and Methods
4.1. Treatment of HT29 Colon Cancer Cells with CPT-CEF Nanocompound
4.2. Isolation of Total RNA from HT29 Colon Cancer Cells
4.3. Library Preparation for RNA-Seq
4.4. RNA-Seq Data Processing and Annotation
4.5. Differentially Expressed Genes between CPT-CEF-Treated and Untreated HT29 Colon Cancer Cells
4.6. Over-Representation Analysis of Differentially Expressed Genes
4.7. Functional Enrichment Analysis of CPT-CEF-Treated HT29 Colon Cancer Cells
4.8. Identification of Associated Genes involved in Epigenetic Modifications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K–AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Moretton, A.; Loizou, J.I. Interplay between cellular metabolism and the DNA damage response in cancer. Cancers 2020, 12, 2051. [Google Scholar] [CrossRef]
- Marine, J.-C.; Dawson, S.-J.; Dawson, M.A. Non-genetic mechanisms of therapeutic resistance in cancer. Nat. Rev. Cancer 2020, 20, 743–756. [Google Scholar] [CrossRef]
- Dai, Z.; Ramesh, V.; Locasale, J.W. The evolving metabolic landscape of chromatin biology and epigenetics. Nat. Rev. Genet. 2020, 21, 737–753. [Google Scholar] [CrossRef]
- Feng, S.; De Carvalho, D.D. Clinical advances in targeting epigenetics for cancer therapy. FEBS J. 2021. [Google Scholar] [CrossRef]
- Hausser, J.; Alon, U. Tumour heterogeneity and the evolutionary trade-offs of cancer. Nat. Rev. Cancer 2020, 20, 247–257. [Google Scholar] [CrossRef]
- Wong, C.; Qian, Y.; Yu, J. Interplay between epigenetics and metabolism in oncogenesis: Mechanisms and therapeutic approaches. Oncogene 2017, 36, 3359–3374. [Google Scholar] [CrossRef]
- Li, X.; Egervari, G.; Wang, Y.; Berger, S.L.; Lu, Z. Regulation of chromatin and gene expression by metabolic enzymes and metabolites. Nat. Rev. Mol. Cell Biol. 2018, 19, 563–578. [Google Scholar] [CrossRef]
- Flavahan, W.A.; Gaskell, E.; Bernstein, B.E. Epigenetic plasticity and the hallmarks of cancer. Science 2017, 357, eaal2380. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, Z.; Cai, X. Identification of epigenetic modulators in human breast cancer by integrated analysis of DNA methylation and RNA-Seq data. Epigenetics 2018, 13, 473–489. [Google Scholar] [CrossRef]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef]
- Wang, Y.-P.; Li, J.-T.; Qu, J.; Yin, M.; Lei, Q.-Y. Metabolite sensing and signaling in cancer. J. Biol. Chem. 2020, 295, 11938–11946. [Google Scholar] [CrossRef]
- Wu, S.; Li, R.W.; Li, W.; Li, C.-J. Transcriptome characterization by RNA-seq unravels the mechanisms of butyrate-induced epigenomic regulation in bovine cells. PLoS ONE 2012, 7, e36940. [Google Scholar] [CrossRef]
- Zhou, Z.; Ibekwe, E.; Chornenkyy, Y. Metabolic alterations in cancer cells and the emerging role of oncometabolites as drivers of neoplastic change. Antioxidants 2018, 7, 16. [Google Scholar] [CrossRef]
- Shao, R.-G.; Cao, C.-X.; Nieves-Neira, W.; Dimanche-Boitrel, M.-T.; Solary, E.; Pommier, Y. Activation of the Fas pathway independently of Fas ligand during apoptosis induced by camptothecin in p53 mutant human colon carcinoma cells. Oncogene 2001, 20, 1852–1859. [Google Scholar] [CrossRef]
- Shen, X.; Chen, J.; Qiu, R.; Fan, X.; Xin, Y. Effect of camptothecin on inducible nitric oxide synthase expression in the colon cancer SW480 cell line. Oncol. Lett. 2015, 10, 3157–3160. [Google Scholar] [CrossRef][Green Version]
- Sharma, V.; Lansdell, T.A.; Peddibhotla, S.; Tepe, J.J. Sensitization of tumor cells toward chemotherapy: Enhancing the efficacy of camptothecin with imidazolines. Chem. Biol. 2004, 11, 1689–1699. [Google Scholar] [CrossRef]
- Krishnan, P.; Rajan, M.; Kumari, S.; Sakinah, S.; Priya, S.P.; Amira, F.; Danjuma, L.; Ling, M.P.; Fakurazi, S.; Arulselvan, P. Efficiency of newly formulated camptothecin with β-cyclodextrin-EDTA-Fe3O4 nanoparticle-conjugated nanocarriers as an anti-colon cancer (HT29) drug. Sci. Rep. 2017, 7, 10962. [Google Scholar] [CrossRef]
- Topuz, F.; Kilic, M.; Durgun, E.; Szekely, G. Fast-dissolving antibacterial nanofibers of cyclodextrin/antibiotic inclusion complexes for oral drug delivery. J. Colloid Interface Sci. 2021, 585, 184–194. [Google Scholar] [CrossRef]
- Ziegler, E.; Brown, A.; Nesnas, N.; Chouinard, C.; Mehta, A.; Palmer, A. β-Cyclodextrin Encapsulation of Synthetic AHLs: Drug Delivery Implications and Quorum-Quenching Exploits. ChemBioChem 2020, 22, 1292–1301. [Google Scholar] [CrossRef]
- Zhou, X.; Luo, Z.; Baidya, A.; Kim, H.J.; Wang, C.; Jiang, X.; Qu, M.; Zhu, J.; Ren, L.; Vajhadin, F.; et al. Biodegradable β-Cyclodextrin Conjugated Gelatin Methacryloyl Microneedle for Delivery of Water-Insoluble Drug. Adv. Healthc. Mater. 2020, 9, 2000527. [Google Scholar] [CrossRef]
- Phan, L.M.; Yeung, S.-C.J.; Lee, M.-H. Cancer metabolic reprogramming: Importance, main features, and potentials for precise targeted anti-cancer therapies. Cancer Biol. Med. 2014, 11, 1. [Google Scholar]
- Wong, N.; Ojo, D.; Yan, J.; Tang, D. PKM2 contributes to cancer metabolism. Cancer Lett. 2015, 356, 184–191. [Google Scholar] [CrossRef]
- Dong, G.; Mao, Q.; Xia, W.; Xu, Y.; Wang, J.; Xu, L.; Jiang, F. PKM2 and cancer: The function of PKM2 beyond glycolysis. Oncol. Lett. 2016, 11, 1980–1986. [Google Scholar] [CrossRef]
- Osthus, R.C.; Shim, H.; Kim, S.; Li, Q.; Reddy, R.; Mukherjee, M.; Xu, Y.; Wonsey, D.; Lee, L.A.; Dang, C.V. Deregulation of glucose transporter 1 and glycolytic gene expression by c-Myc. J. Biol. Chem. 2000, 275, 21797–21800. [Google Scholar] [CrossRef]
- Wise, D.R. DeBerardinis, R.J.; Mancuso, A.; Sayed, N.; Zhang, X.-Y.; Pfeiffer, H.K.; Nissim, I.; Daikhin, E.; Yudkoff, M.; McMahon, S.B. Myc regulates a transcriptional program that stimulates mitochondrial glutaminolysis and leads to glutamine addiction. Proc. Natl. Acad. Sci. USA 2008, 105, 18782–18787. [Google Scholar] [CrossRef]
- David, C.J.; Chen, M.; Assanah, M.; Canoll, P.; Manley, J.L. HnRNP proteins controlled by c-Myc deregulate pyruvate kinase mRNA splicing in cancer. Nature 2010, 463, 364–368. [Google Scholar] [CrossRef]
- Liu, W.; Hancock, C.N.; Fischer, J.W.; Harman, M.; Phang, J.M. Proline biosynthesis augments tumor cell growth and aerobic glycolysis: Involvement of pyridine nucleotides. Sci. Rep. 2015, 5, 17206. [Google Scholar] [CrossRef]
- Liu, W.; Le, A.; Hancock, C.; Lane, A.N.; Dang, C.V.; Fan, T.W.-M.; Phang, J.M. Reprogramming of proline and glutamine metabolism contributes to the proliferative and metabolic responses regulated by oncogenic transcription factor c-MYC. Proc. Natl. Acad. Sci. USA 2012, 109, 8983–8988. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Lovén, J.; Rahl, P.B.; Paranal, R.M.; Burge, C.B.; Bradner, J.E.; Lee, T.I.; Young, R.A. Transcriptional amplification in tumor cells with elevated c-Myc. Cell 2012, 151, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Sabo, A.; Kress, T.R.; Pelizzola, M.; de Pretis, S.; Gorski, M.M.; Tesi, A.; Morelli, M.J.; Bora, P.; Doni, M.; Verrecchia, A. Selective transcriptional regulation by Myc in cellular growth control and lymphomagenesis. Nature 2014, 511, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Poli, V.; Fagnocchi, L.; Fasciani, A.; Cherubini, A.; Mazzoleni, S.; Ferrillo, S.; Miluzio, A.; Gaudioso, G.; Vaira, V.; Turdo, A. MYC-driven epigenetic reprogramming favors the onset of tumorigenesis by inducing a stem cell-like state. Nat. Commun. 2018, 9, 1024. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA methyltransferases in cancer: Biology, paradox, aberrations, and targeted therapy. Cancers 2020, 12, 2123. [Google Scholar] [CrossRef]
- Sheaffer, K.L.; Elliott, E.N.; Kaestner, K.H. DNA hypomethylation contributes to genomic instability and intestinal cancer initiation. Cancer Prev. Res. 2016, 9, 534–546. [Google Scholar] [CrossRef]
- Zsigrai, S.; Kalmár, A.; Nagy, Z.B.; Barták, B.K.; Valcz, G.; Szigeti, K.A.; Galamb, O.; Dankó, T.; Sebestyén, A.; Barna, G. S-Adenosylmethionine Treatment of Colorectal Cancer Cell Lines Alters DNA Methylation, DNA Repair and Tumor Progression-Related Gene Expression. Cells 2020, 9, 1864. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Ohshima, K.; Nagashima, T.; Ohnami, S.; Ohnami, S.; Serizawa, M.; Shimoda, Y.; Maruyama, K.; Akiyama, Y.; Urakami, K. Molecular profiling and sequential somatic mutation shift in hypermutator tumours harbouring POLE mutations. Sci. Rep. 2018, 8, 8700. [Google Scholar] [CrossRef]
- Bellelli, R.; Belan, O.; Pye, V.E.; Clement, C.; Maslen, S.L.; Skehel, J.M.; Cherepanov, P.; Almouzni, G.; Boulton, S.J. POLE3-POLE4 is a histone H3-H4 chaperone that maintains chromatin integrity during DNA replication. Mol. Cell 2018, 72, 112–126.e115. [Google Scholar] [CrossRef]
- Izzo, L.T.; Affronti, H.C.; Wellen, K.E. The Bidirectional Relationship Between Cancer Epigenetics and Metabolism. Annu. Rev. Cancer Biol. 2020, 5, 235–257. [Google Scholar] [CrossRef]
- Yamada, A.; Yu, P.; Lin, W.; Okugawa, Y.; Boland, C.R.; Goel, A. A RNA-Sequencing approach for the identification of novel long non-coding RNA biomarkers in colorectal cancer. Sci. Rep. 2018, 8, 575. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Subontologies | GO Term ID | Description | Log10 Adj p |
|---|---|---|---|
| Molecular function | GO:0003735 | Structural constituent of ribosome | −12.0783 |
| GO:0005198 | Structural molecule activity | −3.4232 | |
| GO:0005488 | Binding | −3.2484 | |
| GO:0009055 | Electron carrier activity | −1.3142 | |
| GO:0050839 | Cell adhesion molecule binding | −8.2226 | |
| GO:0019843 | rRNA binding | −2.6811 | |
| GO:0043021 | Ribonucleoprotein complex binding | −2.0528 | |
| GO:0097159 | Organic cyclic compound binding | −8.567 | |
| GO:0044877 | Macromolecular complex binding | −4.4486 | |
| GO:0036094 | Small-molecule binding | −1.8064 | |
| Cellular component | GO:0005576 | Extracellular region | −2.9459 |
| GO:0005740 | Mitochondrial envelope | −4.2464 | |
| GO:0005925 | Focal adhesion | −7.9031 | |
| GO:0016020 | Membrane | −1.5299 | |
| GO:0031974 | Membrane-enclosed lumen | −19.5114 | |
| GO:0032991 | Macromolecular complex | −13.1798 | |
| GO:0043226 | Organelle | −22.2958 | |
| GO:0098589 | Membrane region | −2.1022 | |
| GO:1990904 | Ribonucleoprotein complex | −14.2526 | |
| GO:0030027 | Lamellipodium | −1.9261 | |
| Biological function | GO:0006614 | SRP-dependent cotranslational protein targeting to Membrane | −16.6126 |
| GO:0008152 | Metabolic process | −9.5346 | |
| GO:0051179 | Localization | −7.2899 | |
| GO:0051347 | Positive regulation of transferase activity | −1.9564 | |
| GO:0070482 | Response to oxygen levels | −2.8992 | |
| GO:0071840 | Cellular component organization or biogenesis | −13.6253 | |
| GO:0000184 | Nuclear-transcribed mRNA catabolic process, nonsense-Mediated decay | −14.0888 | |
| GO:0016032 | Viral process | −11.6289 | |
| GO:0006915 | Apoptotic process | −2.367 | |
| GO:0006091 | Generation of precursor metabolites and energy | −3.2792 |
| Symbol | Description | Function | Adj p | Log2FC |
|---|---|---|---|---|
| CHD6 | Chromodomain helicase DNA Binding protein 6 | Chromatin remodeling | 2.35 × 10−2 | 0.915414 |
| ARID4B | AT rich interactive domain 4B (RBP1-like) | Histone modification write cofactor | 7.71 × 10−3 | 0.912247 |
| KDM5A | Lysine (K)-specific demethylase 5A | Histone modification erase | 5.61 × 10−2 | 0.707035 |
| DAXX | Death-domain associated protein | Transcriptional regulation, cell apoptosis, carcinogenesis | 2.15 × 10−2 | −0.13739 |
| CTBP1 | C-terminal binding protein 1 | Chromatin remodeling | 4.62 × 10−2 | −0.6337 |
| HMGB1 | High mobility group box 1 | Chromatin remodeling | 1.30 × 10−2 | −0.64422 |
| MTA1 | Metastasis associated 1 | Chromatin remodeling cofactor | 5.82 × 10−2 | −0.72427 |
| DDX21 | DEAD (Asp-Glu-Ala-Asp) box helicase 21 | RNA modification | 6.45 × 10−2 | −0.73776 |
| FBL | Fibrillarin | Histone modification write | 3.82 × 10−2 | −0.89229 |
| HDGF | Hepatoma-derived growth factor | Chromatin remodeling, TF | 4.99 × 10−6 | −1.15119 |
| DNMT1 | DNA (cytosine-5-)-methyltransferase 1 | DNA modification | 8.08 × 10−4 | −1.2368 |
| FBRSL1 | Fibrosin like 1 | Histone modification | 4.16 × 10−3 | −1.46469 |
| EXOSC5 | Exosome component 5 | Scaffold protein, RNA modification | 1.79 × 10−3 | −2.01552 |
| WikiPathways ID | NES | Adj p Value |
|---|---|---|
| WP_CYTOPLASMIC_RIBOSOMAL_PROTEINS | −2.53 | 0 |
| WP_METABOLIC_REPROGRAMMING_IN_COLON_CANCER | −2.28 | 4.07 × 10−4 |
| WP_PYRIMIDINE_METABOLISM | −2.25 | 2.71 × 10−4 |
| WP_GLYCOLYSIS_AND_GLUCONEOGENESIS | −2.19 | 4.22 × 10−4 |
| WP_DNA_REPLICATION | −2.13 | 1.03 × 10−3 |
| WP_BASE_EXCISION_REPAIR | −2.11 | 1.43 × 10−3 |
| WP_DNA_IRDAMAGE_AND_CELLULAR_RESPONSE_VIA_ATR | −2.06 | 2.09 × 10−3 |
| WP_GASTRIC_CANCER_NETWORK_2 | −2.00 | 3.85 × 10−3 |
| WP_G1_TO_S_CELL_CYCLE_CONTROL | −2.00 | 3.42 × 10−3 |
| WP_MRNA_PROCESSING | −1.99 | 3.51 × 10−3 |
| WP_ONE_CARBON_METABOLISM | −1.96 | 4.97 × 10−3 |
| WP_DNA_MISMATCH_REPAIR | −1.96 | 4.63 × 10−3 |
| WP_NUCLEAR_RECEPTORS | −1.86 | 1.76 × 10−2 |
| WP_RETINOBLASTOMA_GENE_IN_CANCER | −1.85 | 1.78 × 10−2 |
| WP_TRANSSULFURATION_AND_ONE_CARBON_METABOLISM | −1.79 | 2.92 × 10−2 |
| WP_PURINE_METABOLISM_AND_RELATED_DISORDERS | −1.79 | 2.93 × 10−2 |
| WP_UREA_CYCLE_AND_ASSOCIATED_PATHWAYS | −1.77 | 3.33 × 10−2 |
| WP_PPAR_ALPHA_PATHWAY | −1.74 | 3.79 × 10−2 |
| WP_PHOTODYNAMIC_THERAPYINDUCED_HIF1_SURVIVAL_SIGNALING | −1.74 | 3.82 × 10−2 |
| WP_CELL_CYCLE | −1.69 | 5.56 × 10−2 |
| Symbol | Description | RMS | RES | Core Enrichment |
|---|---|---|---|---|
| PDHA1 | pyruvate dehydrogenase E1 subunit alpha 1 | −0.05 | −0.61 | Yes |
| ACLY | ATP citrate lyase | −0.05 | −0.61 | Yes |
| PPAT | phosphoribosyl pyrophosphate amidotransferase | −0.06 | −0.60 | Yes |
| FH | fumarate hydratase | −0.06 | −0.59 | Yes |
| MDH2 | malate dehydrogenase 2 | −0.06 | −0.58 | Yes |
| PYCR2 | pyrroline-5-carboxylate reductase 2 | −0.08 | −0.60 | Yes |
| IDH2 | isocitrate dehydrogenase (NADP(+)) 2 | −0.08 | −0.58 | Yes |
| PGK1 | phosphoglycerate kinase 1 | −0.08 | −0.55 | Yes |
| TIGAR | TP53 induced glycolysis regulatory phosphatase | −0.08 | −0.53 | Yes |
| SLC2A1 | solute carrier family 2 member 1 | −0.08 | −0.50 | Yes |
| PFKL | phosphofructokinase, liver type | −0.09 | −0.48 | Yes |
| GOT2 | glutamic-oxaloacetic transaminase 2 | −0.09 | −0.45 | Yes |
| LDHA | lactate dehydrogenase A | −0.09 | −0.43 | Yes |
| PYCR1 | pyrroline-5-carboxylate reductase 1 | −0.09 | −0.40 | Yes |
| GPI | glucose-6-phosphate isomerase | −0.11 | −0.38 | Yes |
| PSPH | phosphoserine phosphatase | −0.11 | −0.34 | Yes |
| PKM | pyruvate kinase M1/2 | −0.11 | −0.30 | Yes |
| PGAM1 | phosphoglycerate mutase 1 | −0.11 | −0.26 | Yes |
| SHMT2 | serine hydroxymethyltransferase 2 | −0.11 | −0.22 | Yes |
| RPIA | ribose 5-phosphate isomerase A | −0.11 | −0.19 | Yes |
| SLC16A3 | solute carrier family 16 member 3 | −0.14 | −0.15 | Yes |
| GART | phosphoribosylglycinamide formyltransferase, phosphoribosylglycinamide synthetase, phosphoribosylaminoimidazole synthetase | −0.14 | −0.11 | Yes |
| ENO1 | enolase 1 | −0.15 | −0.06 | Yes |
| PAICS | phosphoribosylaminoimidazole carboxylase and phosphoribosylaminoimidazolesuccinocarboxamide synthase | −0.19 | 0.00 | Yes |
| Symbol | Description | Function | Target Molecule | Adj p Value | Log2FC |
|---|---|---|---|---|---|
| PKM | pyruvate kinase, muscle | Histone modification write cofactor | histone | Yes | −1.02522 |
| DNMT1 | DNA (cytosine-5-)-methyltransferase 1 | DNA modification | DNA | Yes | −1.2368 |
| POLE3 | polymerase (DNA directed), epsilon 3, accessory subunit | Histone chaperone | histone | Yes | −0.54009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farhana, A.; Koh, A.E.-H.; Ling Mok, P.; Alsrhani, A.; Khan, Y.S.; Subbiah, S.K. Camptothecin Encapsulated in β-Cyclodextrin-EDTA-Fe3O4 Nanoparticles Induce Metabolic Reprogramming Repair in HT29 Cancer Cells through Epigenetic Modulation: A Bioinformatics Approach. Nanomaterials 2021, 11, 3163. https://doi.org/10.3390/nano11123163
Farhana A, Koh AE-H, Ling Mok P, Alsrhani A, Khan YS, Subbiah SK. Camptothecin Encapsulated in β-Cyclodextrin-EDTA-Fe3O4 Nanoparticles Induce Metabolic Reprogramming Repair in HT29 Cancer Cells through Epigenetic Modulation: A Bioinformatics Approach. Nanomaterials. 2021; 11(12):3163. https://doi.org/10.3390/nano11123163
Chicago/Turabian StyleFarhana, Aisha, Avin Ee-Hwan Koh, Pooi Ling Mok, Abdullah Alsrhani, Yusuf Saleem Khan, and Suresh Kumar Subbiah. 2021. "Camptothecin Encapsulated in β-Cyclodextrin-EDTA-Fe3O4 Nanoparticles Induce Metabolic Reprogramming Repair in HT29 Cancer Cells through Epigenetic Modulation: A Bioinformatics Approach" Nanomaterials 11, no. 12: 3163. https://doi.org/10.3390/nano11123163
APA StyleFarhana, A., Koh, A. E.-H., Ling Mok, P., Alsrhani, A., Khan, Y. S., & Subbiah, S. K. (2021). Camptothecin Encapsulated in β-Cyclodextrin-EDTA-Fe3O4 Nanoparticles Induce Metabolic Reprogramming Repair in HT29 Cancer Cells through Epigenetic Modulation: A Bioinformatics Approach. Nanomaterials, 11(12), 3163. https://doi.org/10.3390/nano11123163