

Modulating Osteoclast Activity and Immune Responses with Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes for Osteoporotic Bone Regeneration

Abstract
1. Introduction
2. Materials and Methods
2.1. Ag@TiO2-NTs Preparation
2.2. Characterization
2.3. Cell Culture and Proliferation
2.4. Cell Inflammatory Response
2.5. Macrophage–Preosteoblast Co-Culture Assay
2.6. Osteoclastogenesis Assay
2.7. In Vivo Animal Model
2.8. Micro-CT and Histological Analysis
2.9. Gene and Protein Expression
2.10. Statistical Analysis
3. Results
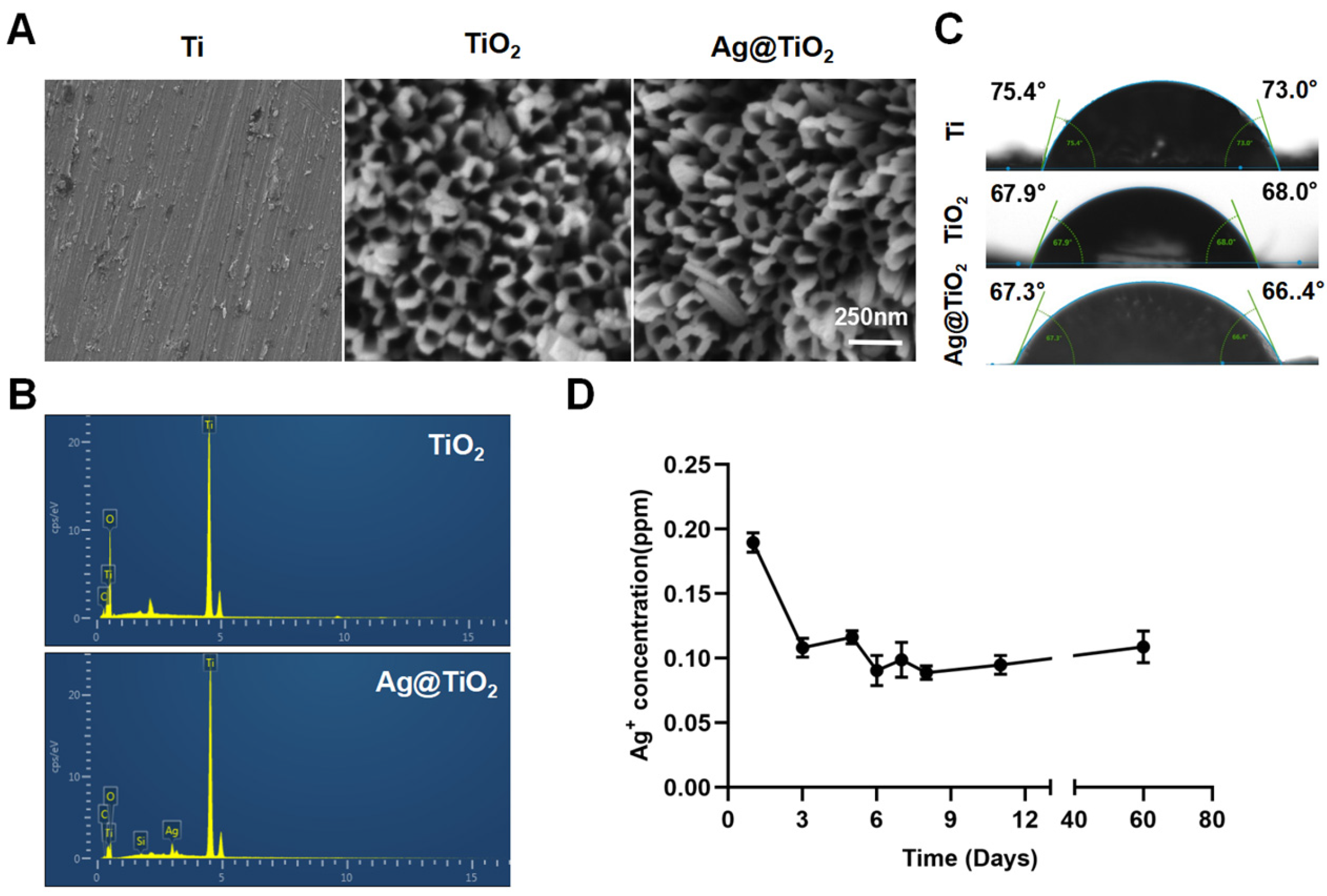
3.1. Surface Characterization, Ag+ Release, and Proliferation
3.2. Osteo-Immunoregulatory Ability In Vitro
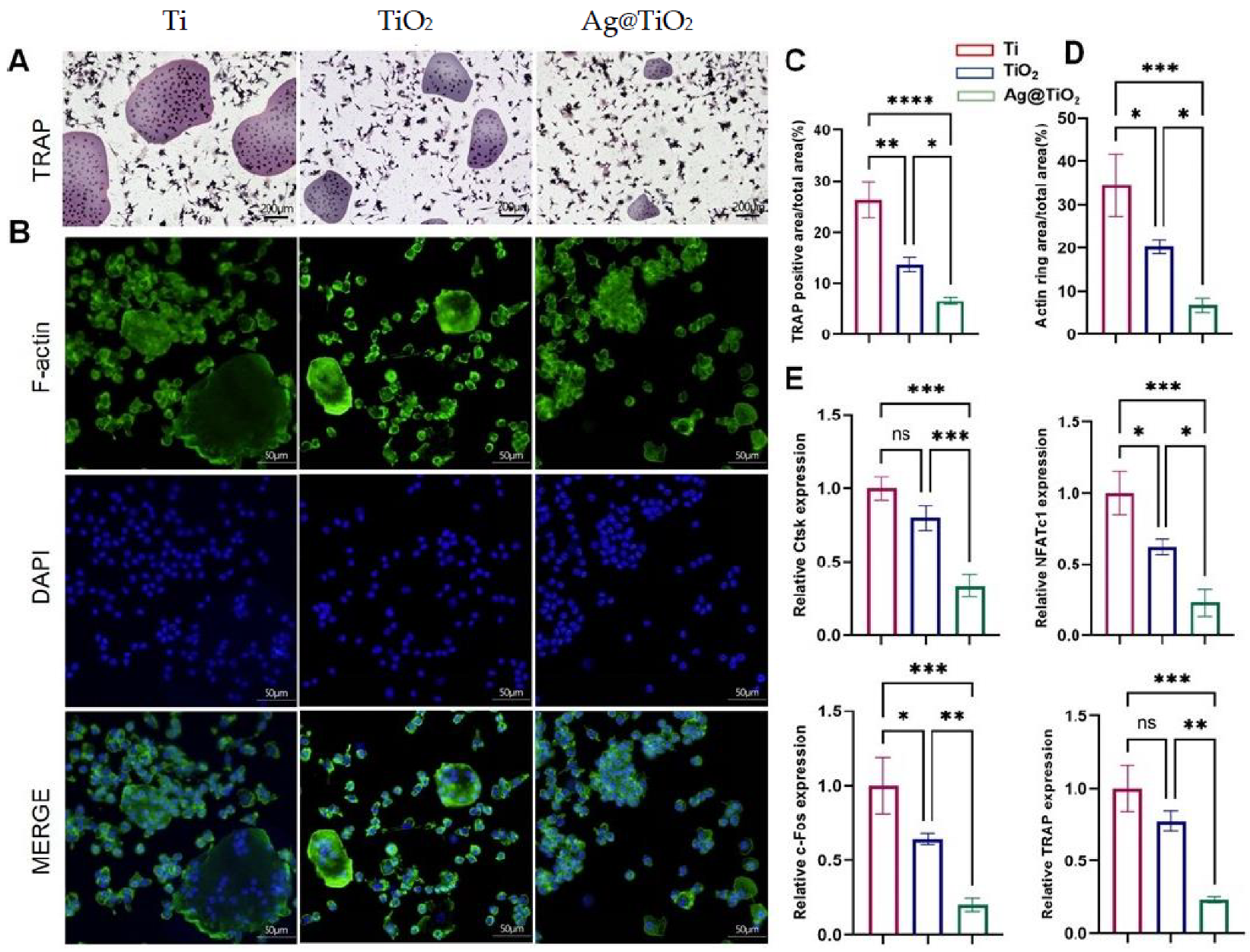
3.3. Ag@TiO2-NTs Inhibited RANKL-Induced Osteoclastogenesis In Vitro
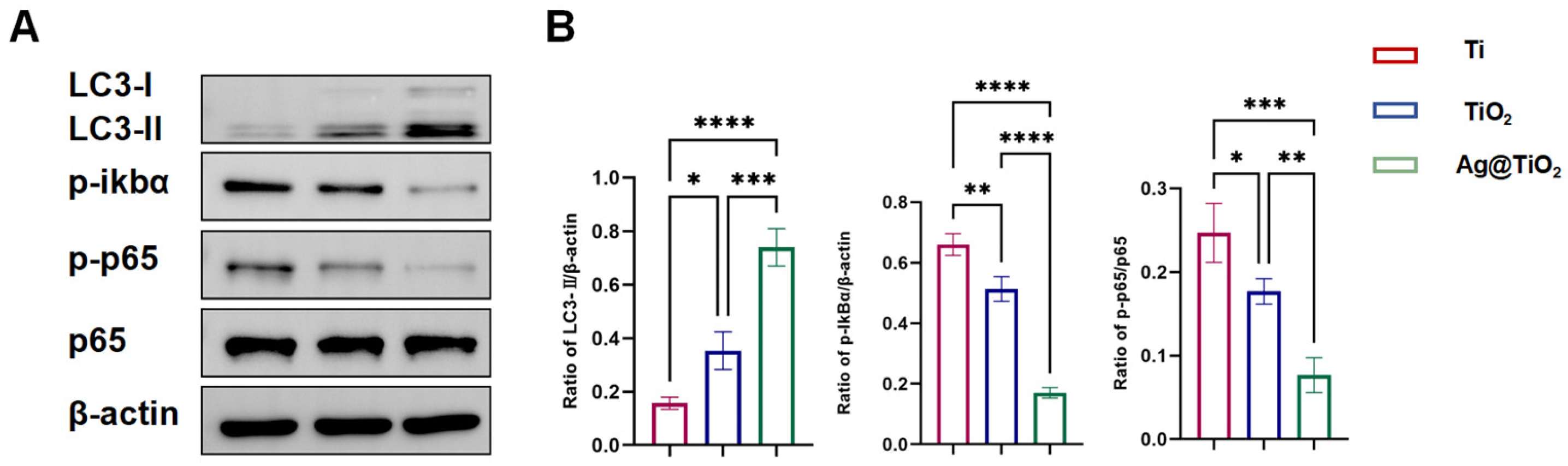
3.4. Ag@TiO2-NTs Inhibited RANKL-Induced Osteoclastogenesis Through Modulation of Autophagy and NF-κB Pathways
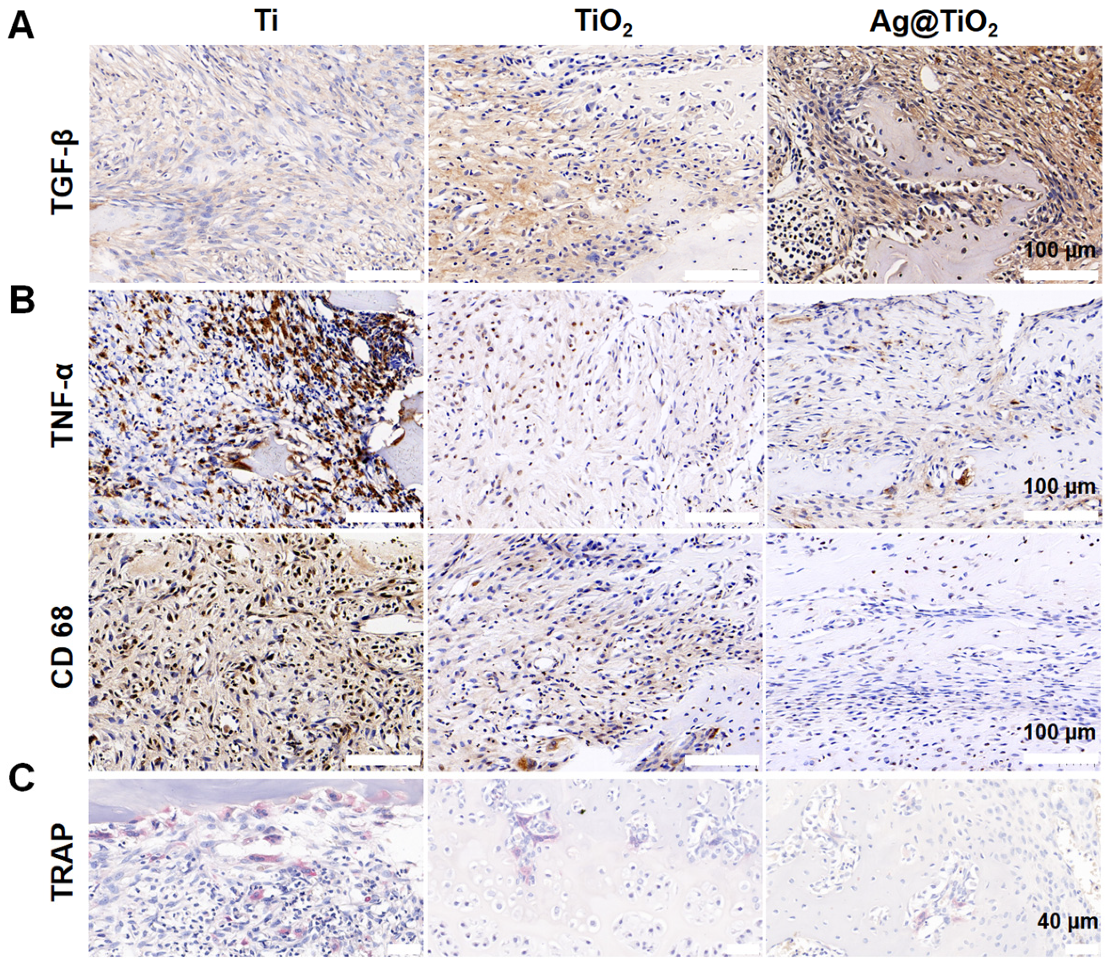
3.5. Ag@TiO2-NTs Implant Stimulated Bone Regeneration in OVX Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| iNOS | GTTCTCAGCCCAACAATACAAGA | GTGGACGGGTCGATGTCAC |
| Arg1 | CTCCAAGCCAAAGTCCTTAGAG | GGAGCTGTCATTAGGGACATCA |
| TGFa | CACTCTGGGTACGTGGGTG | CACAGGTGATAATGAGGACAGC |
| TGFb | CCACCTGCAAGACCATCGAC | CTGGCGAGCCTTAGTTTGGAC |
| ALP | GGCCTTTTACCTTTCACGGTG | TACGGCATTGTGGCTTCTCAA |
| OPG | ACCCAGAAACTGGTCATCAGC | CTGCAATACACACACTCATCACT |
| OCN | CTGAAAAGCCCACAGATACCAG | TGGAGAGGGTTGTTAGTGTGTC |
| RUNX2 | ATGCTTCATTCGCCTCACAAA | GCACTCACTGACTCGGTTGG |
| NFATc1 | GACCCGGAGTTCGACTTCG | TGACACTAGGGGACACATAACTG |
| c-Fos | CGGGTTTCAACGCCGACTA | TTGGCACTAGAGACGGACAGA |
| MMP9 | CTGGACAGCCAGACACTAAAG | CTCGCGGCAAGTCTTCAGAG |
| TRAP | CACTCCCACCCTGAGATTTGT | CATCGTCTGCACGGTTCTG |
| Ctsk | GAAGAAGACTCACCAGAAGCAG | TCCAGGTTATGGGCAGAGATT |
| GAPDH | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |


References
- Yang, T.L.; Shen, H.; Liu, A.; Dong, S.S.; Zhang, L.; Deng, F.Y.; Zhao, Q.; Deng, H.W. A road map for understanding molecular and genetic determinants of osteoporosis. Nat. Rev. Endocrinol. 2020, 16, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Ge, K.; Jin, Y.; Han, Y.; Zhang, H.; Zhou, G.; Yang, X.; Liu, D.; Liu, H.; Liang, X.J.; et al. Bone-Targeted Nanoplatform Combining Zoledronate and Photothermal Therapy To Treat Breast Cancer Bone Metastasis. ACS Nano 2019, 13, 7556–7567. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [PubMed]
- Binding, C.; Bjerring Olesen, J.; Abrahamsen, B.; Staerk, L.; Gislason, G.; Nissen Bonde, A. Osteoporotic Fractures in Patients With Atrial Fibrillation Treated With Conventional Versus Direct Anticoagulants. J. Am. Coll. Cardiol. 2019, 74, 2150–2158. [Google Scholar] [CrossRef]
- Yoo, J.E.; Shin, D.W.; Han, K.; Kim, D.; Yoon, J.W.; Lee, D.Y. Association of Female Reproductive Factors With Incidence of Fracture Among Postmenopausal Women in Korea. JAMA Netw. Open 2021, 4, e2030405. [Google Scholar] [CrossRef]
- Wauquier, F.; Leotoing, L.; Coxam, V.; Guicheux, J.; Wittrant, Y. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar] [CrossRef]
- Anesi, A.; Generali, L.; Sandoni, L.; Pozzi, S.; Grande, A. From Osteoclast Differentiation to Osteonecrosis of the Jaw: Molecular and Clinical Insights. Int. J. Mol. Sci. 2019, 20, 4925. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Stem cells and osteoporosis therapy. Cell Stem Cell 2010, 7, 553–554. [Google Scholar] [CrossRef]
- Rao, S.S.; Hu, Y.; Xie, P.L.; Cao, J.; Wang, Z.X.; Liu, J.H.; Yin, H.; Huang, J.; Tan, Y.J.; Luo, J.; et al. Omentin-1 prevents inflammation-induced osteoporosis by downregulating the pro-inflammatory cytokines. Bone Res. 2018, 6, 9. [Google Scholar] [CrossRef]
- Wang, D.; Chen, M.W.; Wei, Y.J.; Geng, W.B.; Hu, Y.; Luo, Z.; Cai, K.Y. Construction of Wogonin Nanoparticle-Containing Strontium-Doped Nanoporous Structure on Titanium Surface to Promote Osteoporosis Fracture Repair. Adv. Healthc. Mater. 2022, 11, e2201405. [Google Scholar] [CrossRef]
- Gao, Y.; Ge, W. The histone methyltransferase DOT1L inhibits osteoclastogenesis and protects against osteoporosis. Cell Death Dis. 2018, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Yang, W.; Cui, W.; Li, C.; Ma, C.; Ji, X.; Hong, J.; Qu, Z.; Chen, J.; Liu, A.; et al. Therapeutic potential and mechanisms of mesenchymal stem cell-derived exosomes as bioactive materials in tendon-bone healing. J. Nanobiotechnol 2023, 21, 14. [Google Scholar] [CrossRef]
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In sickness and in health: The functional role of extracellular vesicles in physiology and pathology in vivo: Part I: Health and Normal Physiology: Part I: Health and Normal Physiology. J. Extracell. Vesicles 2022, 11, e12151. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Liu, H.; Zhu, R.; Zhang, Z. Contribution of macrophage polarization in bone metabolism: A literature review. Cytokine 2024, 184, 156768. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Luo, F.; Du, X.; Xian, F.; Li, X. The immune cells in modulating osteoclast formation and bone metabolism. Int. Immunopharmacol. 2024, 133, 112151. [Google Scholar] [CrossRef]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64, e12465. [Google Scholar] [CrossRef]
- Zhao, Y.; Jia, L.; Zheng, Y.; Li, W. Involvement of Noncoding RNAs in the Differentiation of Osteoclasts. Stem Cells Int. 2020, 2020, 4813140. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Pan, Y.J.; Shi, X.F.; Yang, L.; Lou, Y.L. Orientin suppresses osteoclastogenesis and ameliorates ovariectomy-induced osteoporosis via suppressing ROS production. Food Sci. Nutr. 2023, 11, 5582–5595. [Google Scholar] [CrossRef]
- Liang, W.; Feng, R.; Li, X.; Duan, X.; Feng, S.; Chen, J.; Li, Y.; Chen, J.; Liu, Z.; Wang, X.; et al. A RANKL-UCHL1-sCD13 negative feedback loop limits osteoclastogenesis in subchondral bone to prevent osteoarthritis progression. Nat. Commun. 2024, 15, 8792. [Google Scholar] [CrossRef]
- Wang, N.; Li, H.; Lü, W.; Li, J.; Wang, J.; Zhang, Z.; Liu, Y. Effects of TiO2 nanotubes with different diameters on gene expression and osseointegration of implants in minipigs. Biomaterials 2011, 32, 6900–6911. [Google Scholar] [CrossRef]
- Xu, W.C.; Dong, X.; Ding, J.L.; Liu, J.C.; Xu, J.J.; Tang, Y.H.; Yi, Y.P.; Lu, C.; Yang, W.; Yang, J.S.; et al. Nanotubular TiO(2) regulates macrophage M2 polarization and increases macrophage secretion of VEGF to accelerate endothelialization via the ERK1/2 and PI3K/AKT pathways. Int. J. Nanomed. 2019, 14, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Chen, Y.; Wei, Y.; Song, H.; Gao, C.; Cheng, H.; Li, Y.; Huo, K.; Fu, J.; Xiong, W. Long-lasting bactericidal activity through selective physical puncture and controlled ions release of polydopamine and silver nanoparticles-loaded TiO(2) nanorods in vitro and in vivo. Int. J. Nanomed. 2019, 14, 2903–2914. [Google Scholar] [CrossRef] [PubMed]
- Damle, A.; Sundaresan, R.; Rajwade, J.M.; Srivastava, P.; Naik, A. A concise review on implications of silver nanoparticles in bone tissue engineering. Bio Adv. 2022, 141, 213099. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Cheng, H.; Xu, J.; Li, F.; Gao, B.; Li, Z.; Gao, C.; Huo, K.; Fu, J.; Xiong, W. Silver-loaded nanotubular structures enhanced bactericidal efficiency of antibiotics with synergistic effect in vitro and in vivo. Int. J. Nanomed. 2017, 12, 731–743. [Google Scholar] [CrossRef]
- Gao, C.; Cheng, H.; Xu, N.; Li, Y.; Chen, Y.; Wei, Y.; Gao, B.; Fu, J.; Huo, K.; Xiong, W. Poly(dopamine) and Ag nanoparticle-loaded TiO(2) nanotubes with optimized antibacterial and ROS-scavenging bioactivities. Nanomedicine 2019, 14, 803–818. [Google Scholar] [CrossRef]
- Zeng, X.; Xiong, S.; Zhuo, S.; Liu, C.; Miao, J.; Liu, D.; Wang, H.; Zhang, Y.; Wang, C.; Liu, Y. Nanosilver/poly (dl-lactic-co-glycolic acid) on titanium implant surfaces for the enhancement of antibacterial properties and osteoinductivity. Int. J. Nanomed. 2019, 14, 1849–1863. [Google Scholar] [CrossRef]
- Xue, Y.; Hong, X.; Gao, J.; Shen, R.; Ye, Z. Preparation and biological characterization of the mixture of poly(lactic-co-glycolic acid)/chitosan/Ag nanoparticles for periodontal tissue engineering. Int. J. Nanomed. 2019, 14, 483–498. [Google Scholar] [CrossRef]
- Saleh, L.S.; Bryant, S.J. The Host Response in Tissue Engineering: Crosstalk Between Immune cells and Cell-laden Scaffolds. Cur OpinBiomed Eng. 2018, 6, 58–65. [Google Scholar] [CrossRef]
- Ma, Q.L.; Zhao, L.Z.; Liu, R.R.; Jin, B.Q.; Song, W.; Wang, Y.; Zhang, Y.S.; Chen, L.H.; Zhang, Y.M. Improved implant osseointegration of a nanostructured titanium surface via mediation of macrophage polarization. Biomaterials 2014, 35, 9853–9867. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, Y.J.; Jang, J.H.; Park, J.W. Modulating macrophage polarization with divalent cations in nanostructured titanium implant surfaces. Nanotechnology 2016, 27, 085101. [Google Scholar] [CrossRef]
- Coronel, M.M.; Geusz, R.; Stabler, C.L. Mitigating hypoxic stress on pancreatic islets via in situ oxygen generating biomaterial. Biomaterials 2017, 129, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Chu, G.; Guan, M.; Jin, J.; Luo, Y.; Luo, Z.; Shi, T.; Liu, T.; Zhang, C.; Wang, Y. Mechanochemically Reprogrammed Interface Orchestrates Neutrophil Bactericidal Activity and Apoptosis for Preventing Implant-Associated Infection. Adv. Mater. 2024, 36, e2311855. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.J.; Gibon, E.; Ma, T.; Yao, Z.; Smith, R.L.; Goodman, S.B. Revision joint replacement, wear particles, and macrophage polarization. Acta Biomater 2012, 8, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Guihard, P.; Danger, Y.; Brounais, B.; David, E.; Brion, R.; Delecrin, J.; Richards, C.D.; Chevalier, S.; Rédini, F.; Heymann, D.; et al. Induction of osteogenesis in mesenchymal stem cells by activated monocytes/macrophages depends on oncostatin M signaling. Stem Cells 2012, 30, 762–772. [Google Scholar] [CrossRef]
- Jin, J.; Xia, X.; Ruan, C.; Luo, Z.; Yang, Y.; Wang, D.; Qin, Y.; Li, D.; Zhang, Y.; Hu, Y.; et al. GAPDH-Silence Microsphere via Reprogramming Macrophage Metabolism and eradicating Bacteria for Diabetic infection bone regeneration. J. Nanobiotechnol. 2024, 22, 517. [Google Scholar] [CrossRef]
- Lahiri, V.; Hawkins, W.D.; Klionsky, D.J. Watch What You (Self-) Eat: Autophagic Mechanisms that Modulate Metabolism. Cell Metabol. 2019, 29, 803–826. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Cao, J.; Wang, Y.; Anwar, N.; Zhang, Z.; Zhang, D.; Ma, Y.; Xiao, Y.; Xiao, L.; et al. The role of autophagy in bone metabolism and clinical significance. Autophagy 2023, 19, 2409–2427. [Google Scholar] [CrossRef]
- Høyer-Hansen, M.; Bastholm, L.; Szyniarowski, P.; Campanella, M.; Szabadkai, G.; Farkas, T.; Bianchi, K.; Fehrenbacher, N.; Elling, F.; Rizzuto, R.; et al. Control of macroautophagy by calcium, calmodulin-dependent kinase kinase-beta, and Bcl-2. Mol. Cell 2007, 25, 193–205. [Google Scholar] [CrossRef]
- Høyer-Hansen, M.; Nordbrandt, S.P.; Jäättelä, M. Autophagy as a basis for the health-promoting effects of vitamin D. Trends Mol. Med. 2010, 16, 295–302. [Google Scholar] [CrossRef]
- Ha, S.W.; Weitzmann, M.N.; Beck, G.R., Jr. Bioactive silica nanoparticles promote osteoblast differentiation through stimulation of autophagy and direct association with LC3 and p62. ACS Nano 2014, 8, 5898–5910. [Google Scholar] [CrossRef]
- Song, W.; Shi, M.; Dong, M.; Zhang, Y. Inducing Temporal and Reversible Autophagy by Nanotopography for Potential Control of Cell Differentiation. ACS Appl. Mater. Inter. 2016, 8, 33475–33483. [Google Scholar] [CrossRef] [PubMed]
- Florance, I.; Cordani, M.; Pashootan, P.; Moosavi, M.A.; Zarrabi, A.; Chandrasekaran, N. The impact of nanomaterials on autophagy across health and disease conditions. Cell Mol. Life Sci. 2024, 81, 184. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Yuan, L.; XuHong, J.; Tian, H.; Zhang, Y.; Deng, J.; Qi, X. Chiral nanomaterials for tumor therapy: Autophagy, apoptosis, and photothermal ablation. J. Nanobiotechnol. 2021, 19, 220. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guan, M.; Ren, R.; Gao, C.; Cheng, H.; Li, Y.; Gao, B.; Wei, Y.; Fu, J.; Sun, J.; et al. Improved Immunoregulation of Ultra-Low-Dose Silver Nanoparticle-Loaded TiO(2) Nanotubes via M2 Macrophage Polarization by Regulating GLUT1 and Autophagy. Int. J. Nanomed. 2020, 15, 2011–2026. [Google Scholar] [CrossRef]
- Zeng, X.Z.; He, L.G.; Wang, S.; Wang, K.; Zhang, Y.Y.; Tao, L.; Li, X.J.; Liu, S.W. Aconine inhibits RANKL-induced osteoclast differentiation in RAW264.7 cells by suppressing NF-κB and NFATc1 activation and DC-STAMP expression. Acta Pharmacol. Sin. 2016, 37, 255–263. [Google Scholar] [CrossRef]
- Si, Y.; Li, Y.; Gu, K.; Yin, H.; Ma, Y. Icariin ameliorates osteoporosis in ovariectomized rats by targeting Cullin 3/Nrf2/OH pathway for osteoclast inhibition. Biomed. Pharmacother. 2024, 173, 116422. [Google Scholar] [CrossRef]
- Wang, Y.; Che, L.; Chen, X.; He, Z.; Song, D.; Yuan, Y.; Liu, C. Repurpose dasatinib and quercetin: Targeting senescent cells ameliorates postmenopausal osteoporosis and rejuvenates bone regeneration. Bioact. Mater. 2023, 25, 13–28. [Google Scholar] [CrossRef]
- Li, C.; Xu, W.; Li, L.; Zhou, Y.; Yao, G.; Chen, G.; Xu, L.; Yang, N.; Yan, Z.; Zhu, C.; et al. Concrete-Inspired Bionic Bone Glue Repairs Osteoporotic Bone Defects by Gluing and Remodeling Aging Macrophages. Adv. Sci. 2024, 11, e2408044. [Google Scholar] [CrossRef]
- Zaidi, M. Skeletal remodeling in health and disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem. Biophys. 2008, 473, 201–209. [Google Scholar] [CrossRef]
- Heo, D.N.; Ko, W.K.; Moon, H.J.; Kim, H.J.; Lee, S.J.; Lee, J.B.; Bae, M.S.; Yi, J.K.; Hwang, Y.S.; Bang, J.B.; et al. Inhibition of osteoclast differentiation by gold nanoparticles functionalized with cyclodextrin curcumin complexes. ACS Nano 2014, 8, 12049–12062. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Heo, D.N.; Kim, H.J.; Ko, W.K.; Lee, S.J.; Heo, M.; Bang, J.B.; Lee, J.B.; Hwang, D.S.; Do, S.H.; et al. Inhibition of Osteoclast Differentiation and Bone Resorption by Bisphosphonate-conjugated Gold Nanoparticles. Sci. Rep. 2016, 6, 27336. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Pan, W.; Liu, Y.; Liu, J.; Xu, G.; Su, Y.; Chen, D.; Ye, X. α-Lipoic acid loaded hollow gold nanoparticles designed for osteoporosis treatment: Preparation, characterization and in vitro evaluation. Artif. Cell Nanomed. B 2023, 51, 131–138. [Google Scholar]
- Ko, W.K.; Heo, D.N.; Moon, H.J.; Lee, S.J.; Bae, M.S.; Lee, J.B.; Sun, I.C.; Jeon, H.B.; Park, H.K.; Kwon, I.K. The effect of gold nanoparticle size on osteogenic differentiation of adipose-derived stem cells. Colloid. Inter. Sci. 2015, 438, 68–76. [Google Scholar] [CrossRef]
- Liang, H.; Jin, C.; Ma, L.; Feng, X.; Deng, X.; Wu, S.; Liu, X.; Yang, C. Accelerated Bone Regeneration by Gold-Nanoparticle-Loaded Mesoporous Silica through Stimulating Immunomodulation. ACS Appl. Mater. Inter. 2019, 11, 41758–41769. [Google Scholar] [CrossRef]
- Lee, D.; Ko, W.K.; Kim, S.J.; Han, I.B.; Hong, J.B.; Sheen, S.H.; Sohn, S. Inhibitory Effects of Gold and Silver Nanoparticles on the Differentiation into Osteoclasts In Vitro. Pharmaceutics 2021, 13, 462. [Google Scholar] [CrossRef]
- Keegan, G.M.; Learmonth, I.D.; Case, C.P. A systematic comparison of the actual, potential, and theoretical health effects of cobalt and chromium exposures from industry and surgical implants. Crit. Rev. Toxicol. 2008, 38, 645–674. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, S.; Han, S.; Crawford, R.; Lu, S.; Wei, F.; Chang, J.; Wu, C.; Xiao, Y. Nanoporous microstructures mediate osteogenesis by modulating the osteo-immune response of macrophages. Nanoscale 2017, 9, 706–718. [Google Scholar] [CrossRef]
- Yan, L.; Liang, M.; Yang, T.; Ji, J.; Jose Kumar Sreena, G.S.; Hou, X.; Cao, M.; Feng, Z. The Immunoregulatory Role of Myeloid-Derived Suppressor Cells in the Pathogenesis of Rheumatoid Arthritis. Front. Immunol. 2020, 11, 568362. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, Z.; Tan, J.; Zhong, J.; Chen, L. TRAF6/ERK/p38 pathway is involved in interleukin-17-mediated autophagy to promote osteoclast precursor cell differentiation. Zhejiang Da Xue Xue Bao Zhejiang Da Xue Xue Bao Yi Xue Ban 2021, 50, 162–170. [Google Scholar] [CrossRef]
- Xing, Y.; Huang, L.; Jian, Y.; Zhang, Z.; Zhao, X.; Zhang, X.; Fu, T.; Zhang, Y.; Wang, Y.; Zhang, X. GORASP2 promotes phagophore closure and autophagosome maturation into autolysosomes. Autophagy 2025, 21, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, B.; Li, J.; Li, H.; Wang, G.; Cai, X.; Zhang, Q.; Liu, X.; Kan, C.; Wang, L.; et al. Alternative mRNA polyadenylation regulates macrophage hyperactivation via the autophagy pathway. Cell Mol. Immunol. 2024, 21, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Sun, C.; Li, N.; Huang, B.; Jiang, J.; Shen, Y.; Wang, C.; Zhao, X.; Cui, B.; Wang, C.; et al. Nanomaterials and nanotechnology for the delivery of agrochemicals: Strategies towards sustainable agriculture. J. Nanobiotechnol 2022, 20, 11. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Wang, D.; Qian, H.; Ruan, C.; Yang, Y.; Li, D.; Wang, G.; Zhu, X.; Hu, Y.; Lei, P. Precision pore structure optimization of additive manufacturing porous tantalum scaffolds for bone regeneration: A proof-of-concept study. Biomaterials 2025, 313, 122756. [Google Scholar] [CrossRef]
- Liu, X.; Wang, D.; Wang, S.; Fan, W.; Yang, Y.; Gao, P.; Chen, M.; Yang, W.; Cai, K. Promoting osseointegration by in situ biosynthesis of metal ion-loaded bacterial cellulose coating on titanium surface. Carbohydr. Polym. 2022, 297, 120022. [Google Scholar] [CrossRef]



| Section | Conclusions |
|---|---|
| Fabrication and Characterization | Ag@TiO2-NTs nanotubes are successfully fabricated with controlled Ag+ release and no significant cytotoxicity. |
| Osteo-immunoregulatory Ability In Vitro | Ag@TiO2-NTs regulates macrophage polarization toward anti-inflammatory M2 phenotype and enhances osteogenic activity. |
| Ag@TiO2-NTs Inhibited Osteoclastogenesis In Vitro | Ag@TiO2-NTs effectively inhibits osteoclastogenesis, reducing bone resorption. |
| Ag@TiO2-NTs Inhibited Osteoclastogenesis | Ag@TiO2-NTs inhibits osteoclastogenesis by promoting autophagy and suppressing the NF-κB pathway. |
| Ag@TiO2-NTs Stimulated Bone Regeneration | Ag@TiO2-NTs implants improve bone regeneration, reduce inflammation, and inhibit osteoclast activity in vivo. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Xiang, P.; Xu, Z.; Gu, M.; Zhang, R.; Li, Y.; Xin, F.; Yi, C. Modulating Osteoclast Activity and Immune Responses with Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes for Osteoporotic Bone Regeneration. J. Funct. Biomater. 2025, 16, 162. https://doi.org/10.3390/jfb16050162
Wang Z, Xiang P, Xu Z, Gu M, Zhang R, Li Y, Xin F, Yi C. Modulating Osteoclast Activity and Immune Responses with Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes for Osteoporotic Bone Regeneration. Journal of Functional Biomaterials. 2025; 16(5):162. https://doi.org/10.3390/jfb16050162
Chicago/Turabian StyleWang, Zhen, Penghui Xiang, Zhe Xu, Meiqi Gu, Rui Zhang, Yifei Li, Fei Xin, and Chengla Yi. 2025. "Modulating Osteoclast Activity and Immune Responses with Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes for Osteoporotic Bone Regeneration" Journal of Functional Biomaterials 16, no. 5: 162. https://doi.org/10.3390/jfb16050162
APA StyleWang, Z., Xiang, P., Xu, Z., Gu, M., Zhang, R., Li, Y., Xin, F., & Yi, C. (2025). Modulating Osteoclast Activity and Immune Responses with Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes for Osteoporotic Bone Regeneration. Journal of Functional Biomaterials, 16(5), 162. https://doi.org/10.3390/jfb16050162