
Tumor-Treating Fields Alter Nanomechanical Properties of Pancreatic Ductal Adenocarcinoma Cells Co-Cultured with Extracellular Matrix

 ,
, 
 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Methods
2.1. ECM Gel Preparation
2.2. Cell Culture
2.3. TTFields Treatment
2.4. AFM
2.5. Confocal Microscopy
2.6. Data Analysis
3. Results
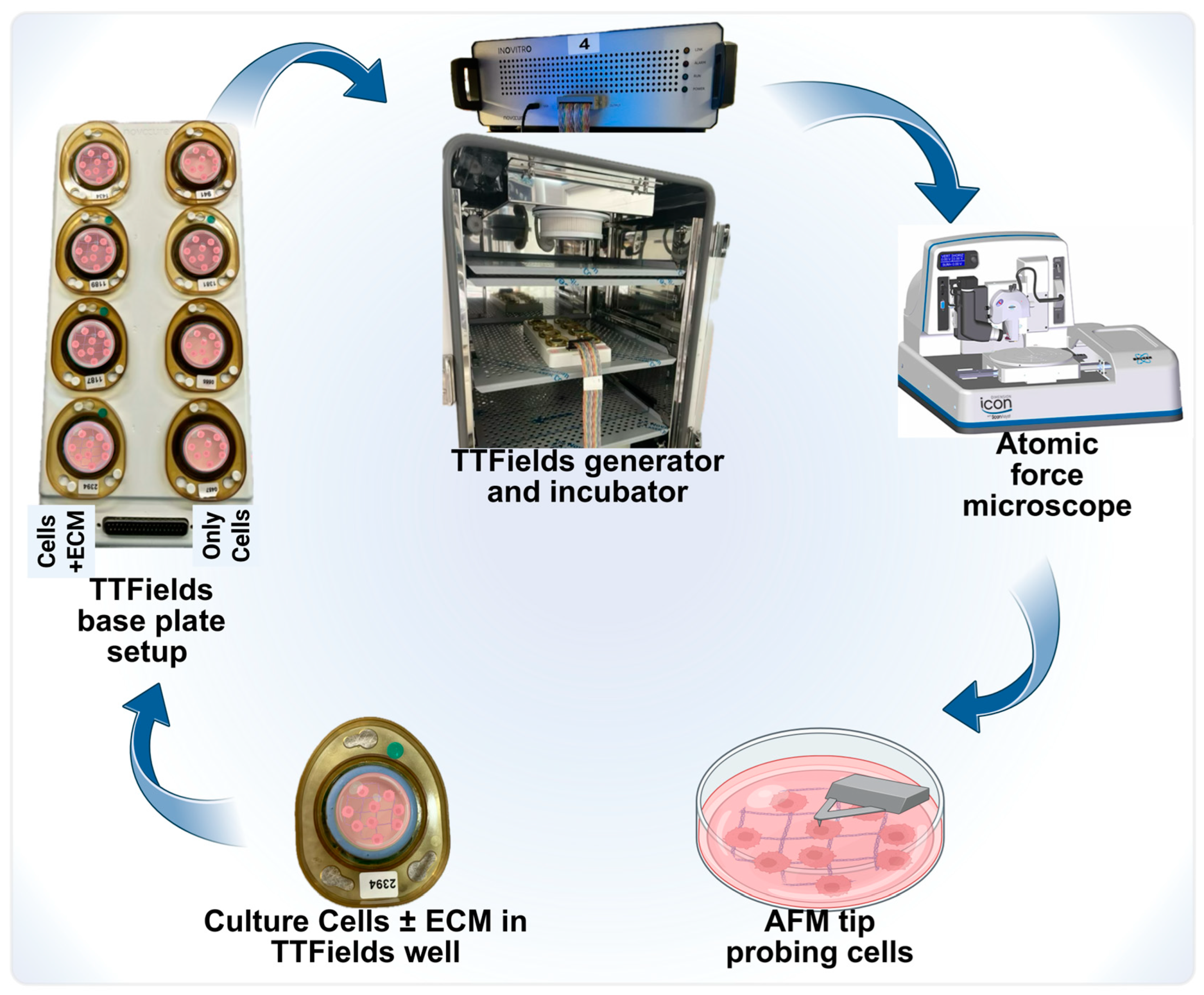
3.1. Application of TTFields to Study Cellular Nanomechanical Properties Using the AFM
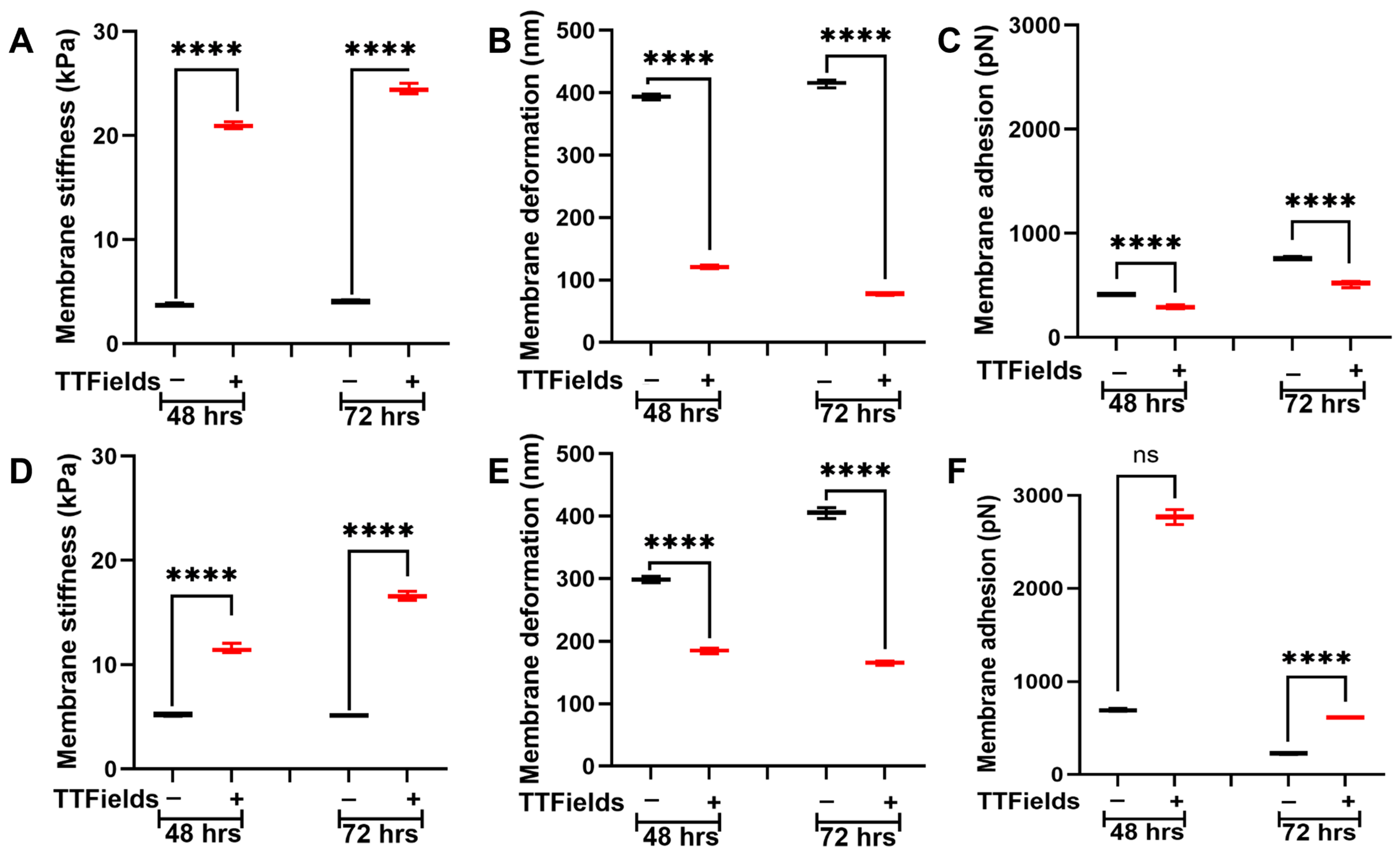
3.2. TTFields Alter Pancreatic Cancer Cell’s Nanomechanical Properties
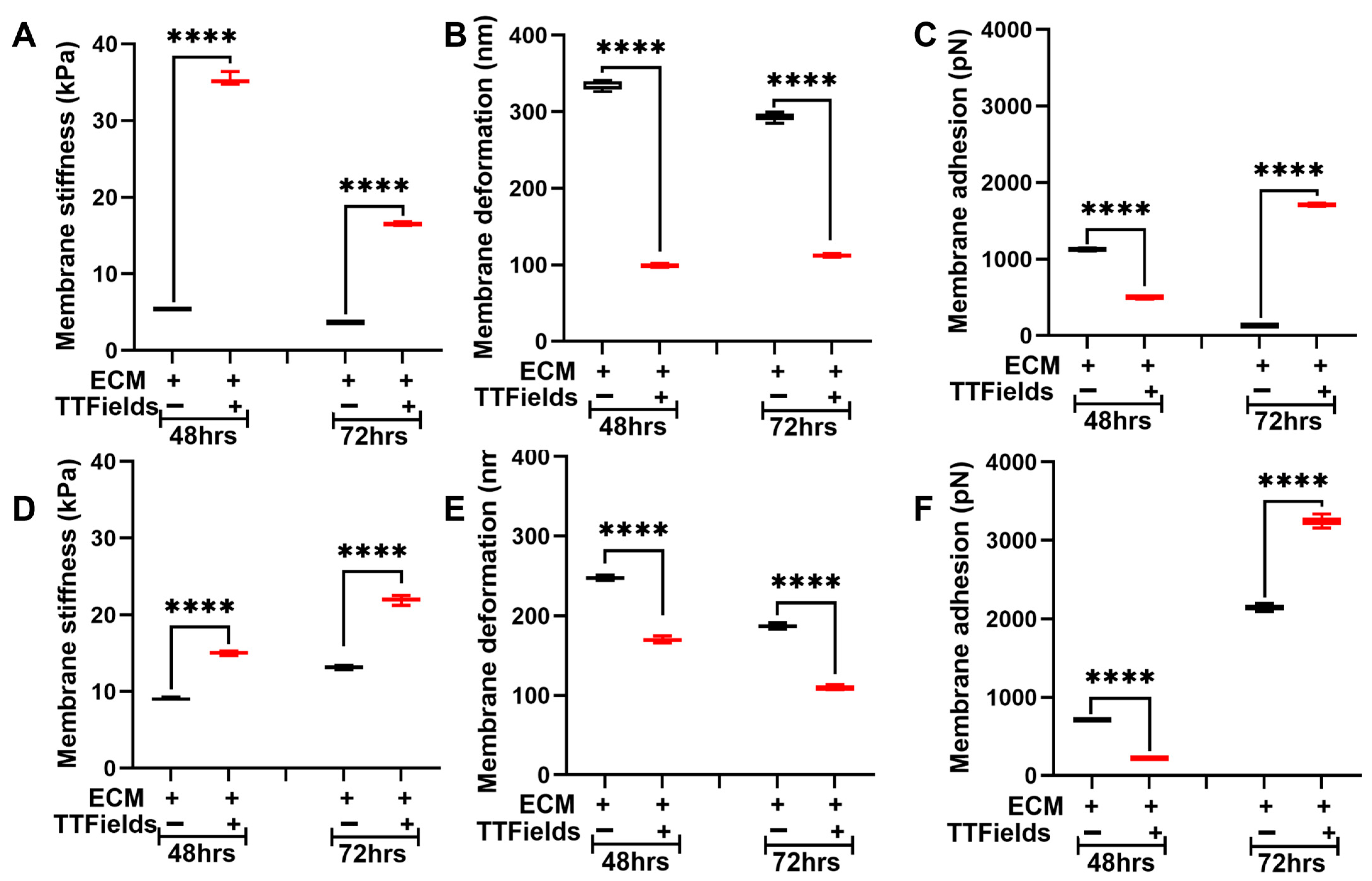
3.3. TTFields Alter Pancreatic Cancer Cell’s Nanomechanical Properties When Co-Cultured with the ECM
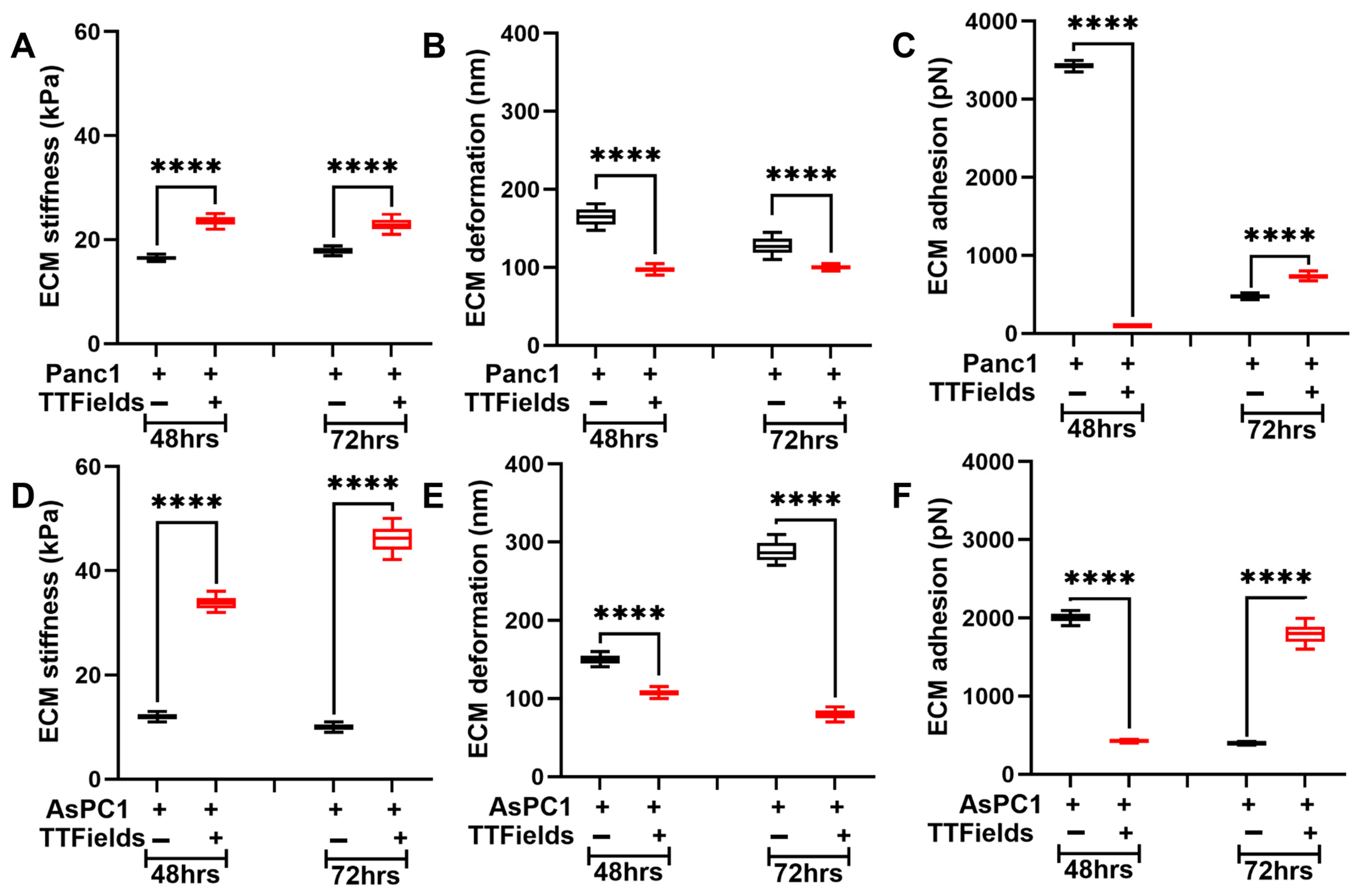
3.4. TTFields Alter ECM’s Nanomechanical Properties When in Co-Culture with Pancreatic Cancer Cells
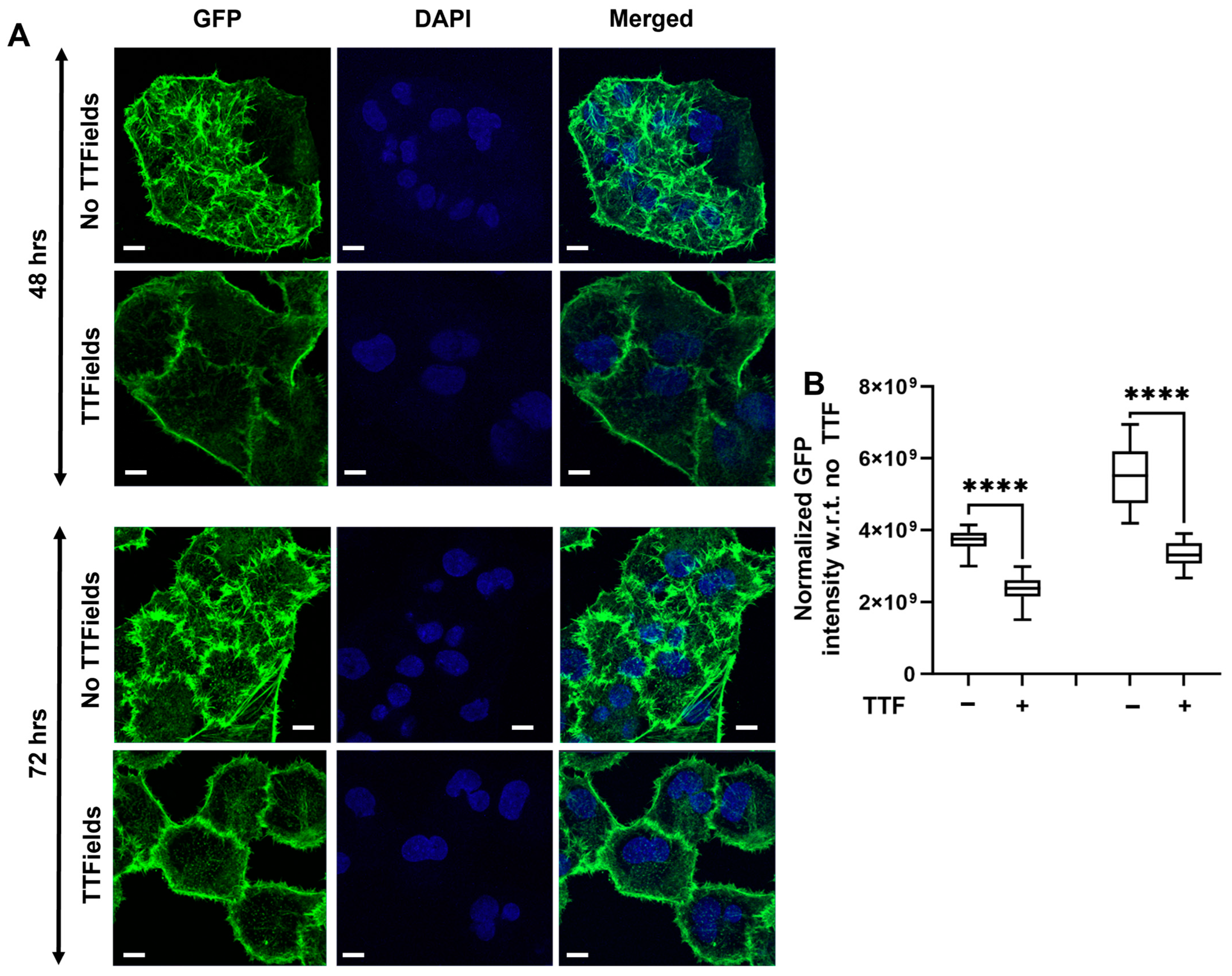
3.5. Confirmation of Stiffness Alterations in the Panc1 Cells Attributed to Dynamic Actin Rearrangement
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vergote, I.; Macarulla, T.; Hirsch, F.R.; Hagemann, C.; Miller, D.S. Tumor Treating Fields (TTFields) Therapy Concomitant with Taxanes for Cancer Treatment. Cancers 2023, 15, 636. [Google Scholar] [CrossRef]
- Voloshin, T.; Schneiderman, R.S.; Volodin, A.; Shamir, R.R.; Kaynan, N.; Zeevi, E.; Koren, L.; Klein-Goldberg, A.; Paz, R.; Giladi, M.; et al. Tumor Treating Fields (TTFields) Hinder Cancer Cell Motility Through Regulation of Microtubule and Acting Dynamics. Cancers 2020, 12, 3016. [Google Scholar] [CrossRef]
- Mun, E.J.; Babiker, H.M.; Weinberg, U.; Kirson, E.D.; Von Hoff, D.D. Tumor-Treating Fields: A Fourth Modality in Cancer Treatment. Clin. Cancer Res. 2018, 24, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Ramos, A.; Sadeghi, S.; Tabatabaeian, H. Battling Chemoresistance in Cancer: Root Causes and Strategies to Uproot Them. Int. J. Mol. Sci. 2021, 22, 9451. [Google Scholar] [CrossRef]
- Gera, N.; Yang, A.; Holtzman, T.S.; Lee, S.X.; Wong, E.T.; Swanson, K.D. Tumor treating fields perturb the localization of septins and cause aberrant mitotic exit. PLoS ONE 2015, 10, e0125269. [Google Scholar] [CrossRef] [PubMed]
- Giladi, M.; Schneiderman, R.S.; Voloshin, T.; Porat, Y.; Munster, M.; Blat, R.; Sherbo, S.; Bomzon, Z.; Urman, N.; Itzhaki, A.; et al. Mitotic Spindle Disruption by Alternating Electric Fields Leads to Improper Chromosome Segregation and Mitotic Catastrophe in Cancer Cells. Sci. Rep. 2015, 5, 18046. [Google Scholar] [CrossRef]
- Voloshin, T.; Kaynan, N.; Davidi, S.; Porat, Y.; Shteingauz, A.; Schneiderman, R.S.; Zeevi, E.; Munster, M.; Blat, R.; Tempel Brami, C.; et al. Tumor-treating fields (TTFields) induce immunogenic cell death resulting in enhanced antitumor efficacy when combined with anti-PD-1 therapy. Cancer Immunol. Immunother. 2020, 69, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Karanam, N.K.; Srinivasan, K.; Ding, L.; Sishc, B.; Saha, D.; Story, M.D. Tumor-treating fields elicit a conditional vulnerability to ionizing radiation via the downregulation of BRCA1 signaling and reduced DNA double-strand break repair capacity in non-small cell lung cancer cell lines. Cell Death Dis. 2017, 8, e2711. [Google Scholar] [CrossRef]
- Karanam, N.K.; Ding, L.; Aroumougame, A.; Story, M.D. Tumor treating fields cause replication stress and interfere with DNA replication fork maintenance: Implications for cancer therapy. Transl. Res. 2020, 217, 33–46. [Google Scholar] [CrossRef]
- Shteingauz, A.; Porat, Y.; Voloshin, T.; Schneiderman, R.S.; Munster, M.; Zeevi, E.; Kaynan, N.; Gotlib, K.; Giladi, M.; Kirson, E.D.; et al. AMPK-dependent autophagy upregulation serves as a survival mechanism in response to Tumor Treating Fields (TTFields). Cell Death Dis. 2018, 9, 1074. [Google Scholar] [CrossRef]
- Arvind, R.; Chandana, S.R.; Borad, M.J.; Pennington, D.; Mody, K.; Babiker, H. Tumor-Treating Fields: A fourth modality in cancer treatment, new practice updates. Crit. Rev. Oncol. Hematol. 2021, 168, 103535. [Google Scholar] [CrossRef]
- Sarantis, P.; Koustas, E.; Papadimitropoulou, A.; Papavassiliou, A.G.; Karamouzis, M.V. Pancreatic ductal adenocarcinoma: Treatment hurdles, tumor microenvironment and immunotherapy. World J. Gastrointest. Oncol. 2020, 12, 173–181. [Google Scholar] [CrossRef]
- Stoop, T.F.; Theijse, R.T.; Seelen, L.W.; Groot Koerkamp, B.; van Eijck, C.H.; Wolfgang, C.L.; van Tienhoven, G.; van Santvoort, H.C.; Molenaar, I.Q.; Wilmink, J.W. Preoperative chemotherapy, radiotherapy and surgical decision-making in patients with borderline resectable and locally advanced pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 101–124. [Google Scholar] [CrossRef]
- Kulkarni, T.; Robinson, O.-M.; Dutta, A.; Mukhopadhyay, D.; Bhattacharya, S. Machine learning-based approach for automated classification of cell and extracellular matrix using nanomechanical properties. Mater. Today Bio. 2024, 25, 100970. [Google Scholar] [CrossRef]
- Hare, J.I.; Lammers, T.; Ashford, M.B.; Puri, S.; Storm, G.; Barry, S.T. Challenges and strategies in anti-cancer nanomedicine development: An industry perspective. Adv. Drug Deliv. Rev. 2017, 108, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, A.A.; Perz, M.T.; Xu, Y.; Cedillo, L.G.; Huang, O.D.; McIntyre, C.A.; Vudatha, V.; Trevino, J.G.; Liu, J.; Wang, P. Unveiling the Promise: Navigating Clinical Trials 1978–2024 for PDAC. Cancers 2024, 16, 3564. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.E.; Jin, Y.-S.; Tondre, J.; Wong, R.; Rao, J.; Gimzewski, J.K. AFM-based analysis of human metastatic cancer cells. Nanotechnology 2008, 19, 384003. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, T.; Tam, A.; Mukhopadhyay, D.; Bhattacharya, S. AFM study: Cell cycle and probe geometry influences nanomechanical characterization of Panc1 cells. Biochim. Biophys. Acta (BBA) Gen. Subj. 2019, 1863, 802–812. [Google Scholar] [CrossRef]
- Kulkarni, T.; Mukhopadhyay, D.; Bhattacharya, S. Nanomechanical insight of pancreatic cancer cell membrane during receptor mediated endocytosis of targeted gold nanoparticles. ACS Appl. Bio. Mater. 2020, 4, 984–994. [Google Scholar] [CrossRef]
- Kulkarni, T.; Mukhopadhyay, D.; Bhattacharya, S. Dynamic alteration of poroelastic attributes as determinant membrane nanorheology for endocytosis of organ specific targeted gold nanoparticles. J. Nanobiotechnol. 2022, 20, 74. [Google Scholar] [CrossRef]
- Rivera, F.; Benavides, M.; Gallego, J.; Guillen-Ponce, C.; Lopez-Martin, J.; Küng, M. Tumor treating fields in combination with gemcitabine or gemcitabine plus nab-paclitaxel in pancreatic cancer: Results of the PANOVA phase 2 study. Pancreatology 2019, 19, 64–72. [Google Scholar] [CrossRef]
- Kan, T.; Khalil, T.H.; Barsheshet, Y.; Voloshin, T.; Vorontsov, A.; Brant, B.; Koren, L.; Blatt, R.; Cahal, S.; Zemer-Tov, E. 772 Induction of an anti-tumor immune response by tumor treating fields (TTFields) in a pancreatic cancer mouse model. BMJ Spec. J. 2024. [Google Scholar] [CrossRef]
- Derjaguin, B.V.; Muller, V.M.; Toporov, Y.P. Effect of contact deformations on the adhesion of particles. J. Colloid. Interface Sci. 1975, 53, 314–326. [Google Scholar] [CrossRef]
- Davies, A.M.; Weinberg, U.; Palti, Y. Tumor treating fields: A new frontier in cancer therapy. Ann. N. Y. Acad. Sci. 2013, 1291, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Mierke, C.T. Extracellular matrix cues regulate mechanosensing and mechanotransduction of cancer cells. Cells 2024, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.D.; Gerlach, B.D. The roles and regulation of the actin cytoskeleton, intermediate filaments and microtubules in smooth muscle cell migration. Respir. Res. 2017, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Rominiyi, O.; Vanderlinden, A.; Clenton, S.J.; Bridgewater, C.; Al-Tamimi, Y.; Collis, S.J. Tumour treating fields therapy for glioblastoma: Current advances and future directions. Br. J. Cancer 2021, 124, 697–709. [Google Scholar] [CrossRef]
- Liu, S.; Shi, W.; Zhao, Q.; Zheng, Z.; Liu, Z.; Meng, L.; Dong, L.; Jiang, X. Progress and prospect in tumor treating fields treatment of glioblastoma. Biomed. Pharmacother. 2021, 141, 111810. [Google Scholar] [CrossRef]
- Shah, S.; Nag, A.; Lucke-Wold, B. Association of tumor treating fields (TTFields) therapy with overall survival in newly diagnosed glioblastoma. Clin. Transl. Oncol. 2025, 1–9. [Google Scholar] [CrossRef]
- Ghiaseddin, A.P.; Shin, D.; Melnick, K.; Tran, D.D. Tumor treating fields in the management of patients with malignant gliomas. Curr. Treat. Options Oncol. 2020, 21, 76. [Google Scholar] [CrossRef]
- Kutuk, T.; Atak, E.; Kaiser, A.; Chuong, M.D. Emerging potential of tumor treating fields as a treatment modality for pancreatic cancer: A narrative review of therapeutic evolution. Dig. Med. Res. 2024, 7, 11. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, L.; Wan, D.; Zhou, L.; Zheng, S.; Lin, S.; Qiao, Y. Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduct. Target. Ther. 2021, 6, 153. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Kanchanawong, P.; Calderwood, D.A. Organization, dynamics and mechanoregulation of integrin-mediated cell–ECM adhesions. Nat. Rev. Mol. Cell Biol. 2023, 24, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Halper, J.; Kjaer, M. Basic components of connective tissues and extracellular matrix: Elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. In Progress in Heritable Soft Connective Tissue Diseases; Springer: Dordrecht, The Netherlands, 2014; pp. 31–47. [Google Scholar]
- Yoon, Y.N.; Lee, D.-S.; Park, H.J.; Kim, J.-S. Barium titanate nanoparticles sensitise treatment-resistant breast cancer cells to the antitumor action of tumour-treating fields. Sci. Rep. 2020, 10, 2560. [Google Scholar] [CrossRef]
- Tian, J.; Mallinger, J.C.; Shi, P.; Ling, D.; Deleyrolle, L.P.; Lin, M.; Khoshbouei, H.; Sarkisian, M.R. Aurora kinase A inhibition plus Tumor Treating Fields suppress glioma cell proliferation in a cilium-independent manner. Transl. Oncol. 2024, 45, 101956. [Google Scholar] [CrossRef]
- Guz, N.; Dokukin, M.; Kalaparthi, V.; Sokolov, I. If cell mechanics can be described by elastic modulus: Study of different models and probes used in indentation experiments. Biophys. J. 2014, 107, 564–575. [Google Scholar] [CrossRef]
- Bouchet, B.P.; Akhmanova, A. Microtubules in 3D cell motility. J. Cell Sci. 2017, 130, 39–50. [Google Scholar] [CrossRef]
- Wang, N.; Stamenovic, D. Contribution of intermediate filaments to cell stiffness, stiffening, and growth. Am. J. Physiol. Cell Physiol. 2000, 279, C188–C194. [Google Scholar] [CrossRef] [PubMed]
- Lehenkari, P.; Horton, M. Single integrin molecule adhesion forces in intact cells measured by atomic force microscopy. Biochem. Biophys. Res. Commun. 1999, 259, 645–650. [Google Scholar] [CrossRef]
- Simon, A.; Cohen-Bouhacina, T.; Porte, M.; Aime, J.; Amedee, J.; Bareille, R.; Baquey, C. Characterization of dynamic cellular adhesion of osteoblasts using atomic force microscopy. Cytom. Part A J. Int. Soc. Anal. Cytol. 2003, 54, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Puech, P.-H.; Poole, K.; Knebel, D.; Muller, D.J. A new technical approach to quantify cell–cell adhesion forces by AFM. Ultramicroscopy 2006, 106, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Pfeifer, T.; Gross, W.; De la Torre, C.; Zhao, S.; Liu, L.; Schaefer, M.; Herr, I. Establishment of tumor treating fields combined with mild hyperthermia as novel supporting therapy for pancreatic cancer. Front. Oncol. 2021, 11, 738801. [Google Scholar] [CrossRef]
- Karanam, N.K.; Story, M.D. An overview of potential novel mechanisms of action underlying Tumor Treating Fields-induced cancer cell death and their clinical implications. Int. J. Radiat. Biol. 2021, 97, 1044–1054. [Google Scholar] [CrossRef]
- Carrieri, F.A.; Smack, C.; Siddiqui, I.; Kleinberg, L.R.; Tran, P.T. Tumor treating fields: At the crossroads between physics and biology for cancer treatment. Front. Oncol. 2020, 10, 575992. [Google Scholar] [CrossRef]
- Bagley, S.; Mansour, M.; Pantel, A.; Schubert, E.; Dulal, C.; Nabavizadeh, A.; Mohan, S.; Maloney, E.; Prior, T.; Angeloni, N. P17. 04.B A phase 2 study of niraparib concomitant with tumor treating fields (ttfields) in patients with recurrent high-grade glioma (hgg). Neuro Oncol. 2023, 25, ii117. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkarni, T.; Banik, S.; Mukhopadhyay, D.; Babiker, H.; Bhattacharya, S. Tumor-Treating Fields Alter Nanomechanical Properties of Pancreatic Ductal Adenocarcinoma Cells Co-Cultured with Extracellular Matrix. J. Funct. Biomater. 2025, 16, 160. https://doi.org/10.3390/jfb16050160
Kulkarni T, Banik S, Mukhopadhyay D, Babiker H, Bhattacharya S. Tumor-Treating Fields Alter Nanomechanical Properties of Pancreatic Ductal Adenocarcinoma Cells Co-Cultured with Extracellular Matrix. Journal of Functional Biomaterials. 2025; 16(5):160. https://doi.org/10.3390/jfb16050160
Chicago/Turabian StyleKulkarni, Tanmay, Sreya Banik, Debabrata Mukhopadhyay, Hani Babiker, and Santanu Bhattacharya. 2025. "Tumor-Treating Fields Alter Nanomechanical Properties of Pancreatic Ductal Adenocarcinoma Cells Co-Cultured with Extracellular Matrix" Journal of Functional Biomaterials 16, no. 5: 160. https://doi.org/10.3390/jfb16050160
APA StyleKulkarni, T., Banik, S., Mukhopadhyay, D., Babiker, H., & Bhattacharya, S. (2025). Tumor-Treating Fields Alter Nanomechanical Properties of Pancreatic Ductal Adenocarcinoma Cells Co-Cultured with Extracellular Matrix. Journal of Functional Biomaterials, 16(5), 160. https://doi.org/10.3390/jfb16050160