

Reduction in Lens Epithelial Cell Senescence Burden through Dasatinib Plus Quercetin or Rapamycin Alleviates D-Galactose-Induced Cataract Progression

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Experimental Animals
2.2. Rat Model of D-Galactose-Induced Cataracts
2.3. Eye Drops Administration and Subconjunctival Injection
2.4. Opacity Index
2.5. Lens Density
2.6. Hematoxylin and Eosin Staining
2.7. Immunohistochemical Staining
2.8. Statistical Analysis
3. Results
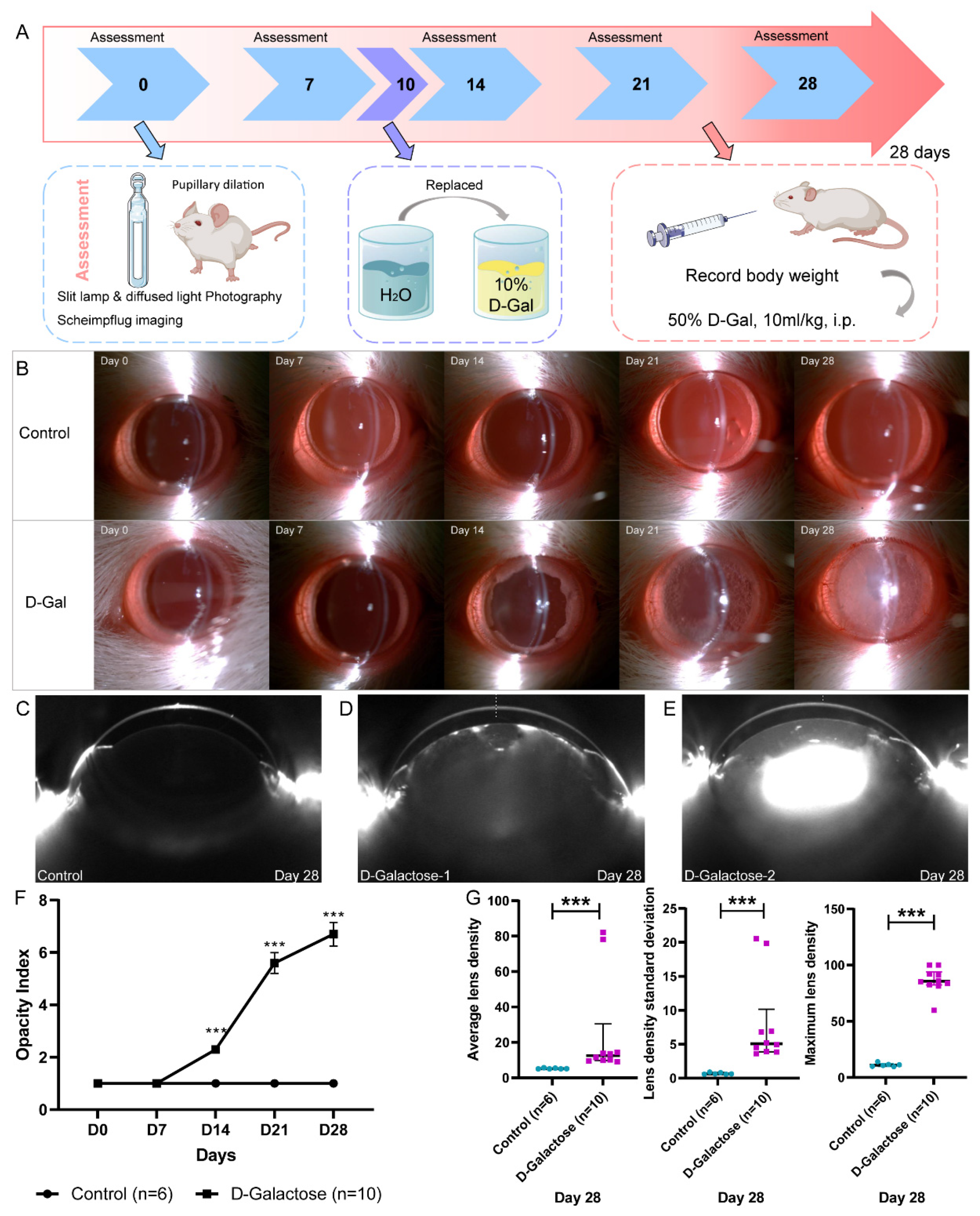
3.1. Establishment of the Rat Model of D-Galactose-Induced Cataracts
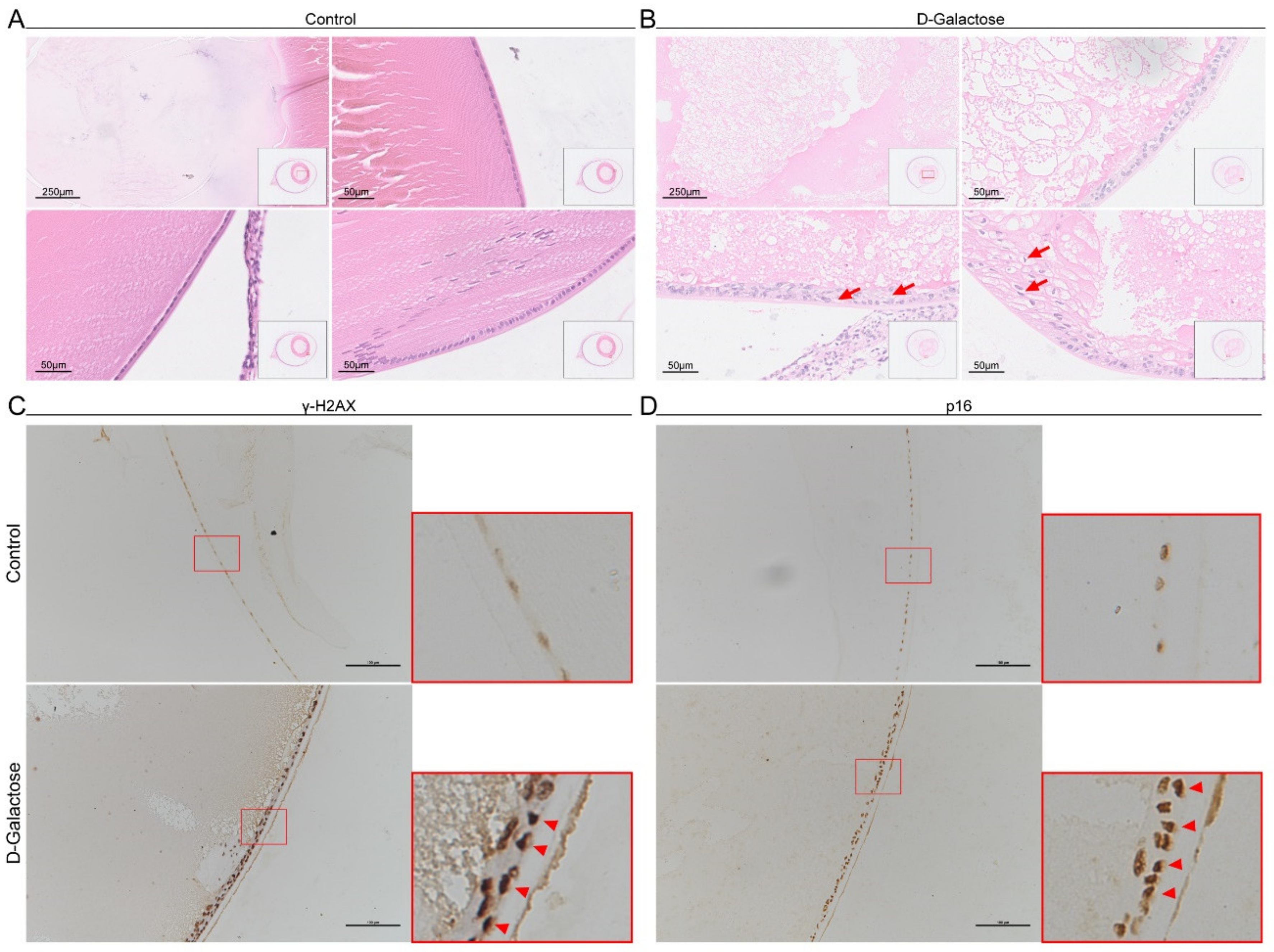
3.2. Senescent Lens Epithelial Cells Accumulate in D-Galactose-Induced Cataracts
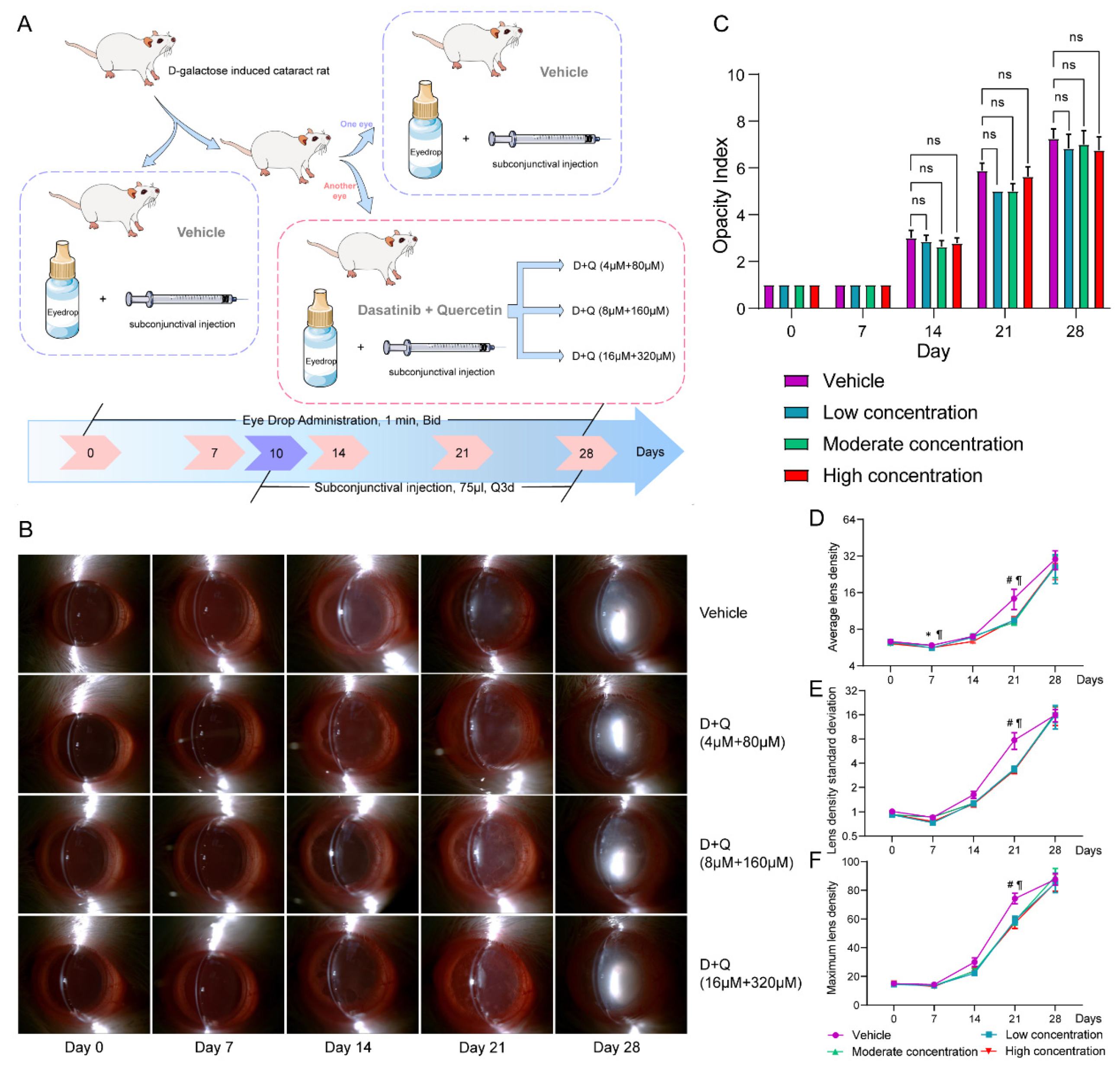
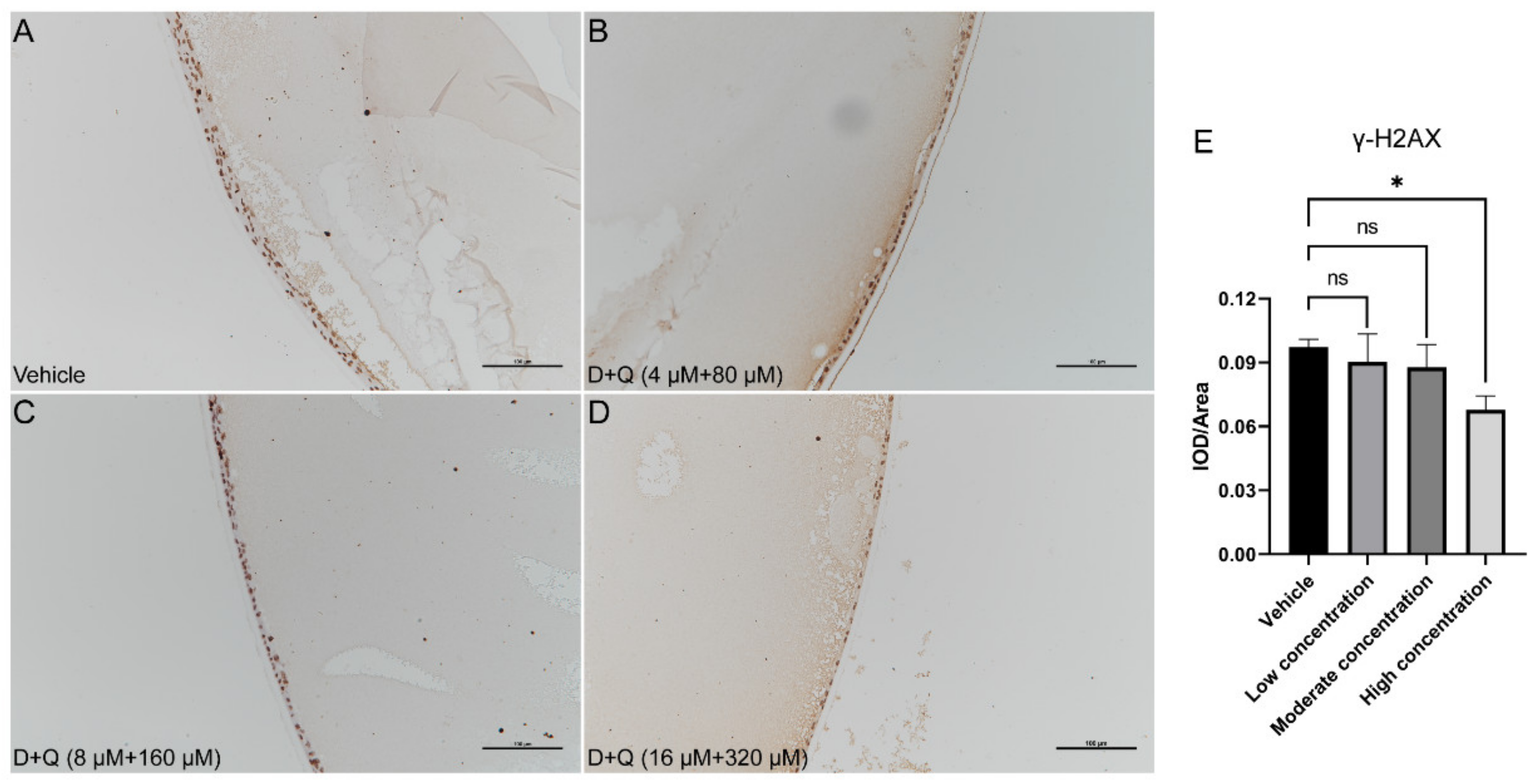
3.3. D+Q Treatment Tends to Alleviate the Progression of D-Galactose-Induced Cataracts in the Early Stage
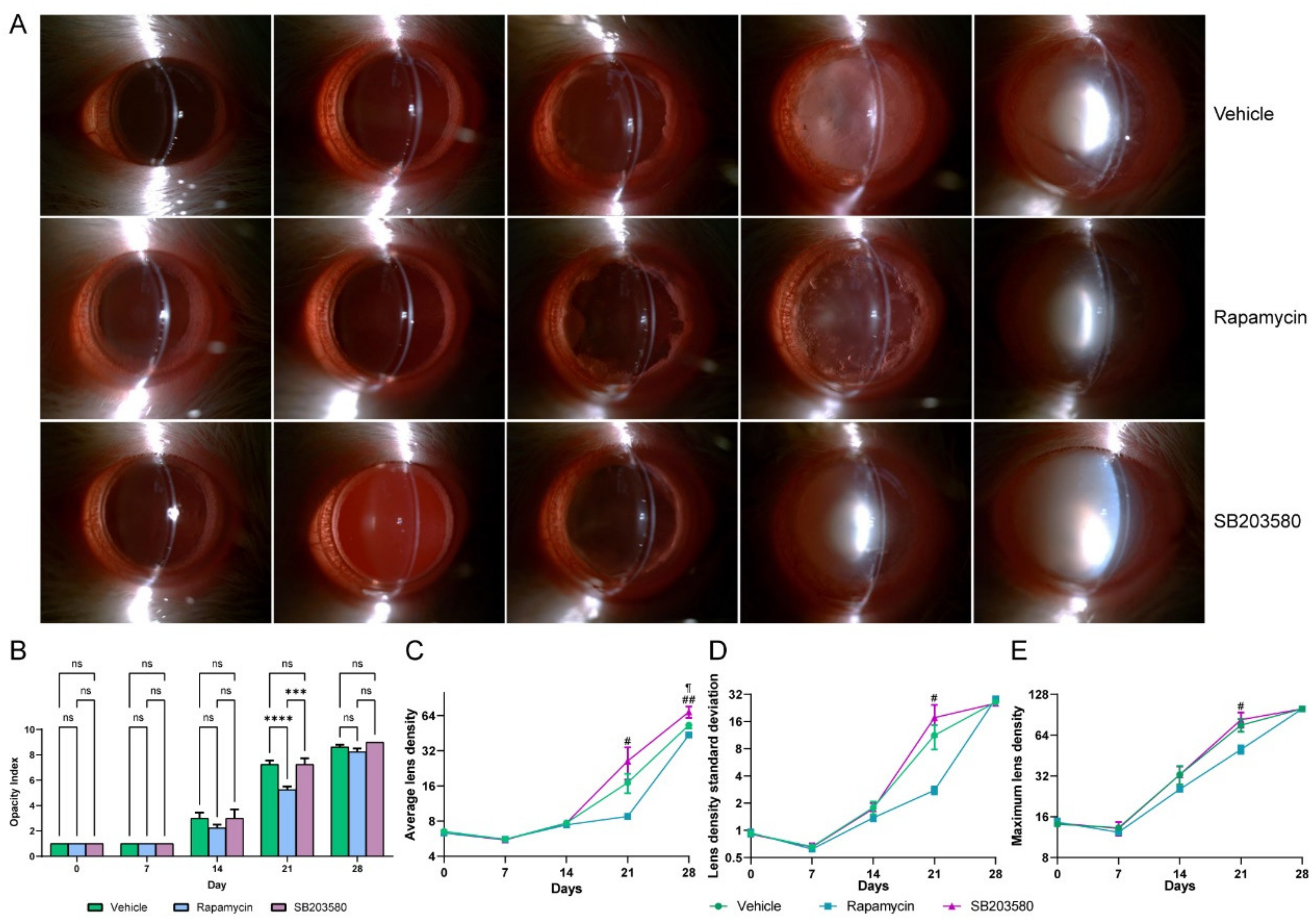
3.4. Rapamycin, a SASP Inhibitor, Tends to Alleviate the Progression of D-Galactose-Induced Cataracts in the Early Stage of Cataract
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALD | average lens density |
| D | dasatinib |
| H&E | hematoxylin and eosin |
| IHC | immunohistochemistry |
| LDSD | lens density standard deviation |
| LEC | lens epithelial cell |
| MLD | maximum lens density |
| Q | quercetin |
| SASP | senescence-associated secretory phenotype |
| γH2AX | gamma H2A.X |
References
- Munoz-Espin, D.; Serrano, M. Cellular senescence: From physiology to pathology. Nat. Rev. Mol. Cell Biol. 2014, 15, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Blindness and Vision Impairment Collaborators; Vision Loss Expert Group of the Global Burden of Disease Study. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: The Right to Sight: An analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e144–e160. [Google Scholar] [CrossRef] [PubMed]
- Wishart, T.F.L.; Flokis, M.; Shu, D.Y.; Das, S.J.; Lovicu, F.J. Hallmarks of lens aging and cataractogenesis. Exp. Eye Res. 2021, 210, 108709. [Google Scholar] [CrossRef]
- Fu, Q.; Qin, Z.; Yu, J.; Yu, Y.; Tang, Q.; Lyu, D.; Zhang, L.; Chen, Z.; Yao, K. Effects of senescent lens epithelial cells on the severity of age-related cortical cataract in humans: A case-control study. Medicine 2016, 95, e3869. [Google Scholar] [CrossRef]
- Yan, Y.; Yu, H.; Sun, L.; Liu, H.; Wang, C.; Wei, X.; Song, F.; Li, H.; Ge, H.; Qian, H.; et al. Laminin α4 overexpression in the anterior lens capsule may contribute to the senescence of human lens epithelial cells in age-related cataract. Aging 2019, 11, 2699–2723. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, C.; Zhou, N.; Wang, X.; Su, D.; Qi, Y. Metformin alleviates oxidative stress-induced senescence of human lens epithelial cells via AMPK activation and autophagic flux restoration. J. Cell. Mol. Med. 2021, 25, 8376–8389. [Google Scholar] [CrossRef]
- Zhong, L.; Wang, T.; Wang, T.; Chen, H.; Deng, J.; Ye, H.; Li, W.; Ling, S. Characterization of an i.p. D-galactose-induced cataract model in rats. . J. Pharmacol. Toxicol. Methods 2021, 107, 106891. [Google Scholar] [CrossRef]
- Feng, W.; Yang, X.; Feng, M.; Pan, H.; Liu, J.; Hu, Y.; Wang, S.; Zhang, D.; Ma, F.; Mao, Y. Alginate Oligosaccharide Prevents against D-galactose-mediated Cataract in C57BL/6J Mice via Regulating Oxidative Stress and Antioxidant System. Curr. Eye Res. 2021, 46, 802–810. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y.; Ma, L.; Xin, G.; Wei, Z.; Zeng, Z.; Xing, Z.; Li, S.; Niu, H.; Huang, W. d-galactose induces premature senescence of lens epithelial cells by disturbing autophagy flux and mitochondrial functions. Toxicol. Lett. 2018, 289, 99–106. [Google Scholar] [CrossRef]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; Lebrasseur, N.K.; Childs, B.G.; Van De Sluis, B.; Kirkland, J.L.; Van Deursen, J.M. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.M.; Jing, X.; Pike, L.A.; Zhou, Q.; Lim, D.-J.; Sams, S.B.; Lund, G.S.; Sharma, V.; Haugen, B.R.; Schweppe, R.E. Targeted inhibition of Src kinase with dasatinib blocks thyroid cancer growth and metastasis. Clin. Cancer Res. 2012, 18, 3580–3591. [Google Scholar] [CrossRef] [PubMed]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: The path to the clinic. Nat. Med. 2022, 28, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- Camell, C.D.; Yousefzadeh, M.J.; Zhu, Y.; Prata, L.G.P.L.; Huggins, M.A.; Pierson, M.; Zhang, L.; O’Kelly, R.D.; Pirtskhalava, T.; Xun, P.; et al. Senolytics reduce coronavirus-related mortality in old mice. Science 2021, 373, eabe4832. [Google Scholar] [CrossRef]
- Novais, E.J.; Tran, V.A.; Johnston, S.N.; Darris, K.R.; Roupas, A.J.; Sessions, G.A.; Shapiro, I.M.; Diekman, B.O.; Risbud, M.V. Long-term treatment with senolytic drugs Dasatinib and Quercetin ameliorates age-dependent intervertebral disc degeneration in mice. Nat. Commun. 2021, 12, 5213. [Google Scholar] [CrossRef]
- Saccon, T.D.; Nagpal, R.; Yadav, H.; Cavalcante, M.B.; Nunes, A.D.D.C.; Schneider, A.; Gesing, A.; Hughes, B.; Yousefzadeh, M.; Tchkonia, T.; et al. Senolytic Combination of Dasatinib and Quercetin Alleviates Intestinal Senescence and Inflammation and Modulates the Gut Microbiome in Aged Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 1895–1905. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Wang, S. The Role of Rapamycin in Healthspan Extension via the Delay of Organ Aging. Ageing Res. Rev. 2021, 70, 101376. [Google Scholar] [CrossRef]
- Abdul Nasir, N.A.; Agarwal, R.; Vasudevan, S.; Tripathy, M.; Alyautdin, R.; Ismail, N.M. Effects of topically applied tocotrienol on cataractogenesis and lens redox status in galactosemic rats. Mol. Vis. 2014, 20, 822–835. [Google Scholar]
- Liu, J.; Zhang, J.; Zhang, G.; Zou, X.; Guan, H.; Wang, Y. CircMRE11A_013 binds to UBXN1 and integrates ATM activation enhancing lens epithelial cells senescence in age-related cataract. Aging 2021, 13, 5383–5402. [Google Scholar] [CrossRef] [PubMed]
- Birch, J.; Gil, J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Du, C.; Wan, S.; Wu, D.; Yan, L.; Zhang, J.; Li, J.; Li, H.; Yang, Z.; Zhang, H.; et al. Deficiency of heat shock factor 4 promotes lens epithelial cell senescence through upregulating p21(cip1) expression. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166233. [Google Scholar] [CrossRef]
- Matsuyama, M.; Tanaka, H.; Inoko, A.; Goto, H.; Yonemura, S.; Kobori, K.; Hayashi, Y.; Kondo, E.; Itohara, S.; Izawa, I.; et al. Defect of mitotic vimentin phosphorylation causes microophthalmia and cataract via aneuploidy and senescence in lens epithelial cells. J. Biol. Chem. 2013, 288, 35626–35635. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Kim, S.R.; Puranik, A.S.; Jiang, K.; Chen, X.; Zhu, X.-Y.; Taylor, I.; Khodadadi-Jamayran, A.; Lerman, A.; Hickson, L.J.; Childs, B.G.; et al. Progressive Cellular Senescence Mediates Renal Dysfunction in Ischemic Nephropathy. J. Am. Soc. Nephrol. 2021, 32, 1987–2004. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Gasek, N.S.; Zhou, Y.; Cohn, R.L.; Martin, D.E.; Zuo, W.; Flynn, W.F.; Guo, C.; Jellison, E.R.; et al. Targeting p21(Cip1) highly expressing cells in adipose tissue alleviates insulin resistance in obesity. Cell Metab. 2022, 34, 75–89. [Google Scholar] [CrossRef]
- Ping, X.; Liang, J.; Shi, K.; Bao, J.; Wu, J.; Yu, X.; Tang, X.; Zou, J.; Shentu, X. Rapamycin relieves the cataract caused by ablation of Gja8b through stimulating autophagy in zebrafish. Autophagy 2021, 17, 3323–3337. [Google Scholar] [CrossRef]
- Wilkinson, J.E.; Burmeister, L.; Brooks, S.V.; Chan, C.-C.; Friedline, S.; Harrison, D.E.; Hejtmancik, J.F.; Nadon, N.; Strong, R.; Wood, L.K.; et al. Rapamycin slows aging in mice. Aging Cell 2012, 11, 675–682. [Google Scholar] [CrossRef]
- Gong, W.; Zhu, G.; Li, J.; Yang, X. LncRNA MALAT1 promotes the apoptosis and oxidative stress of human lens epithelial cells via p38MAPK pathway in diabetic cataract. Diabetes Res. Clin. Pract. 2018, 144, 314–321. [Google Scholar] [CrossRef]
- Bai, J.; Zheng, Y.; Dong, L.; Cai, X.; Wang, G.; Liu, P. Inhibition of p38 mitogen-activated protein kinase phosphorylation decreases H(2)O(2)-induced apoptosis in human lens epithelial cells. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 1933–1940. [Google Scholar] [CrossRef] [PubMed]
- Makhotkina, N.Y.; Berendschot, T.; van den Biggelaar, F.; Weik, A.R.H.; Nuijts, R.M.M.A. Comparability of subjective and objective measurements of nuclear density in cataract patients. Acta Ophthalmol. 2018, 96, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Kunze, S.; Cecil, A.; Prehn, C.; Möller, G.; Ohlmann, A.; Wildner, G.; Thurau, S.; Unger, K.; Rößler, U.; Hölter, S.M.; et al. Posterior subcapsular cataracts are a late effect after acute exposure to 0.5 Gy ionizing radiation in mice. Int. J. Radiat. Biol. 2021, 97, 529–540. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Tseng, Y.; Chen, K.; Wang, X.; Mao, Z.; Li, X. Reduction in Lens Epithelial Cell Senescence Burden through Dasatinib Plus Quercetin or Rapamycin Alleviates D-Galactose-Induced Cataract Progression. J. Funct. Biomater. 2023, 14, 6. https://doi.org/10.3390/jfb14010006
Wang Y, Tseng Y, Chen K, Wang X, Mao Z, Li X. Reduction in Lens Epithelial Cell Senescence Burden through Dasatinib Plus Quercetin or Rapamycin Alleviates D-Galactose-Induced Cataract Progression. Journal of Functional Biomaterials. 2023; 14(1):6. https://doi.org/10.3390/jfb14010006
Chicago/Turabian StyleWang, Yinhao, Yulin Tseng, Keyu Chen, Xinglin Wang, Zebin Mao, and Xuemin Li. 2023. "Reduction in Lens Epithelial Cell Senescence Burden through Dasatinib Plus Quercetin or Rapamycin Alleviates D-Galactose-Induced Cataract Progression" Journal of Functional Biomaterials 14, no. 1: 6. https://doi.org/10.3390/jfb14010006
APA StyleWang, Y., Tseng, Y., Chen, K., Wang, X., Mao, Z., & Li, X. (2023). Reduction in Lens Epithelial Cell Senescence Burden through Dasatinib Plus Quercetin or Rapamycin Alleviates D-Galactose-Induced Cataract Progression. Journal of Functional Biomaterials, 14(1), 6. https://doi.org/10.3390/jfb14010006