Carbonate Apatite and Hydroxyapatite Formulated with Minimal Ingredients to Deliver SiRNA into Breast Cancer Cells In Vitro and In Vivo

 , , and
, , and
Abstract
1. Introduction
2. Methods and Materials
2.1. Reagents
2.2. Preparation of NPs
2.3. Assessment of Particle Growth Via Turbidity Measurement
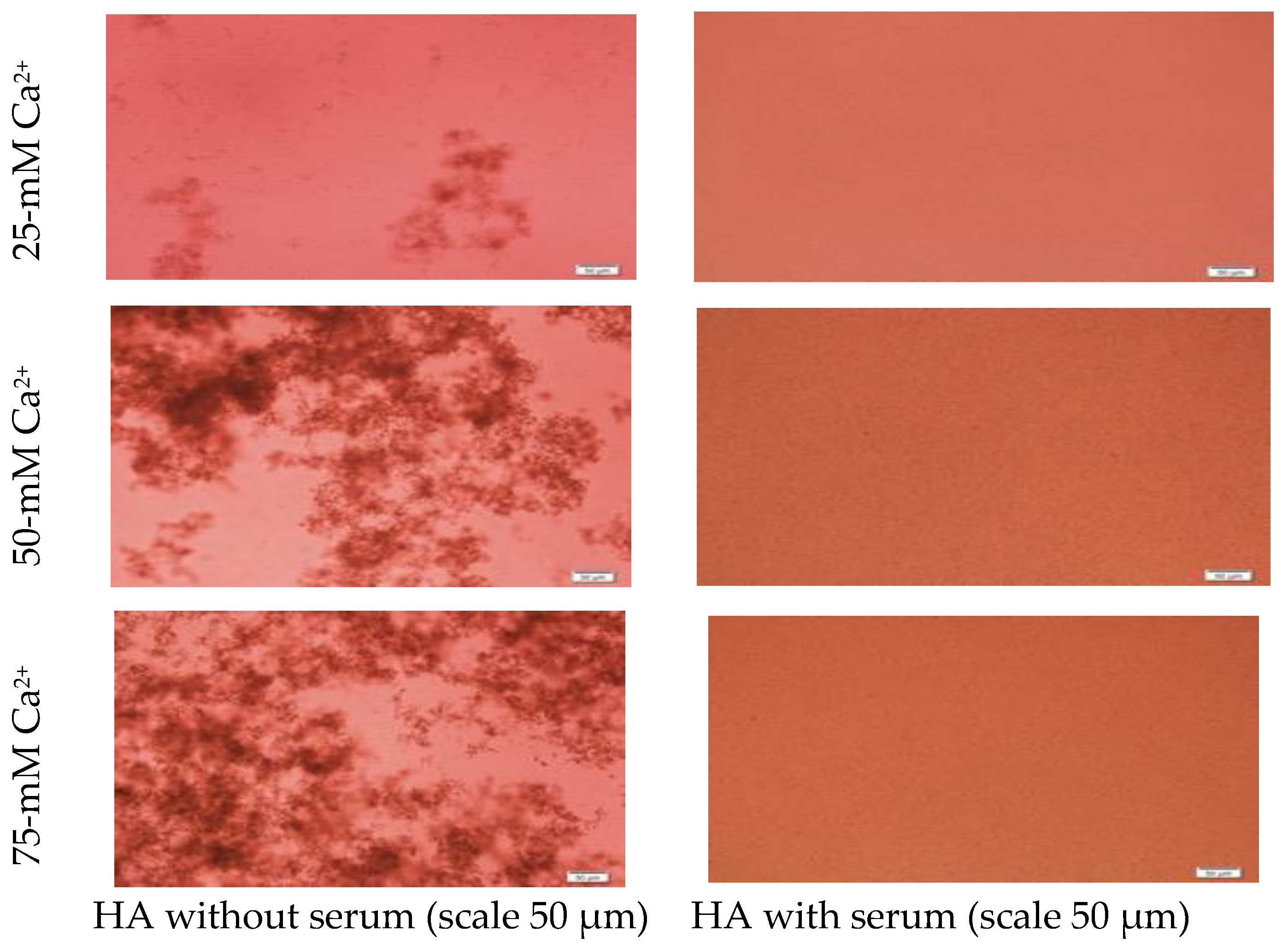
2.4. Microscopic Observation of NPs
2.5. Size Measurement by Dynamic Light Scattering
2.6. Size Measurement with Field Emission Scanning Electron Microscope (FE-SEM)
2.7. Attenuated Total Reflection-Fourier Transform Infrared Spectroscopy (ATR-FTIR)
2.8. pH Sensitivity of the NPs
2.9. Measurement of siRNA-Binding Efficiency
2.10. Stability Analysis for NPs
2.11. Cell Culture and Seeding
2.12. Cell Viability Assessment with 3-(4,5-Dimethylthiazol-2-yl)-2,5 Diphenyltetrazolium Bromide (MTT) Assay
2.13. Microscopic Observation of Cellular Uptake of NPs Bound Fluorescent SiRNA
2.14. Protein Corona Identification via LC–MS/MS
2.14.1. Sample Preparation for Protein Corona (PC) Analysis
2.14.2. In-Solution Tryptic Digestion of Protein
2.14.3. Sample Preparation for Q-TOF Mass Spectrometry
2.14.4. Nanoflow Liquid Chromatography Electrospray—Ionization Coupled with LC–MS/MS
2.14.5. Protein Identification by Automated De Novo Sequencing (PEAKS Studio 8.0)
2.15. Biodistribution of NPs in a Mouse Model of Breast Cancer
2.16. Statistical Analysis
3. Results
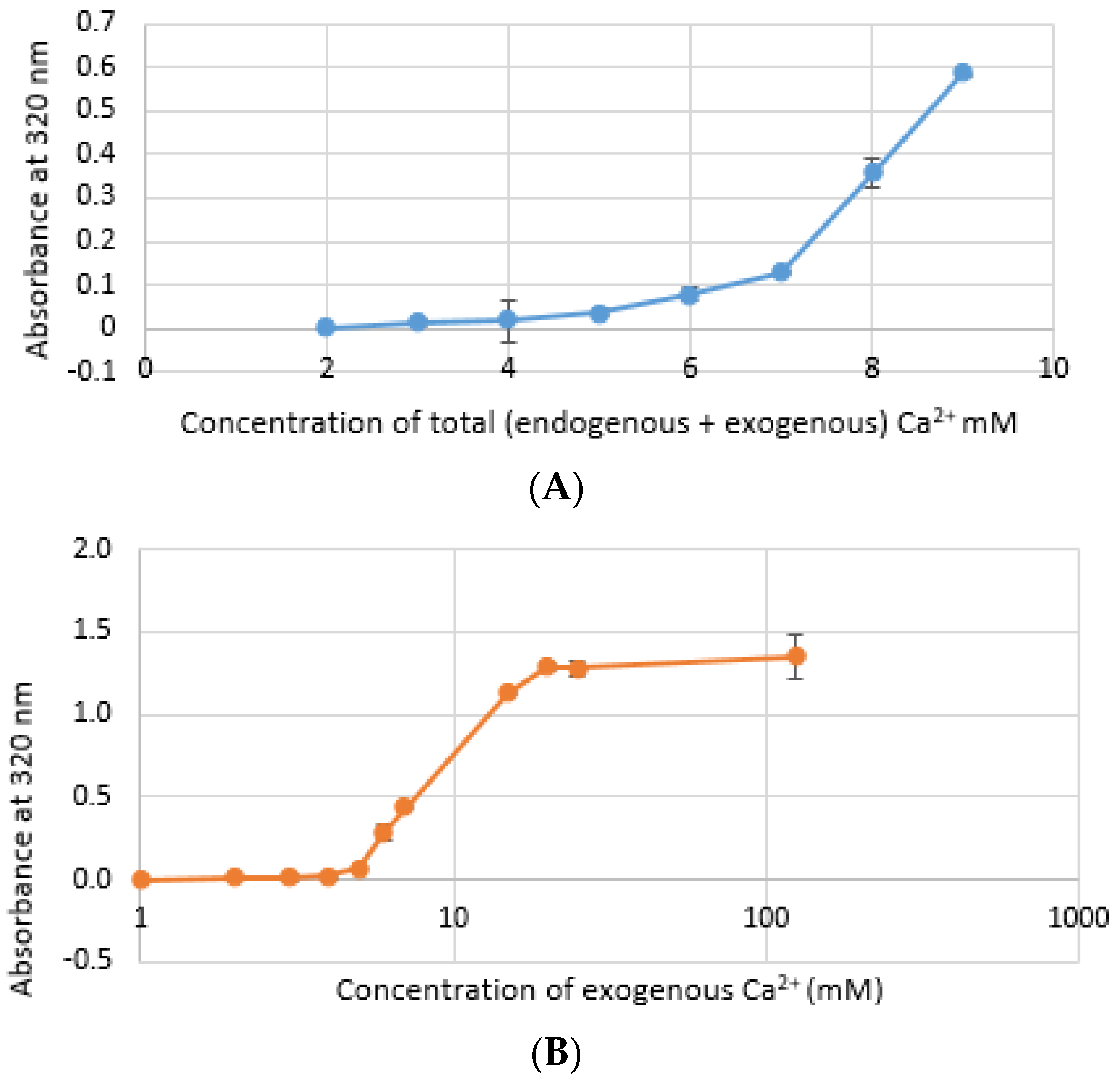
3.1. Pattern of Particle Growth for CA-in-DMEM and CA-in-DMB and HA
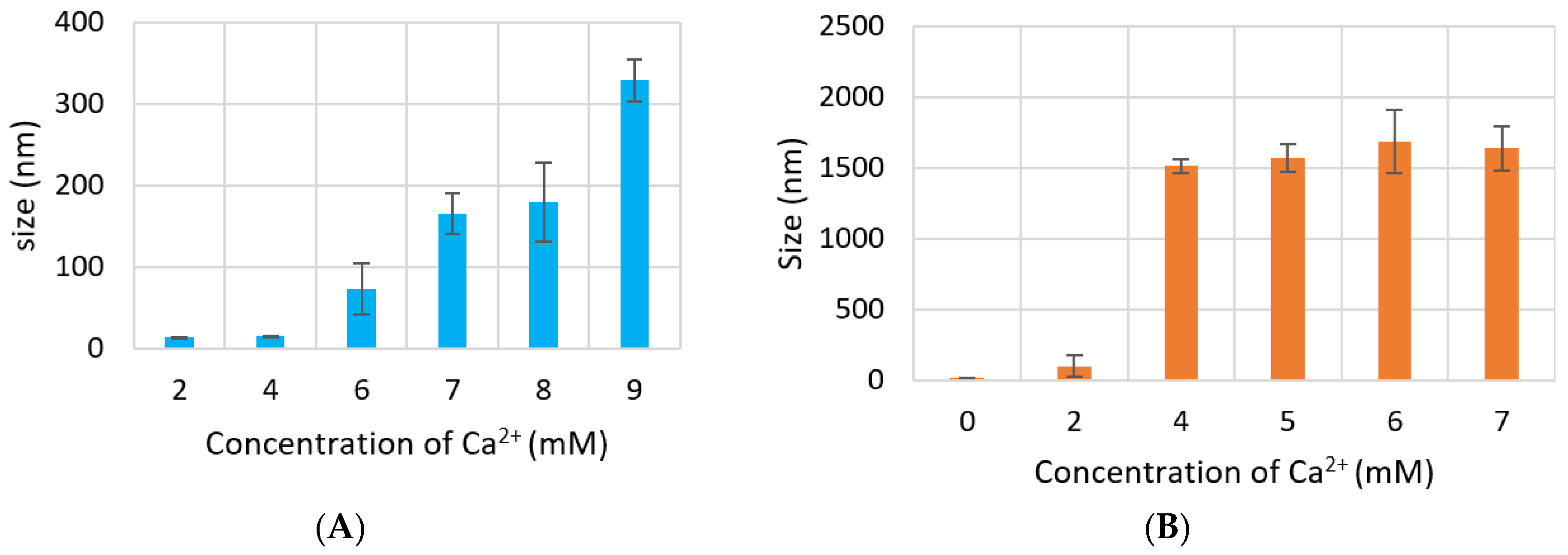
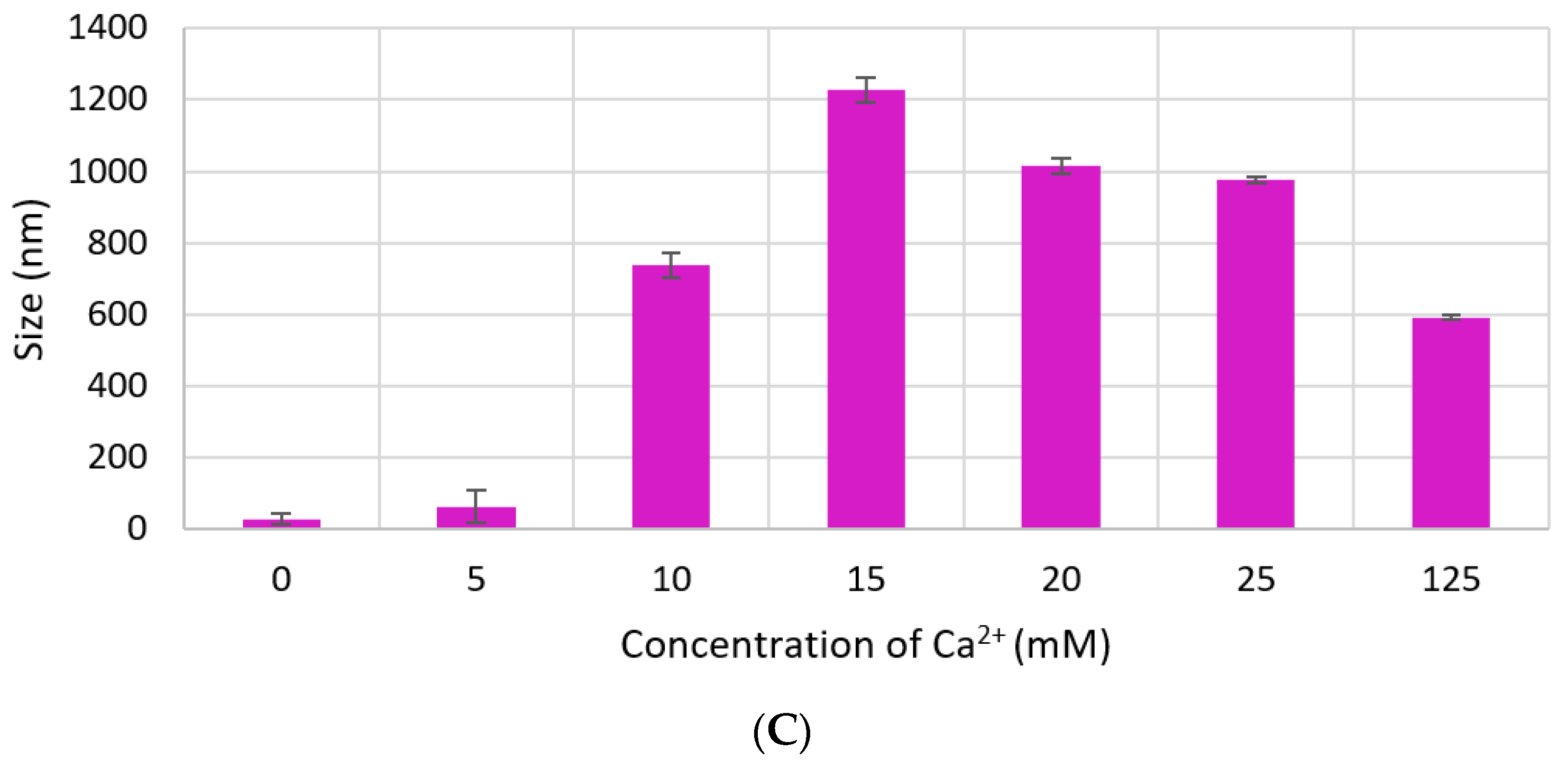
3.2. Measurement of Particle Size by Dynamic Light Scattering and FE-SEM
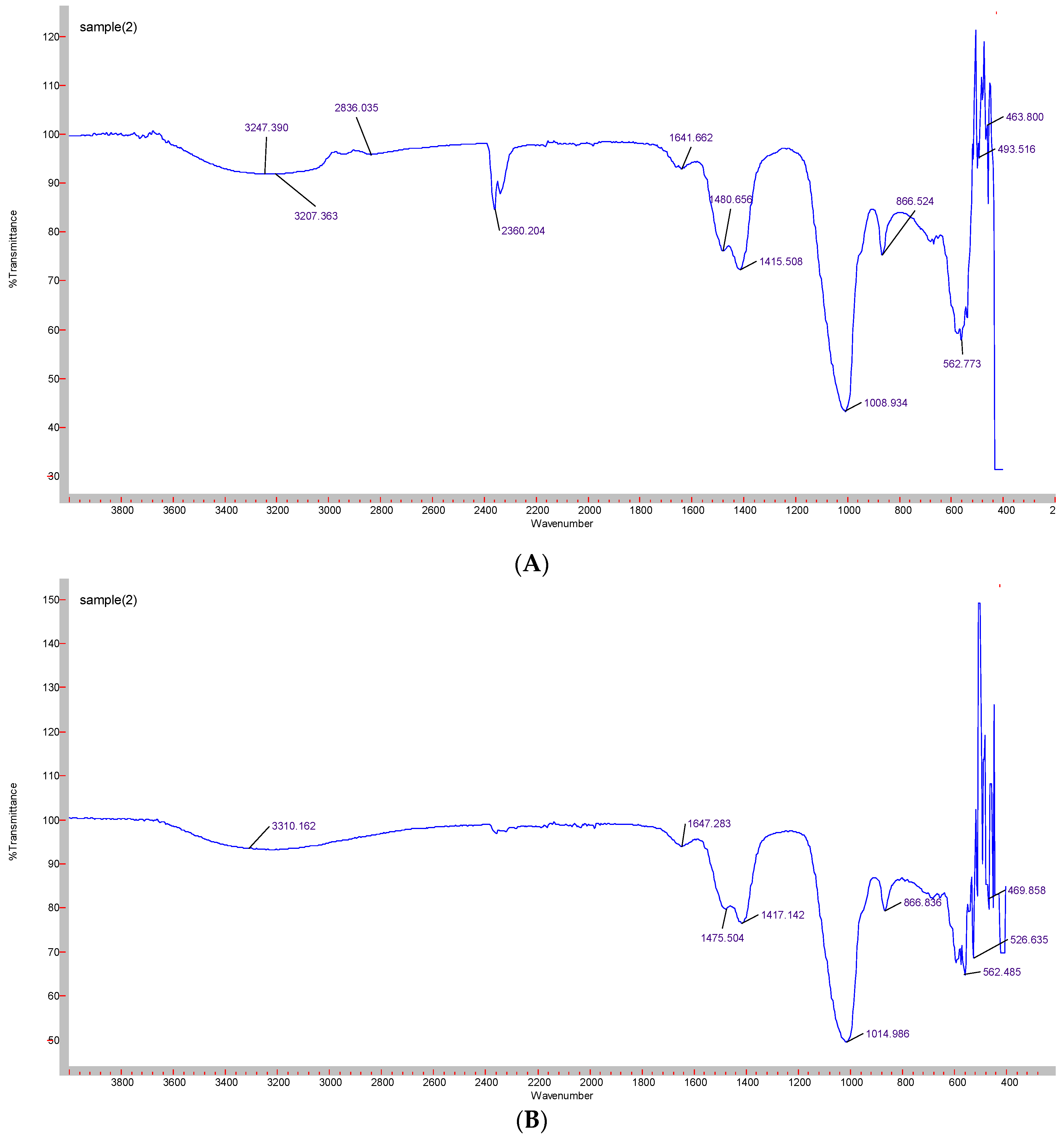
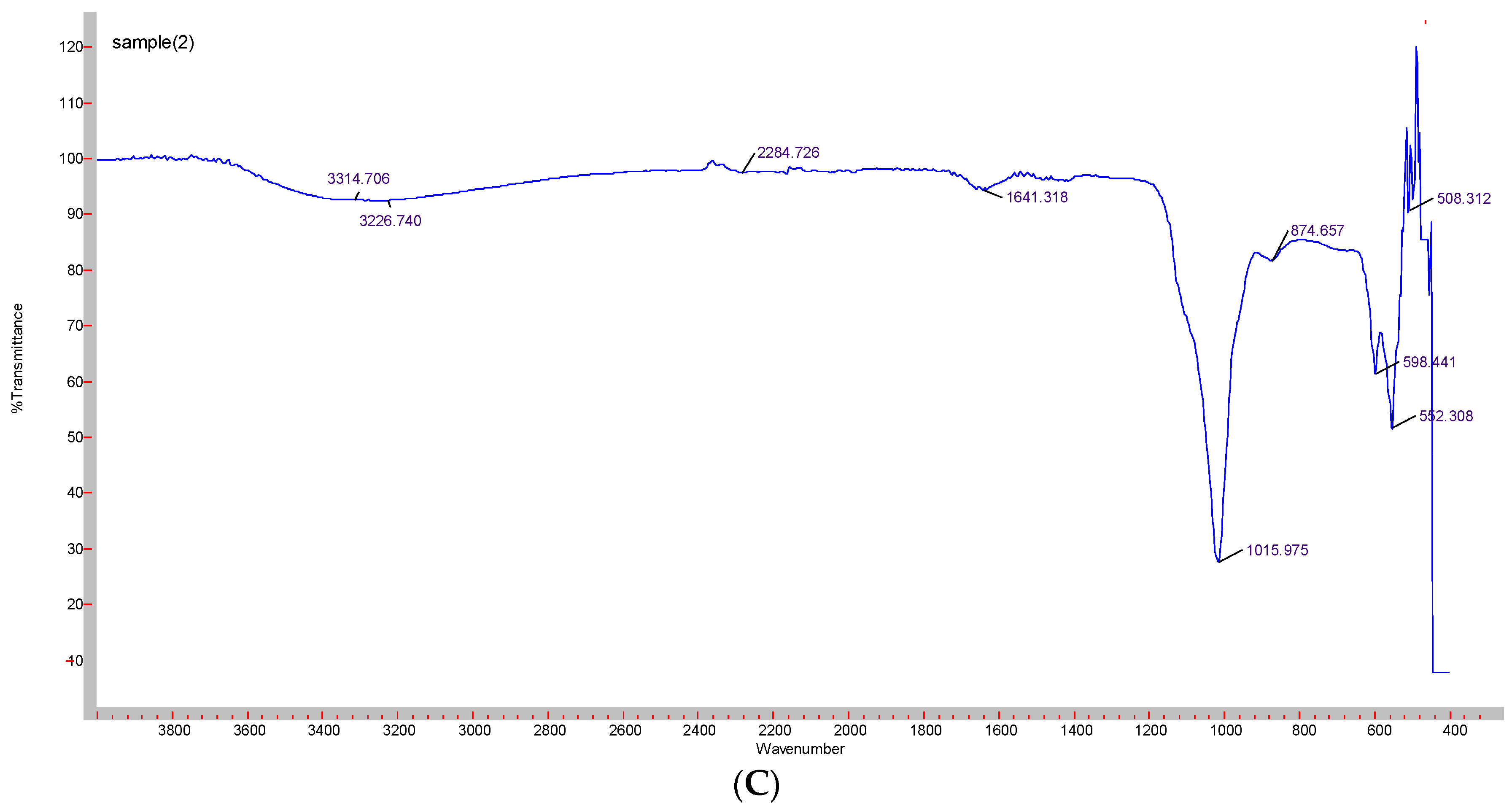
3.3. Confirmation of Functional Groups by ATR-FTIR
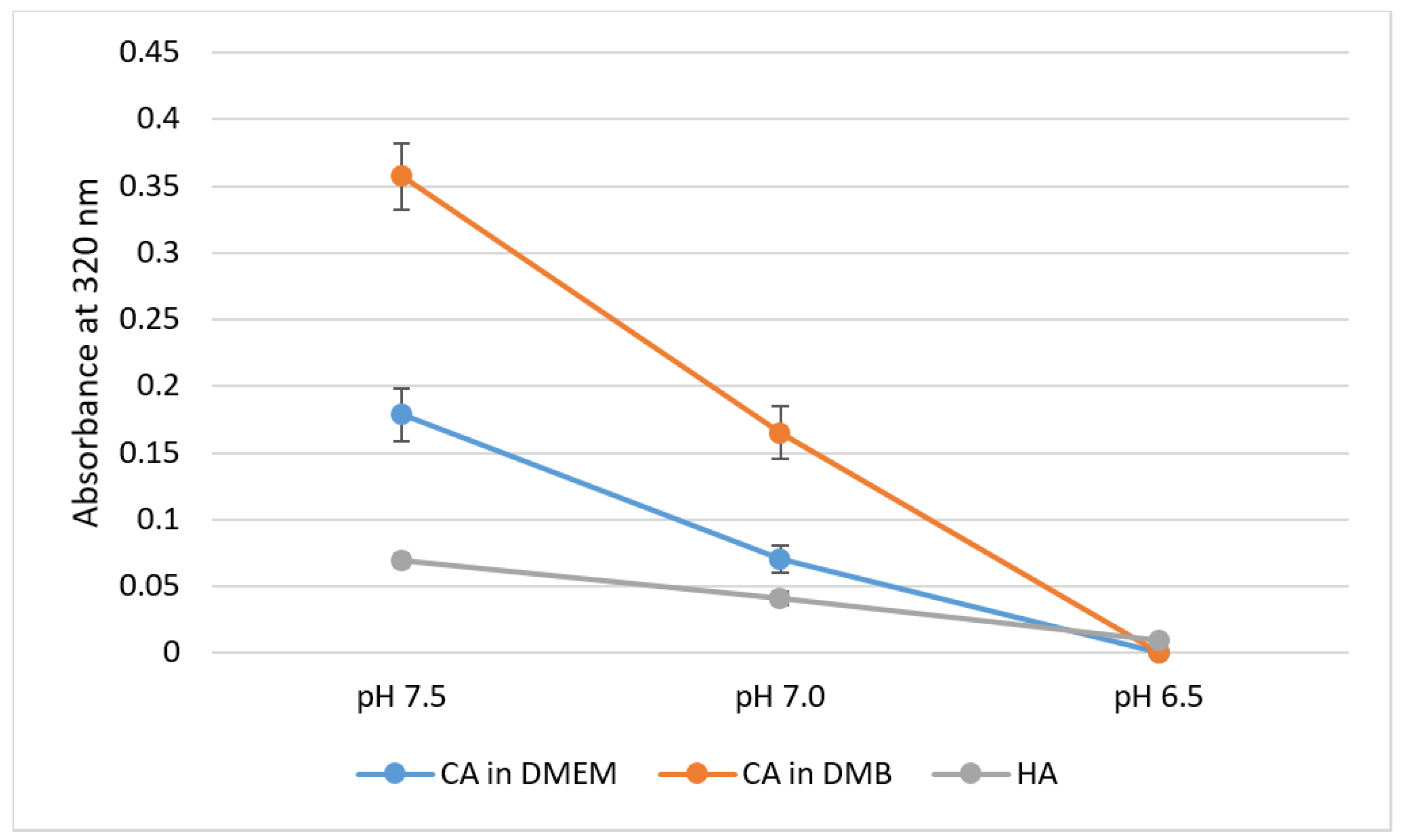
3.4. pH Sensitivity of NPs
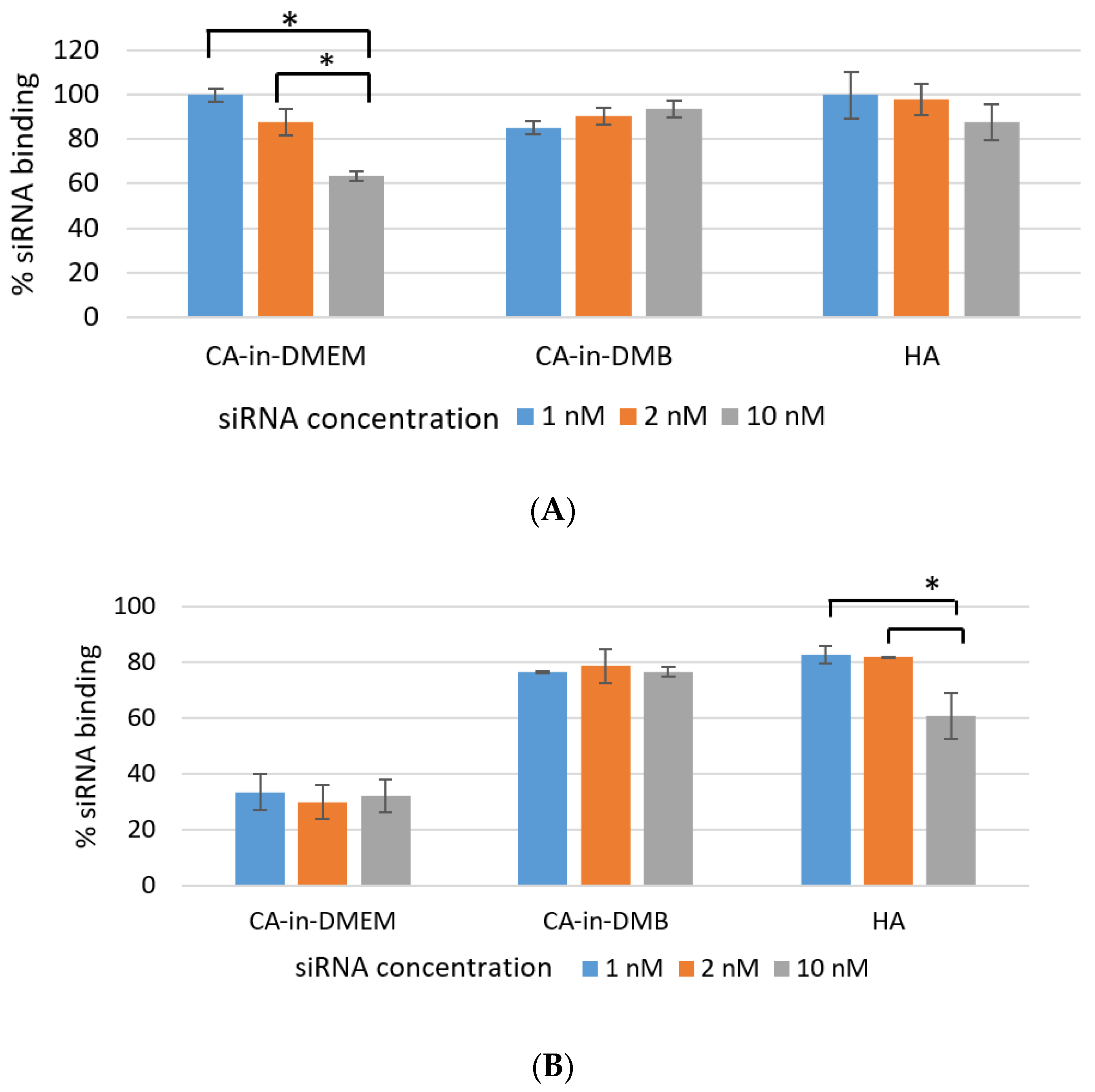
3.5. SiRNA-Binding Efficiency
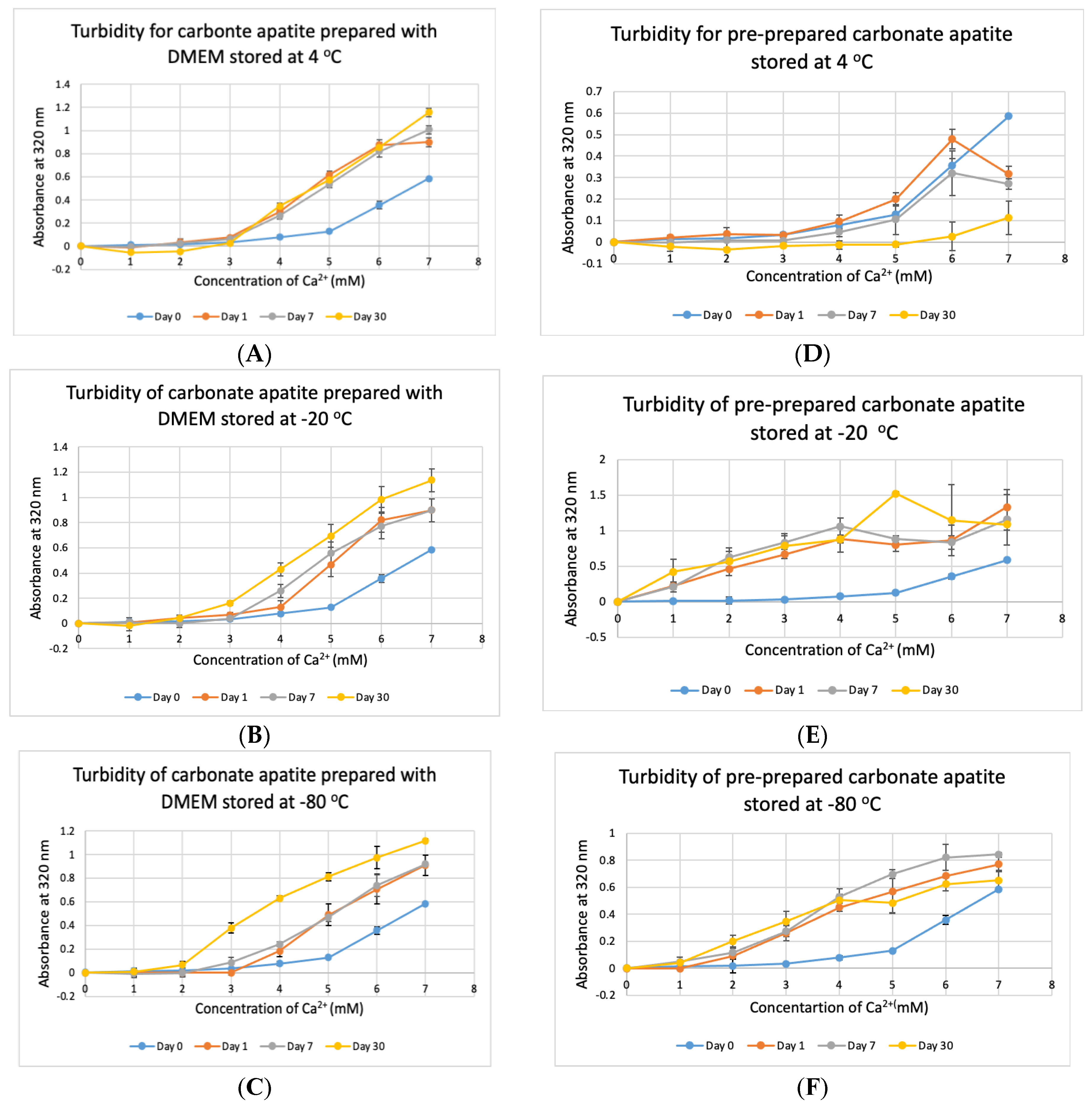
3.6. Stability Test for DMEM and CA Prepared in DMEM
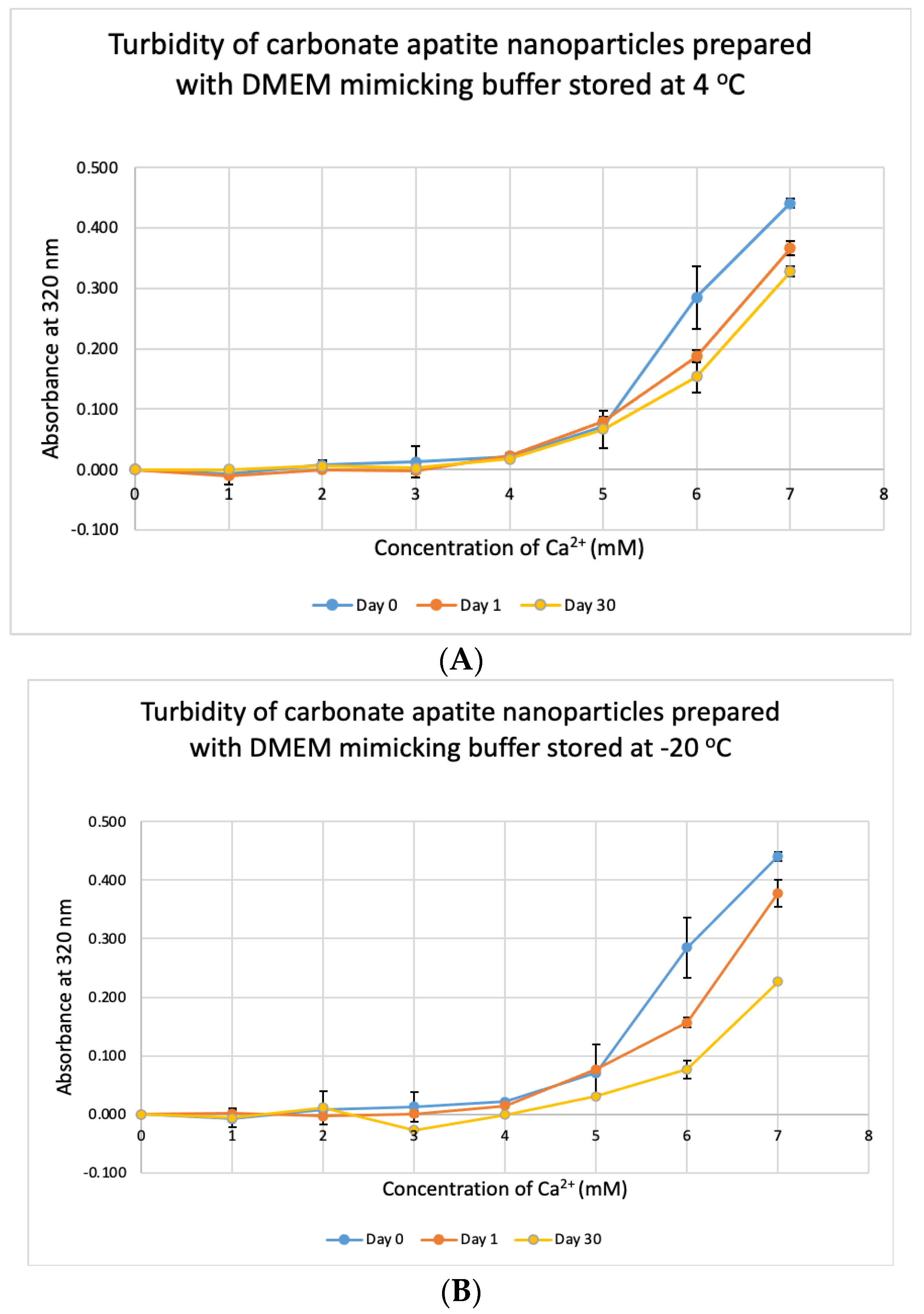
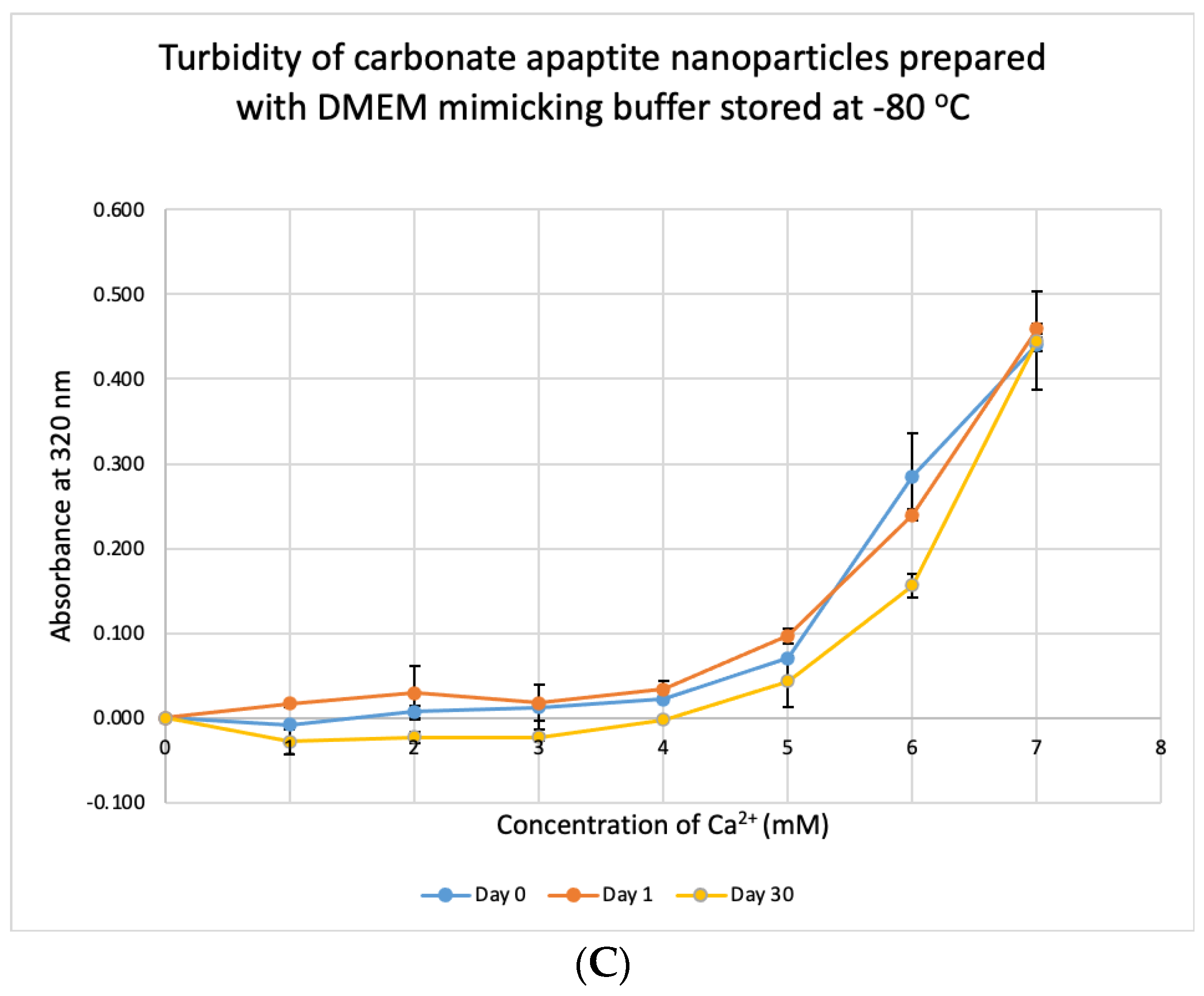
3.7. Stability Test for DMB
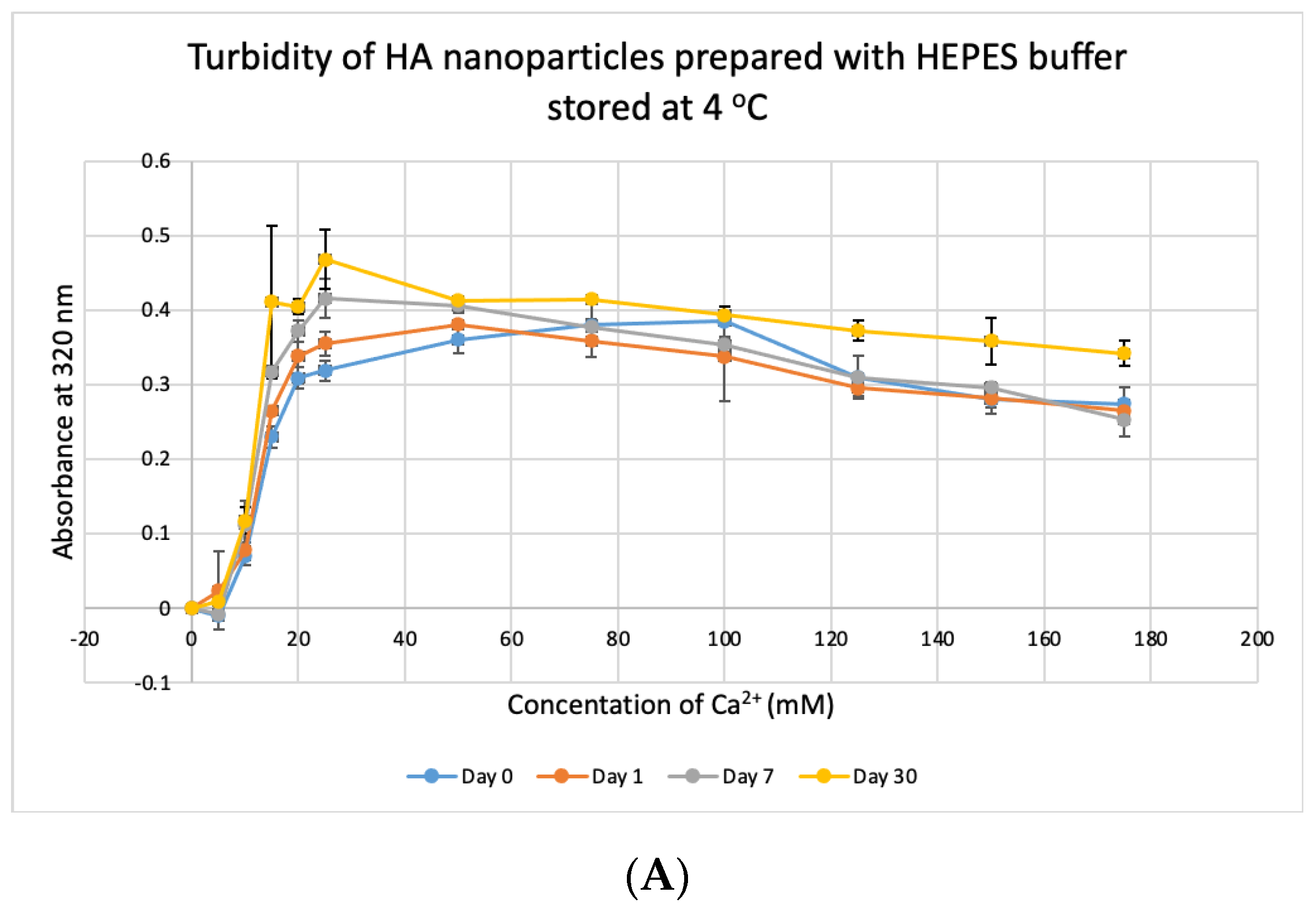
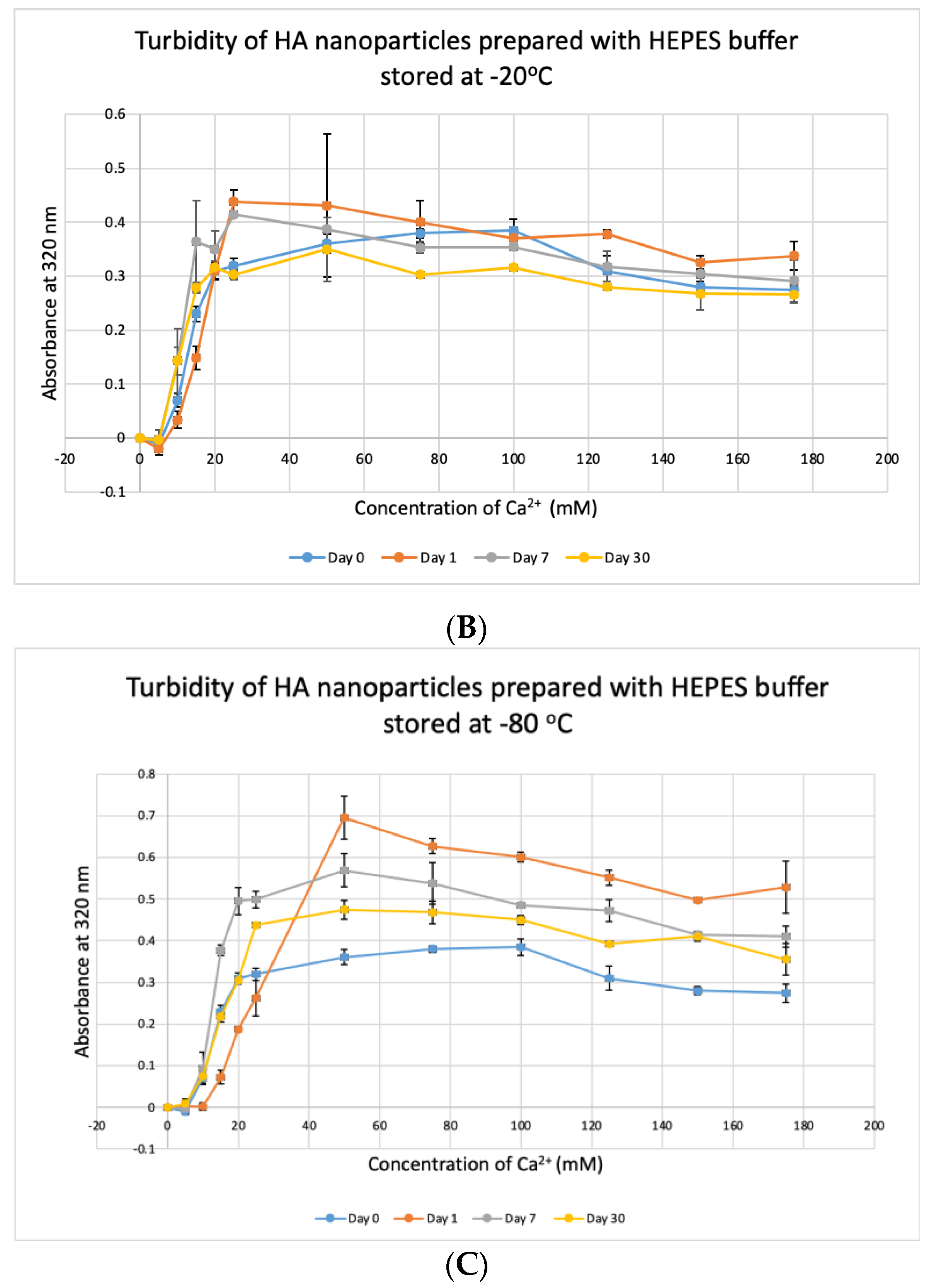
3.8. Stability Test for HBS
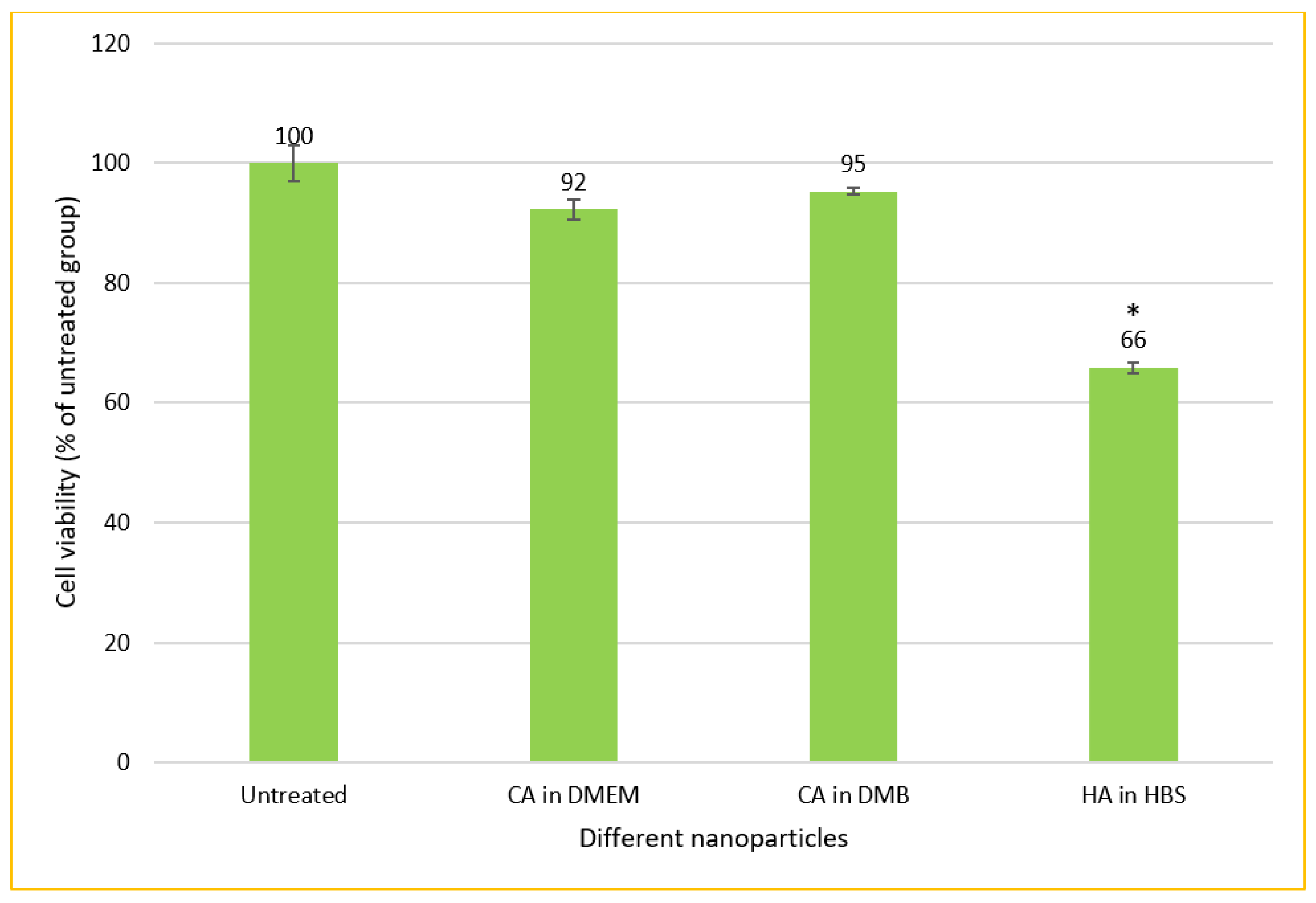
3.9. Cytotoxicity of CA-in-DMEM, CA-in-DMB and HA
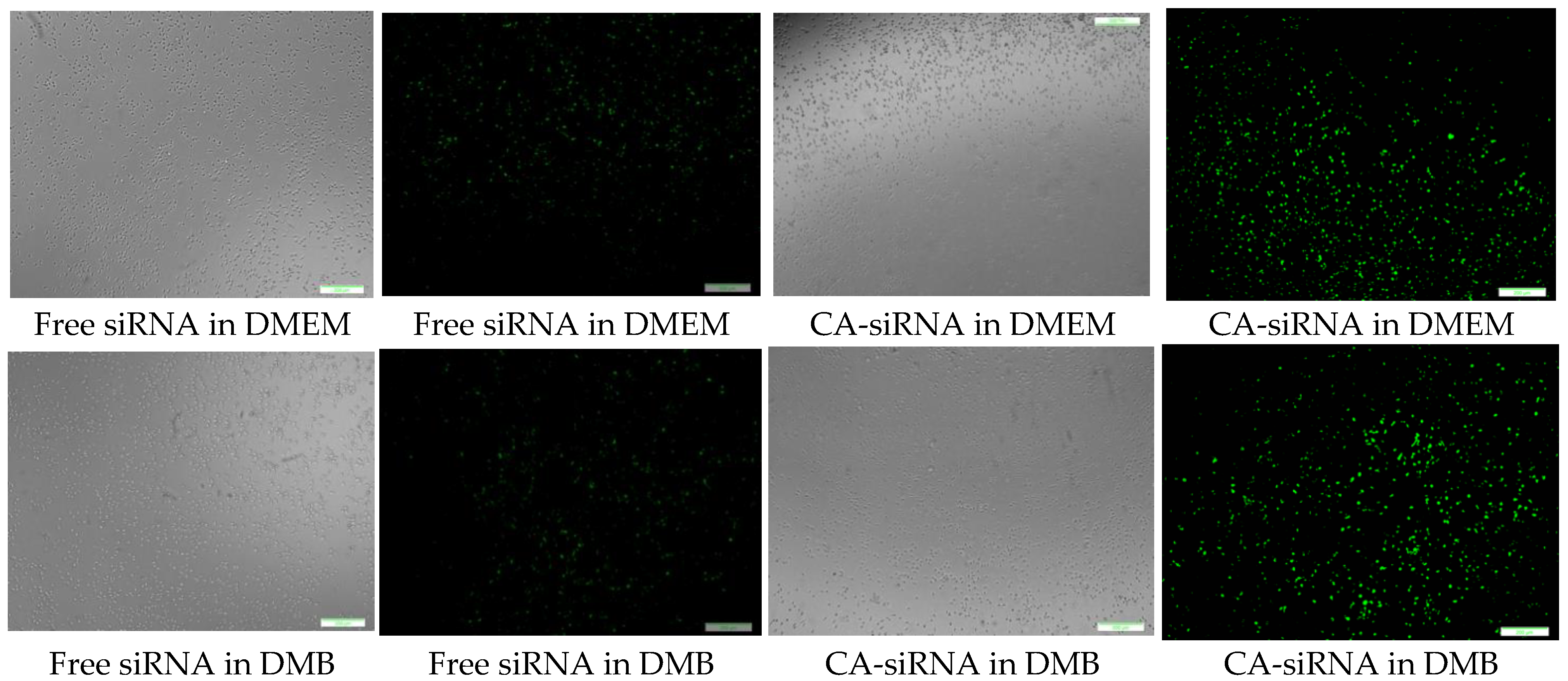
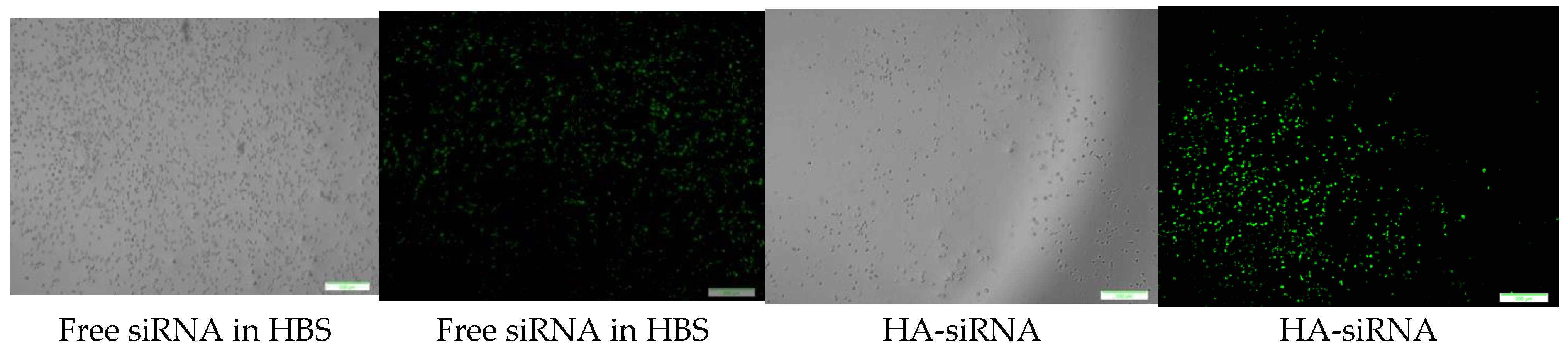
3.10. Cellular Uptake of Fluorescence siRNA-Loaded CA and HA—Microscopic Analysis
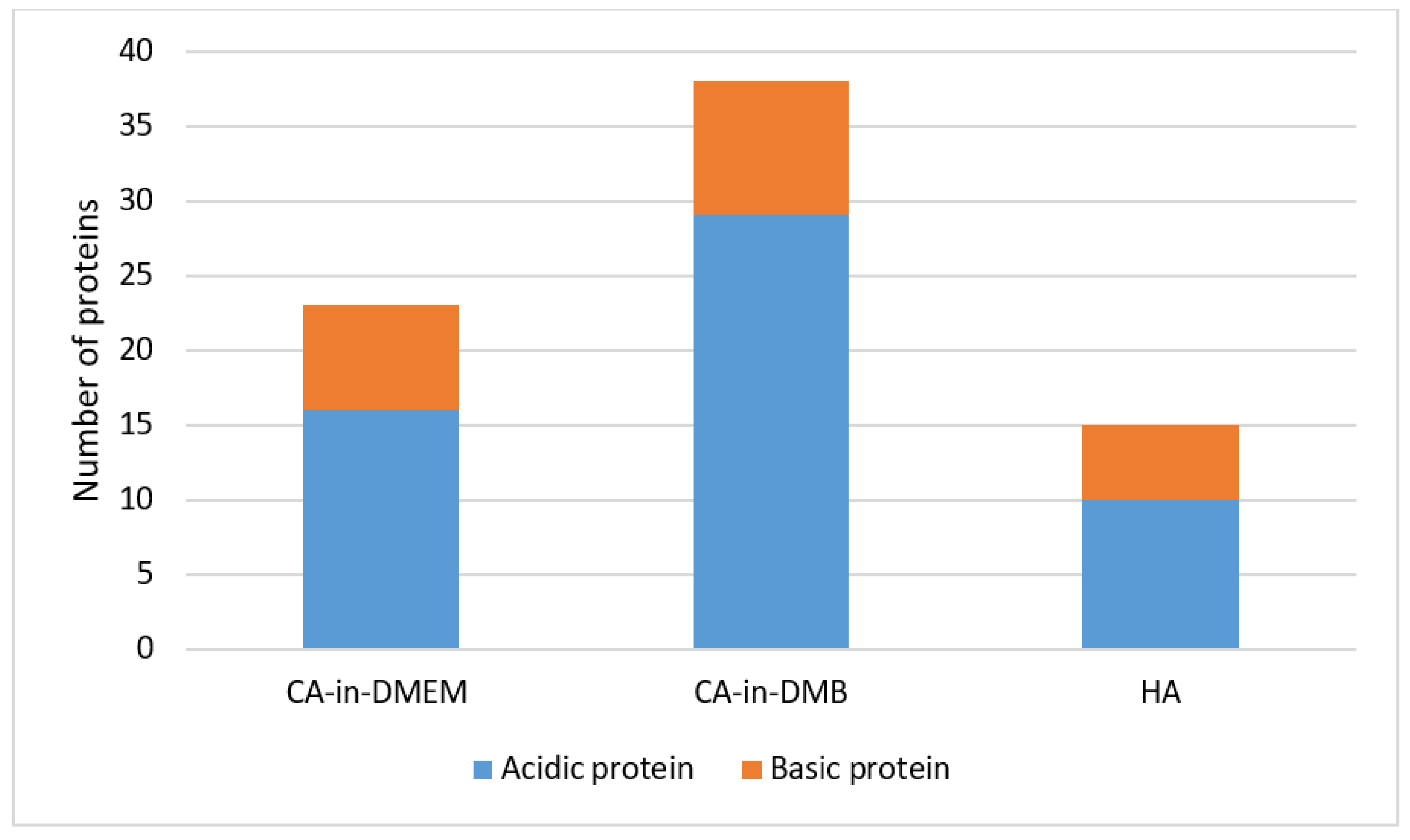
3.11. Identification of Spontaneously Formed PC on NPs, Via LC–MS
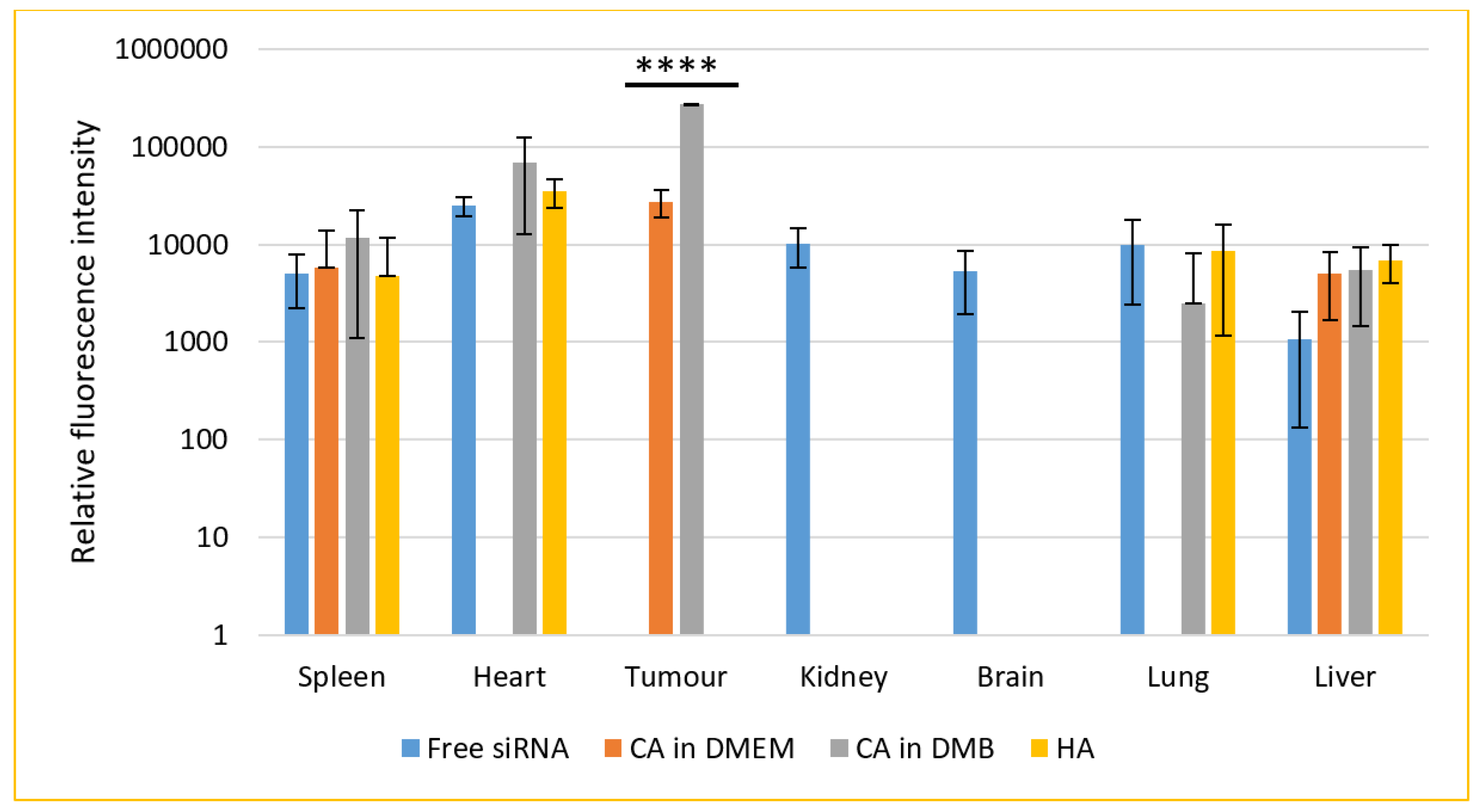
3.12. Biodistribution of NPs in a Mouse Model of Breast Cancer
4. Discussion
4.1. Pattern of Particle Growth, Size and SiRNA-Binding
4.2. Stability of the Buffers and CA-in-DMEM
4.3. Potential Roles of PC Components
4.4. NPs-Mediated SiRNA Delivery In Vitro and In Vivo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NP | Nanoparticles |
| PC | Protein corona |
| HA | Hydroxyapatite |
| CA | Carbonate apatite |
| DMB | DMEM-mimicking buffer |
| HBS | HEPES-buffered solution |
References
- Nemunaitis, J.; Senzer, N.; Khalil, I.; Shen, Y.; Kumar, P.; Tong, A.; Kuhn, J.; Lamont, J.; Rao, D.; Zhang, Y.-A.; et al. Proof concept for clinical justification of network mapping for personalized cancer therapeutics. Cancer Gene Ther. 2007, 14, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.A. Gene-Based Therapies for Cancer, 1st ed.; Springer: New York, NY, USA, 2010. [Google Scholar]
- Li, L.; Shen, Y. Overcoming obstacles to develop effective and safe siRNA therapeutics. Expert Opin. Biol. Ther. 2009, 9, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, E.; Kunou, M.; Nagaoka, M.; Kundu, A.; Hoshiba, T.; Akaike, T. High-efficiency gene delivery for expression in mammalian cells by nanoprecipitates of Ca–Mg phosphate. Gene 2004, 341, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Bochicchio, S.; Dalmoro, A.; Barba, A.A.; Grassi, G.; Lamberti, G. Liposomes as siRNA delivery vectors. Curr. Drug Metab. 2014, 15, 882–892. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Safety Study of CALAA-01 to Treat Solid Tumor Cancers. 2013. Available online: https://clinicaltrials.gov/ct2/show/NCT00689065 (accessed on 7 September 2020).
- Mohammadi-Samani, S.; Ghasemiyeh, P. Solid lipid nanoparticles and nanostructured lipid carriers as novel drug delivery systems: Applications, advantages and disadvantages. Res. Pharm. Sci. 2018, 13, 288–303. [Google Scholar] [CrossRef]
- Loh, X.J.; Lee, T.-C.; Dou, Q.; Deen, G.R. Utilising inorganic nanocarriers for gene delivery. Biomater. Sci. 2016, 4, 70–86. [Google Scholar] [CrossRef]
- Kohane, D.S. Microparticles and nanoparticles for drug delivery. Biotechnol. Bioeng. 2006, 96, 203–209. [Google Scholar] [CrossRef]
- Do, T.N.T.; Lee, W.-H.; Loo, C.-Y.; Zavgorodniy, A.V.; Rohanizadeh, R. Hydroxyapatite nanoparticles as vectors for gene delivery. Ther. Deliv. 2012, 3, 623–632. [Google Scholar]
- Loo, S.; Moore, T.; Banik, B.; Alexis, F.; Loo, S.C.J. Biomedical Applications of Hydroxyapatite Nanoparticles. Curr. Pharm. Biotechnol. 2010, 11, 333–342. [Google Scholar] [CrossRef]
- Fatemian, T.; Chowdhury, E.H. Cytotoxicity Enhancement in Breast Cancer Cells with Carbonate Apatite-Facilitated Intracellular Delivery of Anti-Cancer Drugs. Toxics 2018, 6, 12. [Google Scholar] [CrossRef]
- Kamaruzman, N.I.; Tiash, S.; Ashaie, M.; Chowdhury, E.H. siRNAs Targeting Growth Factor Receptor and Anti-Apoptotic Genes Synergistically Kill Breast Cancer Cells through Inhibition of MAPK and PI-3 Kinase Pathways. Biomedicines 2018, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Ashaie, M.; Islam, R.A.; Kamaruzman, N.I.; Ibnat, N.; Tha, K.K.; Chowdhury, E.H. Targeting Cell Adhesion Molecules via Carbonate Apatite-Mediated Delivery of Specific siRNAs to Breast Cancer Cells in Vitro and in Vivo. Pharmaceutics 2019, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Ibnat, N.; Kamaruzman, N.I.; Ashaie, M.; Chowdhury, E.H. Transfection with p21 and p53 tumor suppressor plasmids suppressed breast tumor growth in syngeneic mouse model. Gene 2019, 701, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.; Yamamoto, H.; Chowdhury, E.H.; Wu, X.; Hirose, H.; Haque, A.; Doki, Y.; Mori, M.; Akaike, T. Fabrication and Intracellular Delivery of Doxorubicin/Carbonate Apatite Nanocomposites: Effect on Growth Retardation of Established Colon Tumor. PLoS ONE 2013, 8, e60428. [Google Scholar] [CrossRef] [PubMed]
- Mozar, F.S.; Chowdhury, E.H. Surface-Modification of Carbonate Apatite Nanoparticles Enhances Delivery and Cytotoxicity of Gemcitabine and Anastrozole in Breast Cancer Cells. Pharmaceutics 2017, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.M.; Abidin, S.A.Z.; Chowdhury, E.H. Krebs Cycle Intermediate-Modified Carbonate Apatite Nanoparticles Drastically Reduce Mouse Tumor Burden and Toxicity by Restricting Broad Tissue Distribution of Anticancer Drugs. Cancers 2020, 12, 161. [Google Scholar] [CrossRef]
- Tiash, S.; Chua, M.J.; Chowdhury, E.H. Knockdown of ROS1 gene sensitizes breast tumor growth to doxorubicin in a syngeneic mouse model. Int. J. Oncol. 2016, 48, 2359–2366. [Google Scholar] [CrossRef]
- Chua, M.; Tiash, S.; Fatemian, T.; Noordin, M.I.; Keng, C.; Chowdhury, E. Carbonate apatite-facilitated intracellular delivery of c-ROS1 small interfering RNA sensitises MCF-7 breast cancer cells to cisplatin and paclitaxel. OA Cancer 2013, 1, 1. [Google Scholar] [CrossRef]
- Uskoković, V.; Uskoković, D.P. Nanosized hydroxyapatite and other calcium phosphates: Chemistry of formation and application as drug and gene delivery agents. J. Biomed. Mater. Res. Part B Appl. Biomater. 2010, 96, 152–191. [Google Scholar] [CrossRef]
- Chowdhury, E.H.; Nagaoka, M.; Ogiwara, K.; Zohra, F.T.; Kutsuzawa, K.; Tada, S.; Kitamura, C.; Akaike, T. Integrin-Supported Fast Rate Intracellular Delivery of Plasmid DNA by Extracellular Matrix Protein Embedded Calcium Phosphate Complexes. Biochemistry 2005, 44, 12273–12278. [Google Scholar] [CrossRef]
- Alexis, F.; Pridgen, E.; Molnar, L.K.; Farokhzad, O.C. Factors Affecting the Clearance and Biodistribution of Polymeric Nanoparticles. Mol. Pharm. 2008, 5, 505–515. [Google Scholar] [CrossRef]
- Matsumura, Y.; Maeda, H. A new concept for macromolecular therapeutics in cancer chemotherapy: Mechanism of tumoritropic accumulation of proteins and the antitumor agent smancs. Cancer Res. 1986, 46 Pt 1, 6387–6392. [Google Scholar]
- Truong, N.; Whittaker, M.; Mak, C.W.; Davis, T.P. The importance of nanoparticle shape in cancer drug delivery. Expert Opin. Drug Deliv. 2014, 12, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.; Tada, S.; Akaike, T.; Chowdhury, E.H. Influences of electrolytes and glucose on formulation of carbonate apatite nanocrystals for efficient gene delivery to mammalian cells. Anal. Biochem. 2010, 397, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Heiss, A.; Eckert, T.; Aretz, A.; Richtering, W.; Van Dorp, W.; Schäfer, C.; Jahnen-Dechent, W. Hierarchical Role of Fetuin-A and Acidic Serum Proteins in the Formation and Stabilization of Calcium Phosphate Particles. J. Biol. Chem. 2008, 283, 14815–14825. [Google Scholar] [CrossRef]
- Wopenka, B.; Pasteris, J.D. A mineralogical perspective on the apatite in bone. Mater. Sci. Eng. C 2005, 25, 131–143. [Google Scholar] [CrossRef]
- Bauer, I.W.; Li, S.; Han, Y.-C.; Yuan, L.; Yin, M.-Z. Internalization of hydroxyapatite nanoparticles in liver cancer cells. J. Mater. Sci. Mater. Electron. 2007, 19, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Pagel, M.D. Evaluating pH in the Extracellular Tumor Microenvironment Using CEST MRI and Other Imaging Methods. Adv. Radiol. 2015, 2015, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, E.H. Nanotherapeutics from Laboratory to Clinic; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Corbo, C.; Molinaro, R.; Tabatabaei, M.; Farokhzad, O.C.; Mahmoudi, M. Personalized protein corona on nanoparticles and its clinical implications. Biomater. Sci. 2017, 5, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of nanoparticles with proteins: Relation to bio-reactivity of the nanoparticle. J. Nanobiotechnol. 2013, 11, 26. [Google Scholar] [CrossRef]
- Mu, Q.; Jiang, G.; Chen, L.X.; Zhou, H.; Fourches, D.; Tropsha, A.; Yan, B. Chemical Basis of Interactions Between Engineered Nanoparticles and Biological Systems. Chem. Rev. 2014, 114, 7740–7781. [Google Scholar] [CrossRef] [PubMed]
- Maiorano, G.; Sabella, S.; Sorce, B.; Brunetti, V.; Malvindi, M.A.; Cingolani, R.; Pompa, P.P. Effects of Cell Culture Media on the Dynamic Formation of Protein−Nanoparticle Complexes and Influence on the Cellular Response. ACS Nano 2010, 4, 7481–7491. [Google Scholar] [CrossRef] [PubMed]
- Hunter, G.K.; Hauschka, P.V.; Poole, R.A.; Rosenberg, L.C.; Goldberg, H.A. Nucleation and inhibition of hydroxyapatite formation by mineralized tissue proteins. Biochem. J. 1996, 317 Pt 1, 59–64. [Google Scholar] [CrossRef]
- Cagliani, R.; Gatto, F.; Bardi, G. Protein Adsorption: A Feasible Method for Nanoparticle Functionalization? Materials 2019, 12, 1991. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Malik, A.B. Protein transport across the lung epithelial barrier. Am. J. Physiol. Cell. Mol. Physiol. 2003, 284, L247–L259. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.S.; Khorshid, H.R.K.; Kitchen, J.A.; Arteta, D.; Dalgleish, R. Characterization of the human secreted phosphoprotein 24 gene (SPP2) and comparison of the protein sequence in nine species. Matrix Biol. 2004, 22, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, J.; Chaudhuri, G. Cystatin superfamily. J. Health Care Poor Underserved 2010, 21 (Suppl. 1), 51–70. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Schäfer, C.; Heiss, A.; Gräber, S.; Kinkeldey, A.; Büscher, A.; Schmitt, M.M.; Bornemann, J.; Nimmerjahn, F.; Herrmann, M.; et al. Clearance of Fetuin-A–Containing Calciprotein Particles Is Mediated by Scavenger Receptor-A. Circ. Res. 2012, 111, 575–584. [Google Scholar] [CrossRef]
- Wang, Y.; Okazaki, Y.; Shi, L.; Kohda, H.; Tanaka, M.; Taki, K.; Nishioka, T.; Hirayama, T.; Nagasawa, H.; Yamashita, Y.; et al. Role of hemoglobin and transferrin in multi-wall carbon nanotube-induced mesothelial injury and carcinogenesis. Cancer Sci. 2016, 107, 250–257. [Google Scholar] [CrossRef]
- Shen, Y.; Li, X.; Dong, D.; Zhang, B.; Xue, Y.; Shang, P. Transferrin receptor 1 in cancer: A new sight for cancer therapy. Am. J. Cancer Res. 2018, 8, 916–931. [Google Scholar]
- Mozar, F.S.; Chowdhury, E.H. PEGylation of Carbonate Apatite Nanoparticles Prevents Opsonin Binding and Enhances Tumor Accumulation of Gemcitabine. J. Pharm. Sci. 2018, 107, 2497–2508. [Google Scholar] [CrossRef]
- Law, R.H.P.; Zhang, Q.; McGowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.; Pike, R.N.; Bird, P.I.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shannahan, J.H.; Lai, X.; Ke, P.C.; Podila, R.; Brown, J.M.; Witzmann, F.A. Silver Nanoparticle Protein Corona Composition in Cell Culture Media. PLoS ONE 2013, 8, e74001. [Google Scholar] [CrossRef] [PubMed]
- Konduru, N.V.; Molina, R.M.; Swami, A.; Damiani, F.; Pyrgiotakis, G.; Lin, P.; Andreozzi, P.; Donaghey, T.C.; Demokritou, P.; Krol, S.; et al. Protein corona: Implications for nanoparticle interactions with pulmonary cells. Part. Fibre Toxicol. 2017, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Ziady, A.-G.; Perales, J.C.; Ferkol, T.; Gerken, T.; Beegen, H.; Perlmutter, D.H.; Davis, P.B. Gene transfer into hepatoma cell lines via the serpin enzyme complex receptor. Am. J. Physiol. Content 1997, 273 Pt 1, G545–G552. [Google Scholar] [CrossRef] [PubMed]
- Nadel, J.A. Role of Mast Cell and Neutrophil Proteases in Airway Secretion. Am. Rev. Respir. Dis. 1991, 144 Pt 2, S48–S51. [Google Scholar] [CrossRef]
- Wiogo, H.T.R.; Lim, M.; Bulmus, V.; Yun, J.; Amal, R. Stabilization of Magnetic Iron Oxide Nanoparticles in Biological Media by Fetal Bovine Serum (FBS). Langmuir 2011, 27, 843–850. [Google Scholar] [CrossRef]
- Casals, E.; Pfaller, T.; Duschl, A.; Oostingh, G.J.; Puntes, V. Time Evolution of the Nanoparticle Protein Corona. ACS Nano 2010, 4, 3623–3632. [Google Scholar] [CrossRef]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef]
- Richens, J.L.; Spencer, H.L.; Butler, M.; Cantlay, F.; Vere, K.-A.; Bajaj, N.; Morgan, K.; O’Shea, P. Rationalising the role of Keratin 9 as a biomarker for Alzheimer’s disease. Sci. Rep. 2016, 6, 22962. [Google Scholar] [CrossRef]
- Homberg, M.; Magin, T.M. Beyond expectations: Novel insights into epidermal keratin function and regulation. Int. Rev. Cell Mol. Biol. 2014, 311, 265–306. [Google Scholar]
- Martin, R.I.; Brown, P.W. Formation of hydroxyapatite in serum. J. Mater. Sci. Mater. Electron. 1994, 5, 96–102. [Google Scholar] [CrossRef]
- Chen, B.; Le, W.; Wang, Y.; Li, Z.; Wang, N.; Ren, L.; Lin, L.; Cui, S.; Hu, J.J.; Hu, Y.; et al. Targeting Negative Surface Charges of Cancer Cells by Multifunctional Nanoprobes. Theranostics 2016, 6, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Mass (Da) | pI | Identified Protein | Molecular Function | Biological Process | CA-in-DMEM | CA-in-DMB | HA |
|---|---|---|---|---|---|---|---|---|
| Q546G4 | 68693 | 5.75 | Albumin 1 | Albumin gene family | Carrier protein | ✓ | ✓ | ✓ |
| A0A0G2JGM6 | 14173 | 5.46 | Vitamin D-binding protein | · | ✓ | · | ||
| Q3TIU3 | 37298 | 5.94 | Alpha-2-HS-glycoprotein | Protease inhibitor | carrier protein, bone matrix metabolism, binds Ca and phosphate | ✓ | ✓ | ✓ |
| Q3UEK5 | 37280 | 6.04 | Alpha-2-HS-glycoprotein | ✓ | ✓ | ✓ | ||
| Q3UEK9 | 37326 | 6.04 | Alpha-2-HS-glycoprotein isoform CRA_a | ✓ | ✓ | ✓ | ||
| Q711L0 | 22677 | 8.9 | Secreted phosphoprotein 24 | Protease inhibitor | Bone remodelling | ✓ | ✓ | ∘ |
| D3YTY9 | 53205 | 4.95 | Kininogen-1 | Protease inhibitor | Blood coagulation | · | ✓ | ∘ |
| A0A0R4J0I1 | 46673 | 5.04 | MCG1051009/Serine protease inhibitor A3K | Alpha 1-antichymotrypsin | Acute phase | ∘ | ✓ | ∘ |
| A0A0R4J0X5 | 45854 | 5.32 | Alpha-1-antitrypsin 1-3 | Alpha 1-antitrypsin | ∘ | ✓ | ∘ | |
| A0A0A0MQA3 | 48796 | 5.98 | Alpha-1-antitrypsin 1-1 | ∘ | ✓ | ∘ | ||
| Q3KQQ4 | 45896 | 5.31 | Serpina1a protein | ∘ | ✓ | ∘ | ||
| A0A1B0GS57 | 21329 | 6.58 | Hemopexin | Heme/iron binding | Heme/iron binding and oxygen transport | ✓ | ✓ | · |
| Q8K1U6 | 31047 | 7.64 | Hemopexin | ✓ | ✓ | · | ||
| D3YYR8 | 25676 | 8.34 | Transferrin | · | ✓ | · | ||
| A0A0C6E3V3 | 51022 | 6.82 | HC protein/alpha-1-microglobulin | · | ✓ | · | ||
| Q9QWJ3 | 12907 | 6.78 | Alpha-1-globin | Hemoglobin alpha subunit | ✓ | ✓ | · | |
| Q91VB8 | 15112 | 7.97 | Alpha globin 1 | ✓ | ✓ | · | ||
| A8DUV3 | 15085 | 7.97 | Alpha-globin | ✓ | ✓ | · | ||
| D0U270 | 15840 | 7.13 | Beta-globin | Hemoglobin beta subunit | ✓ | ✓ | · | |
| A8DUP5 | 15894 | 7.09 | Beta-globin | ✓ | ✓ | · | ||
| A8DUK0 | 15840 | 7.13 | Beta-globin | ✓ | ✓ | · | ||
| A8DUP7 | 15826 | 7.13 | Beta-globin | ✓ | ✓ | · | ||
| Q61650 | 5690 | 4.65 | Beta-globin | ✓ | ✓ | · | ||
| A8DUM2 | 15780 | 7.13 | Beta-globin | ✓ | ✓ | · | ||
| D4N6U4 | 15880 | 7.13 | Beta-globin | ✓ | · | · | ||
| B1Q450 | 15840 | 7.13 | Beta-globin | ✓ | · | · | ||
| D0U269 | 15854 | 7.13 | Beta-globin | ✓ | · | · | ||
| Q9CY06 | 15112 | 7.97 | Globin domain-containing protein | Heme, iron, binding | ✓ | ✓ | · | |
| Q9CY10 | 15202 | 8.95 | Globin domain-containing protein | ✓ | ✓ | · | ||
| Q8BPF4 | 15181 | 8.72 | Globin domain-containing protein | ✓ | ✓ | · | ||
| Q6LD55 | 11319 | 6.57 | APOAII | Lipid binding and transport | Opsonin | · | ✓ | · |
| A7YL62 | 11291 | 6.57 | Apolipoprotein A-II | · | ✓ | · | ||
| H7BX99 | 70212 | 6.04 | Prothrombin | Clotting factor | Blood coagulation | · | ✓ | · |
| Q3TJ94 | 70269 | 6.04 | Prothrombin | · | ✓ | · | ||
| A0A075B5P6 | 50063 | 6.6 | Ig mu chain C region | Immunoglobulin | Complement activator, Opsonin | · | ✓ | · |
| A0A075B6A0 | 52618 | 5.99 | Ig mu chain C region | · | ✓ | · | ||
| A0A075B5P4 | 35752 | 7.2 | Ig gamma-1 chain C region | · | ✓ | · | ||
| A0A0A6YWR2 | 43434 | 6.02 | Ig gamma-1 chain C region | · | ✓ | · | ||
| Q99LC4 | 51008 | 6.6 | Igh protein | · | ✓ | · | ||
| I6L985 | 51976 | 8.12 | Igh protein | · | ✓ | · | ||
| E9Q9C6 | 275239 | 4.86 | Fc fragment of IgG-binding protein | Immunoglobulin binding protein | · | · | ✓ | |
| E9Q0B5 | 275224 | 4.87 | Fc fragment of IgG-binding protein | · | · | ✓ | ||
| A2A513 | 57041 | 5 | Keratin type I cytoskeletal 10 | Epithelial intermediate filament | mechanical support | ✓ | ✓ | ✓ |
| Q9CV72 | 30011 | 5.4 | IF rod domain-containing protein | · | · | ✓ | ||
| Q3UV11 | 59526 | 8.32 | Keratin type II cytoskeletal 6B | · | · | ✓ | ||
| Q0VDR7 | 60273 | 8.33 | Krt6b protein | · | · | ✓ | ||
| Q32P04 | 61767 | 7.59 | Keratin 5 | · | · | ✓ | ||
| Q08EK4 | 61302 | 7.73 | Keratin 77 | · | · | ✓ | ||
| B1AQ78 | 44542 | 5.28 | Keratin 19 | · | · | ✓ | ||
| B1AQ77 | 49494 | 4.79 | Keratin 15 isoform CRA_a | · | · | ✓ | ||
| B2RTP7 | 70923 | 8.26 | Krt2 protein | · | · | ✓ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, R.A.; Al-Busaidi, H.; Zaman, R.; Abidin, S.A.Z.; Othman, I.; Chowdhury, E.H. Carbonate Apatite and Hydroxyapatite Formulated with Minimal Ingredients to Deliver SiRNA into Breast Cancer Cells In Vitro and In Vivo. J. Funct. Biomater. 2020, 11, 63. https://doi.org/10.3390/jfb11030063
Islam RA, Al-Busaidi H, Zaman R, Abidin SAZ, Othman I, Chowdhury EH. Carbonate Apatite and Hydroxyapatite Formulated with Minimal Ingredients to Deliver SiRNA into Breast Cancer Cells In Vitro and In Vivo. Journal of Functional Biomaterials. 2020; 11(3):63. https://doi.org/10.3390/jfb11030063
Chicago/Turabian StyleIslam, Rowshan Ara, Hamed Al-Busaidi, Rahela Zaman, Syafiq Asnawi Zainal Abidin, Iekhsan Othman, and Ezharul Hoque Chowdhury. 2020. "Carbonate Apatite and Hydroxyapatite Formulated with Minimal Ingredients to Deliver SiRNA into Breast Cancer Cells In Vitro and In Vivo" Journal of Functional Biomaterials 11, no. 3: 63. https://doi.org/10.3390/jfb11030063
APA StyleIslam, R. A., Al-Busaidi, H., Zaman, R., Abidin, S. A. Z., Othman, I., & Chowdhury, E. H. (2020). Carbonate Apatite and Hydroxyapatite Formulated with Minimal Ingredients to Deliver SiRNA into Breast Cancer Cells In Vitro and In Vivo. Journal of Functional Biomaterials, 11(3), 63. https://doi.org/10.3390/jfb11030063