Anti-Photoaging Effect of Plant Extract Fermented with Lactobacillus buchneri on CCD-986sk Fibroblasts and HaCaT Keratinocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Plant Extract with L. Buchneri (PELB)
2.3. Cell Culture
2.4. MTT Assay for Cell Viability
2.5. UV Irradiation
2.6. Elastase Inhibition Assay
2.7. Type I Procollagen Synthesis Measurement
2.8. Collagenase Inhibition Assay
2.9. Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
2.10. Statistical Analysis
3. Results
3.1. Effects of PELB on Cell Viability
3.2. Effects of PELB on Elastase Activity
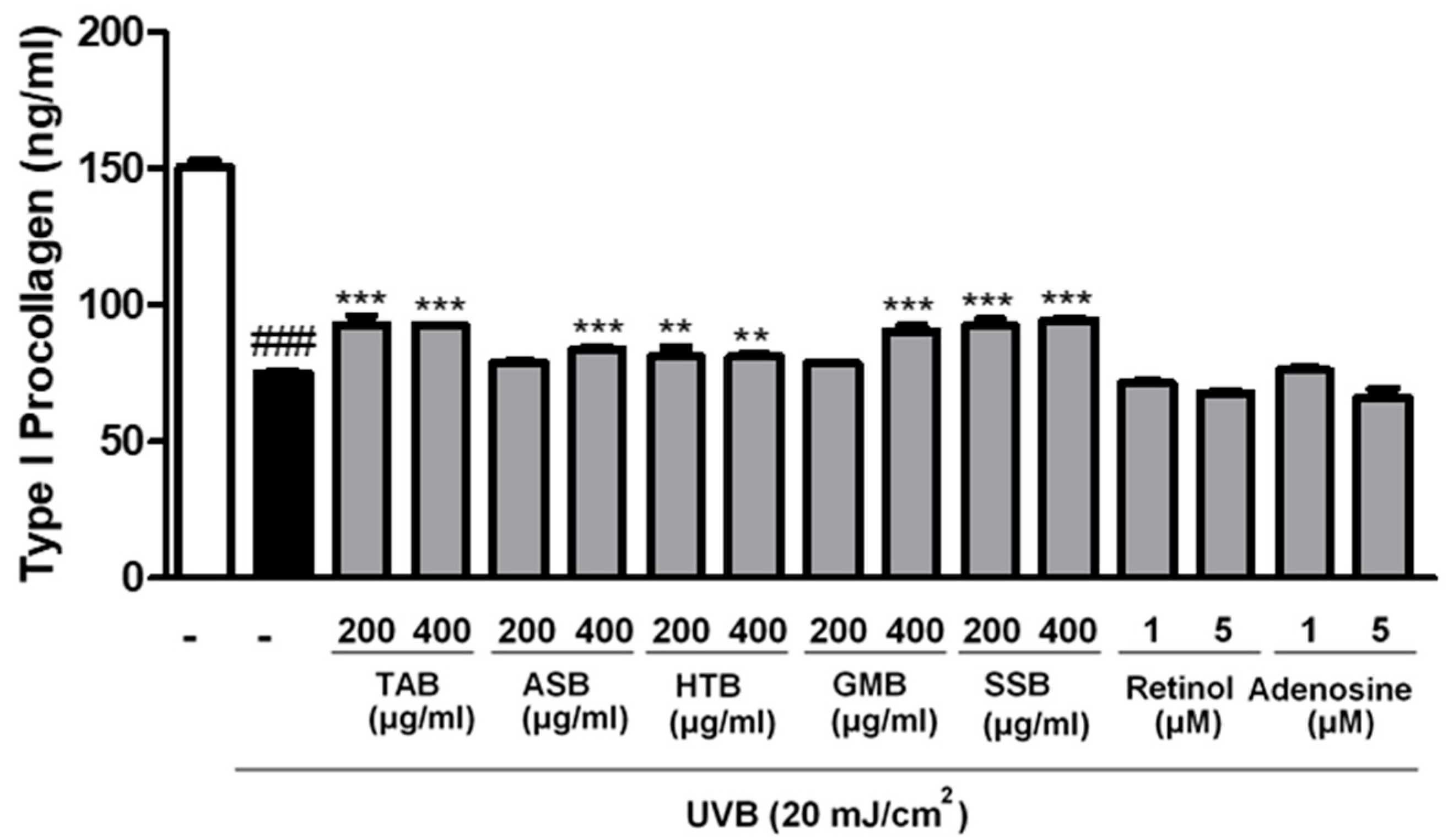
3.3. Effects of PELB on Type I Procollagen Synthesis in UVB-Irradiated CCD-986sk Cells
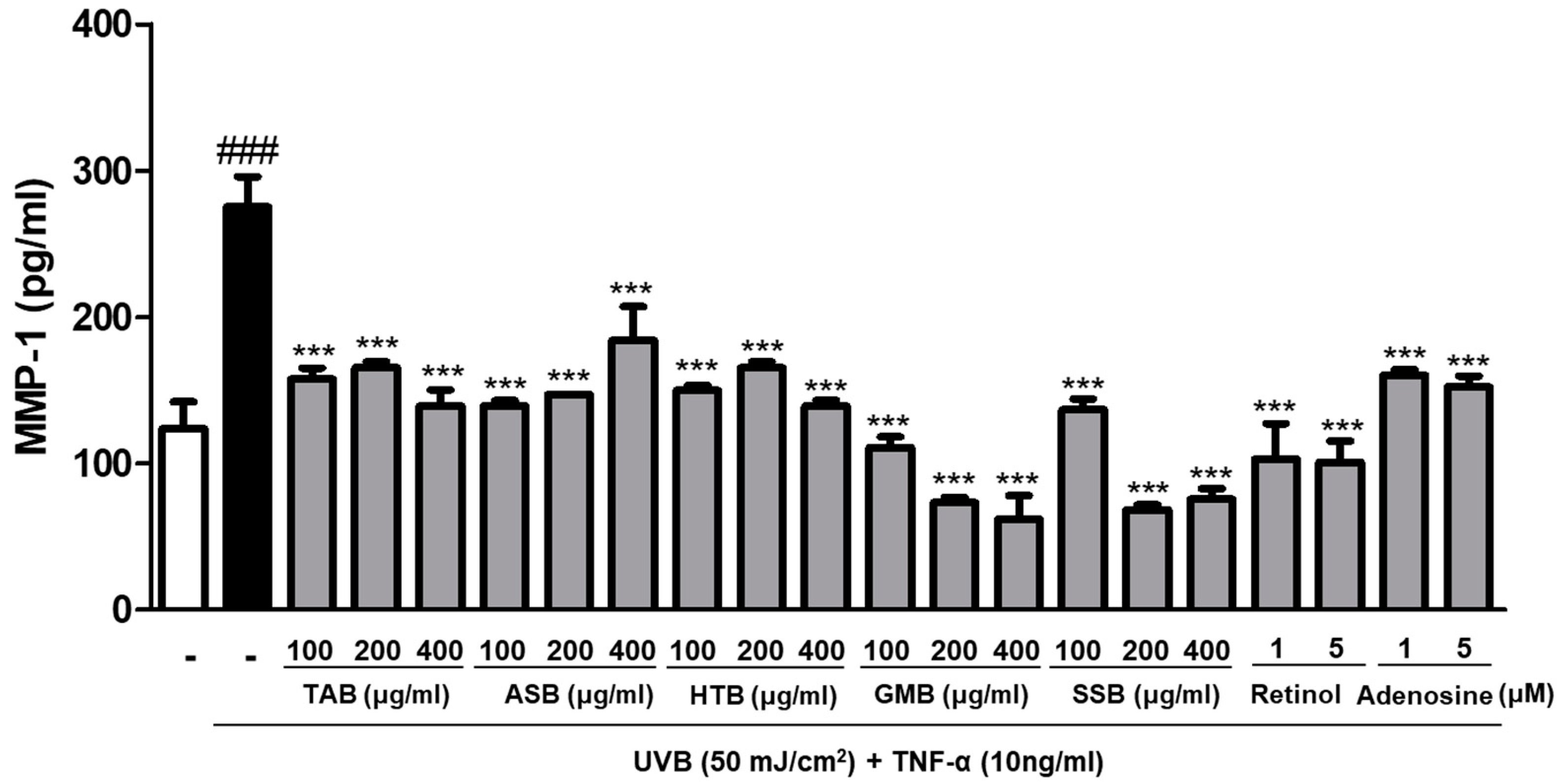
3.4. Effects of PELB on Collagenase Production in UVB-Irradiated HaCaT Keratinocytes
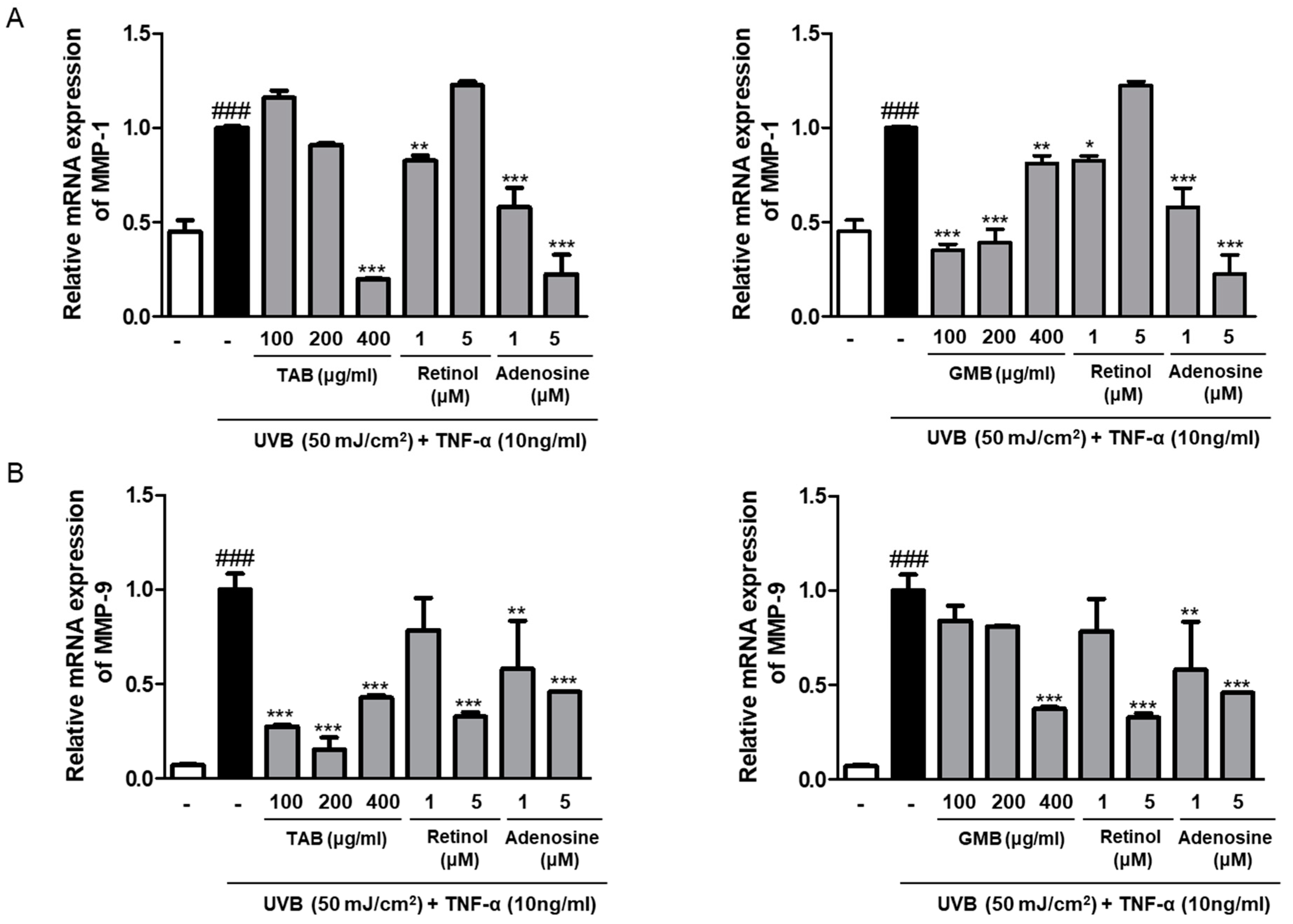
3.5. Effects of PELB on the mRNA Expression of MMPs in UVB-Irradiated HaCaT Keratinocytes
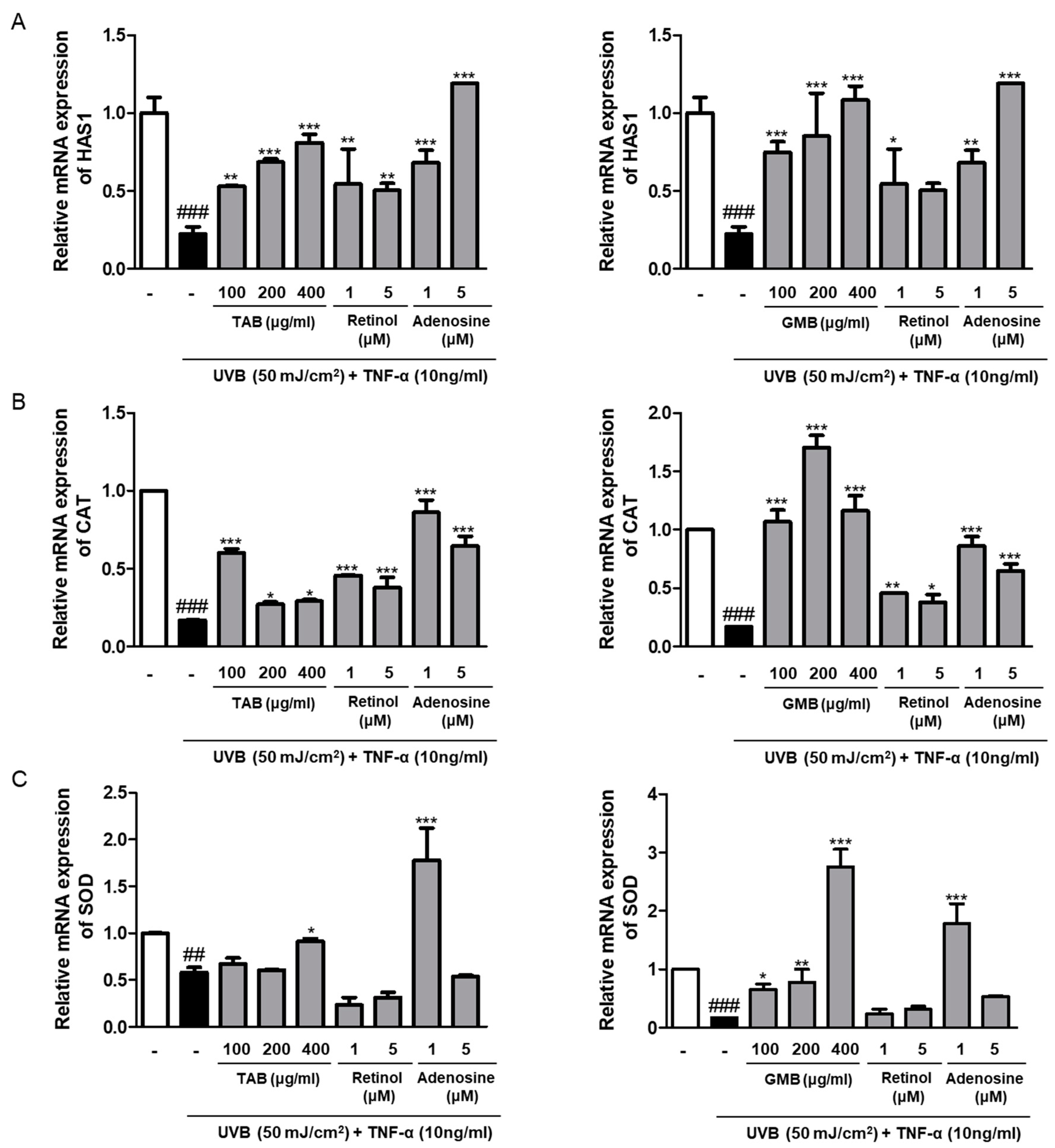
3.6. Effects of PELB on the mRNA Expression of Moisture and Antioxidant Enzymes in UVB-Irradiated HaCaT Keratinocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PELB | Plant extracts fermented with L. Buchneri |
| UV | Ultraviolet |
| ROS | Reactive oxygen species |
| MMPs | Matrix metalloproteinases |
| ECM | Extracellular matrix |
| HAS 1 | Hyaluronic acid synthase 1 |
| CAT | Catalase |
| SOD | Superoxide dismutase |
References
- Kim, M.J.; Woo, S.W.; Kim, M.S.; Park, J.E.; Hwang, J.K. Anti-photoaging effect of aaptamine in UVB-irradiated human dermal fibroblasts and epidermal keratinocytes. J. Asian Nat. Prod. Res. 2014, 16, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Park, S.Y.; Hwang, E.; Zhang, M.; Seo, S.A.; Lin, P.; Yi, T.H. Thymus vulgaris alleviates UVB irradiation induced skin damage via inhibition of MAPK/AP-1 and activation of Nrf2-ARE antioxidant system. J. Cell. Mol. Med. 2017, 21, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Park, E.K.; Lee, H.J.; Lee, H.; Kim, J.H.; Hwang, J.; Koo, J.I.; Kim, S.H. The Anti-Wrinkle Mechanism of Melatonin in UVB Treated HaCaT Keratinocytes and Hairless Mice via Inhibition of ROS and Sonic Hedgehog Mediated Inflammatory Proteins. Int. J. Mol. Sci. 2018, 19, 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.; Hwang, E.; Seo, S.A.; Cho, J.G.; Yang, J.E.; Yi, T.H. Eucalyptus globulus extract protects against UVB-induced photoaging by enhancing collagen synthesis via regulation of TGF-beta/Smad signals and attenuation of AP-1. Arch. Biochem. Biophys. 2018, 637, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Azmi, N.; Hashim, P.; Hashim, D.M.; Halimoon, N.; Majid, N.M. Anti-elastase, anti-tyrosinase and matrix metalloproteinase-1 inhibitory activity of earthworm extracts as potential new anti-aging agent. Asian Pac. J. Trop. Biomed. 2014, 4, S348–S352. [Google Scholar] [CrossRef] [Green Version]
- Xuan, S.H.; Park, Y.M.; Ha, J.H.; Jeong, Y.J.; Park, S.N. The effect of dehydroglyasperin C on UVB-mediated MMPs expression in human HaCaT cells. Pharmacol. Rep. 2017, 69, 1224–1231. [Google Scholar] [CrossRef]
- Pham, Q.L.; Jang, H.J.; Kim, K.B. Antiwrinkle effect of fermented black ginseng on human fibroblasts. Int. J. Mol. Med. 2017, 39, 681–686. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Evaluation of Health and Nutritional Properties of Powder Milk and Live Lactic Acid Bacteria; Food and Agriculture Organization of the United Nations and World Health Organization Expert Consultation Report; FAO: Rome, Italy, 2001. [Google Scholar]
- Sharma, D.; Kober, M.M.; Bowe, W.P. Anti-Aging Effects of Probiotics. J. Drugs Dermatol. 2016, 15, 9–12. [Google Scholar]
- Im, A.R.; Kim, H.S.; Hyun, J.W.; Chae, S. Potential for tyndalized Lactobacillus acidophilus as an effective component in moisturizing skin and anti-wrinkle products. Exp. Ther. Med. 2016, 12, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Singla, V.; Chakkaravarthi, S. Applications of prebiotics in food industry: A review. Food Sci. Technol. Int. 2017, 23, 649–667. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.S. Intestinal flora, probiotics, prebiotics, symbiotics and novel foods. Rev. Esp. Enferm. Dig. 2007, 99, 653–658. [Google Scholar] [PubMed] [Green Version]
- Frei, R.; Akdis, M.; O’Mahony, L. Prebiotics, probiotics, synbiotics, and the immune system: Experimental data and clinical evidence. Curr. Opin. Gastroenterol. 2015, 31, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foolad, N.; Armstrong, A.W. Prebiotics and probiotics: The prevention and reduction in severity of atopic dermatitis in children. Benef. Microbes 2014, 5, 151–160. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Kim, S.S. Probiotics and Prebiotics: Present Status and Future Perspectives on Metabolic Disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Yang, W.; Hostetler, A.; Schultz, N.; Suckow, M.A.; Stewart, K.L.; Kim, D.D.; Kim, H.S. Characterization of the anti-inflammatory Lactobacillus reuteri BM36301 and its probiotic benefits on aged mice. BMC Microbiol. 2016, 16, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, J.; Shan, C.; Liu, S.; Zheng, H.; Liu, C.; Liu, M.; Jin, F.; Wang, L. Skin resistance to UVB-induced oxidative stress and hyperpigmentation by the topical use of Lactobacillus helveticus NS8-fermented milk supernatant. J. Appl. Microbiol. 2017, 123, 511–523. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wu, C.S.; Chao, Y.H.; Lin, C.C.; Tsai, H.Y.; Li, Y.R.; Chen, Y.Z.; Tsai, W.H.; Chen, Y.K. Lactobacillus pentosus GMNL-77 inhibits skin lesions in imiquimod-induced psoriasis-like mice. J. Food Drug Anal. 2017, 25, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of gamma-aminobutyric acid (GABA) by Lactobacillus buchneri isolated from kimchi and its neuroprotective effect on neuronal cells. J. Microbiol. Biotechnol. 2007, 17, 104–109. [Google Scholar] [PubMed]
- Holzer, M.; Mayrhuber, E.; Danner, H.; Braun, R. The role of Lactobacillus buchneri in forage preservation. Trends Biotechnol. 2003, 21, 282–287. [Google Scholar] [CrossRef]
- Zeng, X.Q.; Pan, D.D.; Guo, Y.X. The probiotic properties of Lactobacillus buchneri P2. J. Appl. Microbiol. 2010, 108, 2059–2066. [Google Scholar] [PubMed]
- Zhou, Y.; Ni, X.; Wen, B.; Duan, L.; Sun, H.; Yang, M.; Zou, F.; Lin, Y.; Liu, Q.; Zeng, Y.; et al. Appropriate dose of Lactobacillus buchneri supplement improves intestinal microbiota and prevents diarrhoea in weaning Rex rabbits. Benef. Microbes 2018, 9, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.F.; Lee, H.; Jung, B.J.; Jang, S.; Chung, D.K.; Kim, H. Lipoteichoic acid isolated from Lactobacillus plantarum down-regulates UV-induced MMP-1 expression and up-regulates type I procollagen through the inhibition of reactive oxygen species generation. Mol. Immunol. 2015, 67, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Ko, R.K.; Kim, G.O.; Hyun, C.G.; Jung, D.S.; Lee, N.H. Compounds with tyrosinase inhibition, elastase inhibition and DPPH radical scavenging activities from the branches of Distylium racemosum Sieb. et Zucc. Phytother. Res. 2011, 25, 1451–1456. [Google Scholar] [CrossRef] [PubMed]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care (New Rochelle) 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Brennan, M.; Bhatti, H.; Nerusu, K.C.; Bhagavathula, N.; Kang, S.; Fisher, G.J.; Varani, J.; Voorhees, J.J. Matrix metalloproteinase-1 is the major collagenolytic enzyme responsible for collagen damage in UV-irradiated human skin. Photochem. Photobiol. 2003, 78, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Wong, H.K.; Serrano, J.; Randhawa, M.; Kaur, S.; Southall, M.D.; Parsa, R. Topical stabilized retinol treatment induces the expression of HAS genes and HA production in human skin in vitro and in vivo. Arch. Dermatol. Res. 2017, 309, 275–283. [Google Scholar] [CrossRef]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Cho, J.H.; Hong, S.H.; Kim, D.H.; Jung, H.Y.; Kang, I.K.; Cho, Y.J. Whitening and anti-wrinkle activities of ferulic acid isolated from Tetragonia tetragonioides in B16F10 melanoma and CCD-986sk fibroblast cells. J. Nat. Med. 2018, 72, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Rittie, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Kanimozhi, G.; Prasad, N.R.; Agilan, B.; Ganesan, M.; Mohana, S.; Srithar, G. 7-Hydroxycoumarin prevents UVB-induced activation of NF-kappaB and subsequent overexpression of matrix metalloproteinases and inflammatory markers in human dermal fibroblast cells. J. Photochem. Photobiol. B 2016, 161, 170–176. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Scientific Name | Abbreviation |
|---|---|---|
| 1 | Extract of Triticum aestivum Leaf fermented with Lactobacillus buchneri | TAB |
| 2 | Extract of Avena sativa L. fermented with Lactobacillus buchneri | ASB |
| 3 | Extract of Helianthus tuberosus L. fermented with Lactobacillus buchneri | HTB |
| 4 | Extract of Glycine max Merr. with inner color-greenish fermented with Lactobacillus buchneri | GMB |
| 5 | Extract of Smallanthus sonchifolius fermented with Lactobacillus buchneri | SSB |
| Concentration (μg/mL) | Cell Viability (%) | ||||
|---|---|---|---|---|---|
| TAB | ASB | HTB | GMB | SSB | |
| 0 | 100.00 ± 2.80 | 100.00 ± 2.80 | 100 ± 2.80 | 100 ± 2.80 | 100 ± 2.80 |
| 6.25 | 96.87 ± 3.13 | 98.52 ± 2.21 | 95.92 ± 1.04 | 100.26 ± 2.79 | 97.01 ± 1.29 |
| 12.5 | 96.87 ± 3.55 | 95.83 ± 2.11 | 98.96 ± 2.22 | 99.56 ± 2.39 | 100.49 ± 2.43 |
| 25 | 99.57 ± 2.27 | 97.57 ± 2.81 | 97.13 ± 1.53 | 99.83 ± 3.42 | 97.10 ± 6.01 |
| 50 | 99.39 ± 3.77 | 97.91 ± 0.84 | 96.00 ± 1.70 | 100.09 ± 1.19 | 97.36 ± 3.03 |
| 100 | 100.70 ± 4.70 | 98.87 ± 1.57 | 97.13 ± 0.30 | 101.56 ± 0.75 | 95.88 ± 5.91 |
| 200 | 102.17 ± 3.01 | 99.30 ± 1.45 | 98.52 ± 0.69 | 101.91 ± 1.38 | 98.40 ± 2.74 |
| 400 | 101.91 ± 1.84 | 100.17 ± 1.29 | 100.35 ± 4.05 | 101.48 ± 2.79 | 97.10 ± 5.82 |
| Concentration (μg/mL) | Cell Viability (%) | ||||
|---|---|---|---|---|---|
| TAB | ASB | HTB | GMB | SSB | |
| 0 | 100.00 ± 5.20 | 100.00 ± 5.20 | 100.00 ± 5.20 | 100.00 ± 5.20 | 100.00 ± 5.20 |
| 6.25 | 98.51 ± 6.43 | 102.88 ± 4.37 | 105.22 ± 2.78 | 103.06 ± 5.58 | 103.04 ± 2.49 |
| 12.5 | 100.94 ± 1.98 | 109.41 ± 2.12 | 107.80 ± 4.49 | 105.16 ± 4.88 | 100.97 ± 4.14 |
| 25 | 111.01 ± 5.40 | 108.31 ± 5.44 | 105.55 ± 2.77 | 104.63 ± 0.84 | 94.57 ± 1.65 |
| 50 | 112.53 ± 7.18 * | 110.86 ± 3.26 | 102.79 ± 4.68 | 118.83 ± 0.84 *** | 99.15 ± 2.59 |
| 100 | 130.13 ± 3.97 *** | 112.18 ± 3.80 * | 105.64 ± 4.10 | 114.20 ± 2.89 *** | 103.94 ± 2.20 |
| 200 | 160.24 ± 5.86 *** | 111.35 ± 7.95 * | 115.44 ± 3.44 *** | 132.13 ± 2.51 *** | 102.28 ± 1.92 |
| 400 | 159.50 ± 1.37 *** | 126.46 ± 2.92 *** | 118.46 ± 0.84 *** | 146.48 ± 8.25 *** | 107.54 ± 1.85 * |
| Concentration (μg/mL) | Elastase Inhibition Ratio (%) * | Concentration (μM) | Elastase inhibition Ratio (%) | |||||
|---|---|---|---|---|---|---|---|---|
| TAB | ASB | HTB | GMB | SSB | Retinol | Adenosine | ||
| 6.25 | 16.49 ± 2.29 | −0.63 ± 4.59 | 5.95 ± 6.63 | 3.95 ± 4.41 | 2.62 ± 1.92 | 1.5625 | 6.32 ± 2.54 | 16.55 ± 1.83 |
| 12.5 | 11.39 ± 5.78 | −0.75 ± 6.37 | 9.89 ± 4.54 | 4.36 ± 1.14 | 0.09 ± 1.05 | 3.125 | 13.16 ± 3.09 | 4.37 ± 4.43 |
| 25 | 3.43 ± 3.33 | −2.12 ± 3.28 | 8.82 ± 7.57 | 2.37 ± 7.17 | −1.68 ± 3.35 | 6.25 | 6.90 ± 2.09 | 8.08 ± 1.46 |
| 50 | 7.34 ± 5.65 | 0.80 ± 3.53 | 6.17 ± 5.76 | 3.11 ± 9.31 | −4.18 ± 2.39 | 12.5 | 13.42 ± 1.22 | 2.98 ± 2.09 |
| 100 | 9.94 ± 6.38 | 1.26 ± 3.39 | 10.20 ± 2.32 | 0.22 ± 2.22 | −1.35 ± 3.93 | 25 | 10.09 ± 1.09 | 11.10 ± 3.94 |
| 200 | 1.91 ± 0.37 | −1.36 ± 4.02 | 14.04 ± 9.53 | 6.66 ± 3.34 | −4.54 ± 3.49 | 50 | 14.87 ± 3.80 | 12.87 ± 1.00 |
| 400 | −1.68± 2.37 | 1.21 ± 6.02 | 4.83 ± 2.48 | 5.22 ± 8.88 | −1.87 ± 3.20 | 100 | 1.01 ± 2.24 | 9.94 ± 1.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.-M.; Hong, C.-H.; Kang, S.-H.; Seo, D.-S.; Kim, S.-O.; Lee, H.-Y.; Sim, H.-J.; An, H.-J. Anti-Photoaging Effect of Plant Extract Fermented with Lactobacillus buchneri on CCD-986sk Fibroblasts and HaCaT Keratinocytes. J. Funct. Biomater. 2020, 11, 3. https://doi.org/10.3390/jfb11010003
Kang Y-M, Hong C-H, Kang S-H, Seo D-S, Kim S-O, Lee H-Y, Sim H-J, An H-J. Anti-Photoaging Effect of Plant Extract Fermented with Lactobacillus buchneri on CCD-986sk Fibroblasts and HaCaT Keratinocytes. Journal of Functional Biomaterials. 2020; 11(1):3. https://doi.org/10.3390/jfb11010003
Chicago/Turabian StyleKang, Yun-Mi, Chul-Hee Hong, Sa-Haeng Kang, Dong-Seok Seo, Seong-Oh Kim, Hoon-Yeon Lee, Hyeon-Jae Sim, and Hyo-Jin An. 2020. "Anti-Photoaging Effect of Plant Extract Fermented with Lactobacillus buchneri on CCD-986sk Fibroblasts and HaCaT Keratinocytes" Journal of Functional Biomaterials 11, no. 1: 3. https://doi.org/10.3390/jfb11010003
APA StyleKang, Y.-M., Hong, C.-H., Kang, S.-H., Seo, D.-S., Kim, S.-O., Lee, H.-Y., Sim, H.-J., & An, H.-J. (2020). Anti-Photoaging Effect of Plant Extract Fermented with Lactobacillus buchneri on CCD-986sk Fibroblasts and HaCaT Keratinocytes. Journal of Functional Biomaterials, 11(1), 3. https://doi.org/10.3390/jfb11010003