Abstract
A simple cell population growth model is proposed, where cells are assumed to have a physiological structure (e.g., a model describing cancer cell maturation, where cells are structured by maturation stage, size, or mass). The main question is whether we can guarantee, using the death rate as a control mechanism, that the total number of cells or the total cell biomass has prescribed dynamics, which may be applied to modeling the effect of chemotherapeutic agents on malignant cells. Such types of models are usually described by partial differential equations (PDE). The population dynamics are modeled by an inverse problem for PDE in our paper. The main idea is to reduce this model to a simplified integral equation that can be more easily studied by various analytical and numerical methods. Our results were obtained using the characteristics method.
1. Introduction
The central problem in population ecology is determining how the age structure of a population evolves over time and understanding the factors that regulate animal and plant populations. But age is just one of the many structure variables that demographers and ecologists study. Any quantity that is characteristic of an individual’s state can be used as a structural variable; these include maturation level, size, mass, and so on. Thus, age-structured models are close to more general physiologically structured models. The use of physiologically structured models to describe biological systems has attracted the interest of many researchers and has a long standing tradition. The books by Brauer and Castillo-Chavez [1], Ewens [2], Edelstein-Keshet [3], Kot [4], Rutz [5], and Murray [6] give a good survey of the wide spectrum of applications of such models.
In many real-life problems, all of the input (functional) parameters may not be known a priori and cannot be observed directly. So, in parameter identification problems (a subclass of inverse problems), we ask whether it is possible to take certain additional measurements or, mathematically, to impose some additional conditions describing the modeling process, and thereby determine unknown parameters from these additional experimental data. Inverse problems are currently of great research interest and, hence, the development of efficient analytical and numerical methods to deal with such problems in population biology is an important task of applied mathematics. Direct problems for population models incorporating physiological structures have been extensively studied for many kinds of models. In recent years, many researchers have focused their attention on developing methodologies for solving inverse problems of structured population dynamics (see, e.g., Shcheglov [7], Andrusyak [8], Mobius [9], Bansal et al. [10], Kirk [11], Charleboisa [12], Gyllenberg et al. [13], Perthame and Zubelli [14], Cortes et al. [15], and Borges [16]).
This research is devoted to a simple cell population growth model in which cells are assumed to have a physiological structure (e.g., a model describing cancer cell maturation, where cells are structured by maturation level, size, or mass). For example, Brauer and Castillo-Chavez [1] suggested a simplified model for a chemotherapy course of leukemia; a more detailed motivation is given in Edelstein-Keshet [3] and the role of mathematics in oncology is also discussed in Logan [17] and Murray [6]. We study the simple case when cellular dynamics is modeled by a single first-order PDE analogous to the McKendrick-von Foerster equation. Such an equation is hyperbolic, and the fundamental idea associated with hyperbolic equations is the notion of a characteristic, a curve in space-time along which signals propagate. So, using the effective hyperbolic continuous-time models method of characteristics (see, e.g., Brauer and Castillo-Chavez [1], Logan [17], Mantzaris [18]), a solution to the inverse problem can be expressed as a fixed point of some appropriately chosen integral operator in a suitable metric space. We note that the techniques and methods of this study are close to those used in the author’s previous papers [8,19,20].
The authors [21] study the model of cell division and consider the issue of determining the division rate based on the analysis of the solution of the inverse problem for the differential expansion of the differential–dilation equation. The results of the theoretical analysis are confirmed by several numerical experiments for real data based on the analysis of tumor cells. In paper [22], the inverse problem that describes the model of age-structured cell populations is considered. This model uses the McKendrick-Von Foerster-like equation with a mitosis-dependent death rate. The correctness of the solution is proved based on Fredholm’s integral equation. The size-structured model of cell division and the question of determining the birth rate are considered in [23]. For the numerical analysis, a one-dimensional inverse problem for the integro-differential equation, set on a half-line, was used. The majority of attention is paid to the comparison of the efficiency of different algorithms from the computational point of view. The authors of [24] studied the mathematical properties of a general two-dimensional model of cell division. Since the problem of eigenvalues was solved, the long-time solution convergence was derived. Reference [25] considers the problem of determining the inverse coefficient of recovery of unknown parameters in the biofilm growth model, which is reduced to a problem for a nonlinear partial differential equation with Dirichlet initial conditions and boundary conditions. The conducted numerical studies made it possible to improve the definition of a set of physical quantities related to the biofilm growth model. In [26], the inverse optimization problem with constraints was solved for the partial differential equation, which is the basis of the convection–diffusion model of colorectal cancer cell growth. The conducted test studies illustrate the effectiveness of the proposed method of parameter determination and predict its application to real patient data. The authors of [27] develop and study a tumor growth model and the related inverse problem. The diffusion equation was used to determine the growth rate, depending on the oxygen concentration in the cell. It is supposed that the source of saturation is localized on the blood vessels. The inverse problem of restoring the position of blood vessels under the condition of the distribution of tumor cells was investigated. In [28], the issue of identifying the usually unknown mutation law of cancer cells was analyzed. Local and non-local mathematical models of cellular dynamics and movement were considered. Mutation laws were identified based on macroscopic images of tumors. Since the investigated inverse problems are incorrect, the Tikhonov regularization technique was used. Reference [29] presents a model that describes the dynamics of two adjacent cell populations with different proliferative and mechanical characteristics. The continuous analogue of this discrete model is reduced to a problem with a free boundary for cell densities. An existence result for the free boundary problem is proved and solutions are constructed.
As shown by a review of the problem’s state and an analysis of literary sources, the problem considered in the article is relevant, and finding effective approaches for solving it is an important and challenging task. The purpose of this work is to show the application of PDE theory to a population model. This approach allows us to reduce a quite complicated mathematical problem to much simpler integral equation, incorporating all the required conditions and relationships imposed by the problem.
2. Population Model with Physiological Distribution
Suppose that a population is structured by mass or some other physiological quantity that is characteristic of an individual’s state, say, maturation level, size, and so on. Let be an unknown density of the population at time t with respect to a structural variable x. Here, and in the entire article, we assume, without loss of generality, that x denotes a mass variable. Thus, the number of individuals (cells) at any time t having a mass between and is . Therefore, the total number of individuals (cells) at time t is , where is the mass of all newborns, and M is the maximum possible mass that can be accumulated over lifetime. Assume, also, that individuals (cells) of mass x accumulate mass with a growth rate (i.e., all cells with the same mass experience the same growth rate). In addition, suppose that members may leave the population through death, and that there is a time-dependent per capita death rate . This means that, over the time interval from to , the number of cells with a mass between and die. In the case of modeling the population dynamics of malignant cells, the death rate may vary with chemotherapeutic drug concentration .
But the population may be changed by migration processes, so let the rate of migration be described by a function ; that is, over the time interval from to , the total number of migrants with masses between and is . In population models of cancer cell growth, by migration we mean the process of carcinogenesis, that is, the creation of cancer cells when the genes (DNA) responsible for regulating cell division are damaged, whereby normal cells are transformed into cancer cells. Damage to DNA can be caused by exposure to radiation, chemicals, and other environmental sources, but mutations also accumulate naturally over time through uncorrected errors in DNA transcription. Certain types of cancer can be caused by oncoviruses, and genetics is also significant.
Thus, we obtain the conservation law in integral form
for all .
Assuming the smoothness of and , as well as the continuity of and , Equation (1) may be transformed into the single PDE
for all .
A parent cell at maturity, which is , divides to form two daughter cells of mass . Mathematically, this is translated into a nonlocal boundary condition. Thus, the total number of cell divisions between time and time is . Since this quantity must also be half of , we obtain the renewal boundary condition
3. Formulation of an Inverse Problem; Definition of a Weak Solution
Now we consider the inverse problem of determining how the physiological structure of a cell population evolves over time, finding an unknown functional parameter, which is the time-dependent death rate, so that a weighted integral of the density has prescribed dynamics given by
This integral may represent the total number of cells in the case or the total cell biomass in the case . In other words, in the inverse problem (the parameter identification problem) we seek both the density and the death rate that satisfy the PDE (2) (or, equivalently, the integral Equation (1)) along with conditions (3)–(5).
Note that the physiological variable x and time t are related by the characteristic equation
This equation has the set of solution curves (called characteristics)
where g is assumed to be a positive function, such that is Lebesgue integrable in . Therefore, the time required for individuals to grow from mass to mass is . Characteristics are the fundamental concept in the analysis of hyperbolic problems because PDEs simplify to ODEs along these curves. Denote . Since the function is strictly increasing, and thus it is invertible, the characteristic curve, in coordinate system, passing through the point is easily found to be
where is regarded as a function of with parameters x and t. Here, and subsequently, for notational simplicity, we drop the variables x, t, and write instead of , while keeping in mind that depends on the choice of .
Differentiating the solution along the characteristics yields
Using this relation, Equation (2) can be transformed to
Then, by the variation of parameters method, which works for all linear equations, we can find a particular solution to (7)
Thus, the general solution to (7) has the following form:
where is to be determined.
Make the change of variables , to obtain
and, therefore,
Taking , , that is, considering the previous equation on the characteristics that emanate from points on the x axis, where , gives, for all , ,
Similarly, taking , , that is, considering Equation (8) on the characteristics that emanate from points , gives, for all , ,
Definition 1.
4. Reduction of the Inverse Problem to an Integral Equation
Now we find, from the last two equations, an explicit formula for the unknown in terms of . To this end, we first consider the set , in which, by substituting the initial condition (4) into Equation (11), we have
where .
Further, in the set , applying the renewal boundary condition (3) to Equation (12), we obtain
from which, by Equation (13),
where, for simplicity of notation, we write instead of dropping the variables x, t but keeping in mind that depends on the choice of x and t.
Similarly, in the set , we have
where we write instead of , dropping the variables x and t.
Continuing in this way, in each set , where n is an integer greater than one, we deduce, after a finite number of steps, that
where for all nonnegative integers k.
For convenience, we also introduce the notation . Note that can be interpreted as the probability that a malignant cell will not be killed by chemotherapeutic agents up to time t, given that it was alive at time . Substituting Equations (13) and (14) into the additional condition (5) and changing the order of integration gives, in the interval ,
In a similar way, substituting Equations (14) and (15) into condition (5), we obtain, in the interval ,
Continuing in this way, in each interval , where n is a positive integer, we finally deduce that
Theorem 1.
Finding a solution to the inverse problem can be reduced to solving, for the unknown E, the integral Equations (17) and (18) considered in the corresponding intervals. More precisely,
- a weak solution to the inverse problem gives the solution of these integral equations, and this solution is locally absolutely continuous in ;
Note that, from the property of E being locally absolutely continuous in , it follows that this function is differentiable, i.e., in this interval, and its derivative is locally Lebesgue integrable (see [19]).
5. An Example of Integral Equation Modelling the Dynamics of an Unknown Coefficient
Let us fix the parameters and known functions of the problem: , , , , where , , , . Then, consider the following problem:
Using the notation from Equation (19), along with conditions (20) and (21), we deduce the equality
in the set , and
in the set .
Substituting the equalities above into condition (22), we have an integral equation for the unknown E in the interval :
The last equation can be rewritten in the following form:
which is a linear Volterra equation of the second kind. This equation can be solved numerically by the method of successive approximations using programming. To make visualizations of approximations, fix the constants: , , . Applying four iterations, we see, in Figure 1, the corresponding exponent-like approximations to the solution of the integral equation in the interval .
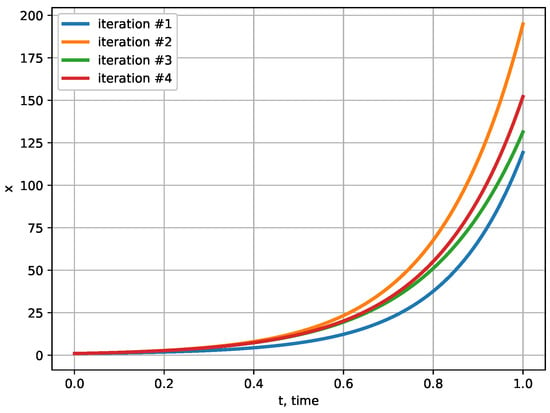
Figure 1.
Exponent-like approximations to solution of integral equation.
To investigate the exponential behaviour of these curves, let us logarithmize the obtained approximations. Plotting the logarithm of approximations, we see they tend to a linear function with a slope of 5, that is the function (see Figure 2). Moreover, the graphs of all approximations starting from the fourth are linear in the considered interval.
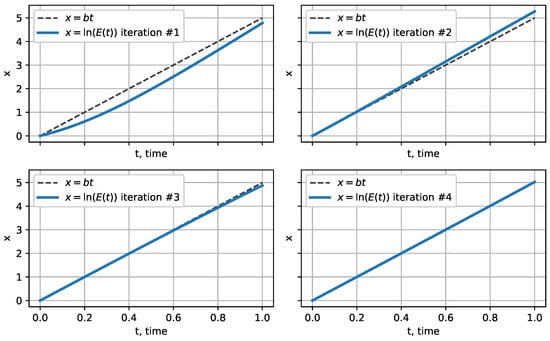
Figure 2.
Approximations tend to a linear function with a slope of 5.
Therefore, we conclude that successive approximations of solution E approach the exponential function , which can be clearly seen in Figure 3.
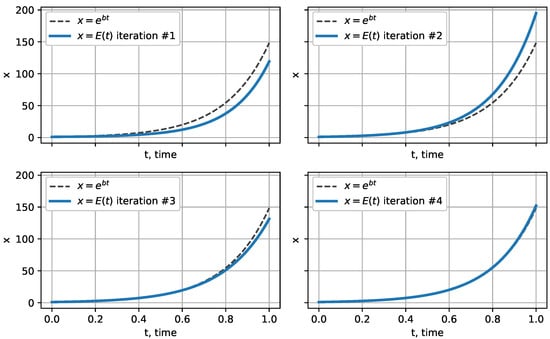
Figure 3.
Successive approximations approach the exponential function .
6. Discussion
In recent decades, integral boundary conditions have been actively used as override conditions for solving inverse initial boundary value problems for parabolic partial differential equations [30]. The first-order hyperbolic systems with smoothing boundary conditions of the integral type (5) appear, in particular, in applications in population dynamics [31,32]. The approaches and results of this paper are a development of the ideas and methods mentioned above and have a similar work plan for describing a mathematical model of population growth. The imposition of redefined conditions (5) of the integral type is also motivated by the results obtained in [33]. The strong regularity properties obtained in this paper for solutions of hyperbolic problems with integral boundary conditions show that such problems are well suited for the implementation of numerical schemes (since these properties ensure the stability of the schemes). The computational complexity of the method of successive approximations for the numerical solution of the linear Volterra equation of the second kind is defined as the number of iterations as per the computational complexity of the numerical quadratures used in the work (the method of numerical integration). The computational complexity can be estimated approximately by the formula , where k is the number of iterations, N is the number of points in the grid to calculate the integral, and M is the number of calculations to evaluate the integral (depending on the chosen numerical method). It should also be borne in mind that the accuracy of the approximation can affect the computational complexity, and some iterative methods may require a lot of computation to achieve the desired accuracy. Thus, for each iteration of the method of successive approximations using the Gaussian quadrature formula with N nodes, the number of calculations is approximately . That is, In the proposed work, , so . A small number of iterations will reduce the computational complexity. This is an advantage of the proposed approach compared to others. Note, in particular, that the use of the collocation method under the conditions of piecewise constant approximation of the desired function for the numerical solution of an integral equation has a computational complexity of [34].
Thus, in the case of using another method for solving an integral equation, the computational complexity is , while based on the proposed algorithm it is . The use of the method of characteristics made it possible to reduce the solution of the original problem to a simplified form. Thus, the computational complexity is reduced and a more efficient algorithm is obtained. We also note that the use of traditional numerical methods for solving the original (direct) problem also leads to higher computational complexity [35]. Thus, the use of the inverse problem approach is also motivated by computational aspects.
We additionally provide the link to the GitHub repository https://github.com/ivanna-andrusyak/volterra-integral-equation/blob/main/volterra_eq.ipynb with (accessed on 20 January 2024) the underlying computer code replicating the calculations in our paper.
Experimental data for real biological problems show, in particular, the significant heterogeneity observed in cancer populations under physiological conditions. Therefore, this fact undoubtedly introduces limitations for the mathematical modeling of the growth process. The mathematical model considered in this paper is no exception. In this regard, one of the possible improvement strategies is the procedure of homogenising the environment. The homogenisation (averaging) of the population may make it possible to consider the model problem studied in this article as a certain approximation of a structurally homogeneous environment to the actual heterogeneous structure of a cell population. The possibility of such averaging (homogenisation) has been proposed in many works investigating processes in structurally heterogeneous environments (see, for example, [36]). It is clear that only experimental studies should evaluate the effectiveness of the choice of homogenisation procedure for different types of non-heterogeneous populations. In addition, they would make it possible to analyze the practical adequacy of the approach, the theoretical advantages of which are in the significant simplification of the computational method. This task is interesting in itself for independent scientific research and will be the subject of further study.
7. Conclusions
Population dynamics have traditionally been and still are a dominant branch of mathematical biology. Population models play a critical role in helping us understand the dynamic processes involved, make actionable predictions, and thus better understand the natural world. The results obtained in this paper show that the cell population can be modeled using an inverse problem for an unknown density function and a coefficient representing the death rate. We have developed a technique for solving such an inverse problem. It consists of reducing the search for a solution to the problem to solving an integral equation with an unknown function that depends only on death rate. It is clear that solving such an integral equation is a simpler problem, and various numerical approaches can be used to obtain an approximation of the solution. Based on the methodology developed in this paper and the obtained results, it is possible to consider the conditions required for the existence of a unique classical solution, as well as a solution in the weak sense for the inverse problem, in the future, which is important for the justification of numerical approaches.
The results of this study can be used as a starting point for further research into complex population models arising in various areas of real life—medicine, ecology, epidemiology, etc. Also, they can be used as a mathematical background for developing programming solutions for population growth modelling, which could contribute to making population dynamics more manageable and predictable.
Studying the existence of positive cell death rate , the model solvability in the case of monotonic , representing the decreasing total number of cells or the total cell biomass or, more generally, the conditions under which the problem is well-posed in the weak sense are all interesting areas for further research, as these properties are important for controlling cell dynamics and could have applications in real-world problems. The considered population model was certainly hard to solve numerically, but the results of our work simplify it significantly and can be easily programmed. This study should be rather considered as a basis for further investigation in that direction.
Author Contributions
Conceptualization, I.A. and P.P.; methodology I.A., M.V. and O.B.; software, O.B.; validation, M.V. and O.B.; investigation, P.P. and M.V.; writing—original draft preparation, M.V. and I.A.; writing—review and editing, I.A. and P.P.; visualization, M.V. and O.B.; project administration, I.A. and P.P.; funding acquisition, P.P. and M.V. All authors have read and agreed to the published version of the manuscript.
Funding
The results were obtained as part of the work under a grant from the Ministry of Education and Science of Ukraine (project number 0123U101691).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Brauer, F.; Castillo-Chavez, C. Mathematical Models in Population Biology and Epidemiology; Springer: New York, NY, USA; Dordrecht, The Netherlands; Heidelberg, Germany; London, UK, 2012. [Google Scholar] [CrossRef]
- Ewens, W.J. Mathematical Population Genetics; Springer: New York, NY, USA, 2004. [Google Scholar] [CrossRef]
- Edelstein-Keshet, L. Mathematical Models in Biology; SIAM: Philadelphia, PA, USA, 2008. [Google Scholar]
- Kot, M. Elements of Mathematical Ecology; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar] [CrossRef]
- Borges Rutz, R. Mathematical Models of Physiologically Structured Cell Populations; Universitat Autònoma de Barcelona: Barcolona, Spain, 2012; Available online: http://surl.li/nmxxc (accessed on 20 November 2023).
- Murray, J.D. Mathematical Biology I. An Introduction, 3rd ed.; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Shcheglov, A.Y. Inverse Problem for a Model of the Dynamics of a Population with Symmetric Cell Division. Mosc. Univ. Comput. Math. Cybern. 2022, 46, 50–59. [Google Scholar] [CrossRef]
- Andrusyak, R.V. Inverse problem for a physiologically structured population model with variable-effort harvesting. Open Math. 2017, 15, 433–445. [Google Scholar] [CrossRef]
- Mobius, W.; Laan, L. Physical and Mathematical Modeling in Experimental Papers. Cell 2015, 163, 1577–1583. [Google Scholar] [CrossRef][Green Version]
- Bansal, S.; Read, J.; Pourbohloul, B.; Meyers, L. The dynamic nature of contactnetworks in infectious disease epidemiology. J. Biol. Dyn. 2010, 4, 478–489. [Google Scholar] [CrossRef]
- Kirk, P.; Thorne, T.; Stumpf, M.P. Model selection in systems and synthetic biology. Curr. Opin. Biotechnol. 2013, 24, 767–774. [Google Scholar] [CrossRef]
- Charleboisa, D.A.; Gabor, B. Modeling cell population dynamics. Silico Biol. 2019, 13, 21–39. [Google Scholar] [CrossRef]
- Gyllenberg, M.; Osipov, A.; Päivärinta, L. The inverse problem of linear age-structured population dynamics. J. Evol. Equ. 2002, 2, 223–239. [Google Scholar] [CrossRef]
- Perthame, B.; Zubelli, J.P. On the inverse problem for a size-structured population model. Inverse Probl. 2007, 23, 1037–1052. [Google Scholar] [CrossRef]
- Cortes, M.G.; Trinh, J.T.; Zeng, L.; Balázsi, G. Late-arriving signals contribute less to cell fate decisions. Biophys. J. 2017, 113, 2110–2120. [Google Scholar] [CrossRef]
- Borges, R.; Calsina, A.; Cuadrado, S. Oscillations in a molecular structured cell population model. Nonlinear Anal. Real World Appl. 2011, 12, 1911–1922. [Google Scholar] [CrossRef][Green Version]
- Logan, J.D. An Introduction to Nonlinear Partial Differential Equations; Wiley Interscience: New York, NY, USA, 2008; Available online: http://surl.li/nmxyx (accessed on 20 November 2023).
- Mantzaris, N.V.; Daoutidis, P. Cell population balance modeling and control in continuous bioreactors. J. Process Control 2004, 14, 775–784. [Google Scholar] [CrossRef]
- Leoni, G. A First Course in Sobolev Spaces; AMS: Providence, RI, USA, 2009; Available online: https://cvgmt.sns.it/paper/937/ (accessed on 29 November 2023).
- Andrusyak, I.V.; Filevych, P.V. The minimal growth of entire functions with given zeros along unbounded sets. Mat. Stud. 2020, 54, 146–153. [Google Scholar] [CrossRef]
- Groh, A.; Krebs, J.; Wagner, M. Efficient solution of an inverse problem in cell population dynamics. Inverse Probl. 2011, 27, 065009. [Google Scholar] [CrossRef]
- Clément, F.; Laroche, B.; Robin, F. Analysis and numerical simulation of an inverse problem for a structured cell population dynamics model. Math. Biosci. Eng. 2019, 16, 3018–3046. [Google Scholar] [CrossRef]
- Zubelli, J.P.; Albani, V. Structured Models for Cell Populations: Direct and Inverse Problems. ITM Web Conf. 2015, 5, 00016. [Google Scholar] [CrossRef]
- Doumic, M. Analysis of a Population Model Structured by the Cells Molecular Content. Math. Model. Nat. Phenom. 2007, 2, 121–152. [Google Scholar] [CrossRef][Green Version]
- Brander, T.; Lesnic, D.; Cao, K. Inverse problems for a model of biofilm growth. IMA J. Appl. Math. 2023, 88, 258–281. [Google Scholar] [CrossRef]
- Figueiredo, I.N.; Leal, C. Physiologic parameter estimation using inverse problems. SIAM J. Appl. Math. 2013, 73, 1164–1182. [Google Scholar] [CrossRef]
- Colin, T.; Iollo, A.; Lagaert, J.-B.; Saut, O. An inverse problem for the recovery of the vascularization of a tumor. J. Inverse ILL-Posed Probl. 2014, 22, 759–786. [Google Scholar] [CrossRef]
- Alwuthaynani, M.; Eftimie, R.; Trucu, D. Inverse problem approaches for mutation laws in heterogeneous tumours with local and nonlocal dynamics. Math. Biosci. Eng. 2022, 19, 3720–3747. [Google Scholar] [CrossRef]
- Lorenzi, T.; Murray, P.J.; Ptashnyk, M. From individual-based mechanical models of multicellular systems to free-boundary problems. Interfaces Free Bound. 2020, 22, 205–244. [Google Scholar] [CrossRef]
- Lesnic, D.; Yousefi, S.A.; Ivanchov, M. Determination of a time-dependent diffusivity from nonlocal conditions. J. Appl. Math. Comput. 2013, 41, 301–320. [Google Scholar] [CrossRef]
- Eftimie, R. Hyperbolic and kinetic models for self-organized biological aggregations and movement: A brief review. J. Math. Biol. 2012, 65, 35–75. [Google Scholar] [CrossRef]
- Magal, P.; Ruan, S. Center Manifolds for Semilinear Equations with Non-Dense Domain and Applications to Hopf Bifurcation in Age Structured Models; American Mathematical Society; Providence, RI, USA, 2009; Volume 202, pp. 1–80. [Google Scholar] [CrossRef]
- Kmit, I. Smoothing effect and Fredholm property for first-order hyperbolic PDEs. Oper. Theory Adv. Appl. 2013, 231, 219–238. [Google Scholar] [CrossRef]
- Mochurad, L.; Shakhovska, K.; Montenegro, S. Parallel Solving of Fredholm Integral Equations of the First Kind by Tikhonov Regularization Method Using OpenMP Technology. Adv. Intell. Syst. Comput. 2020, 1080, 25–35. [Google Scholar] [CrossRef]
- Lapidus, L.; Pinder, G.F. Numerical Solution of Partial Differential Equations in Science and Engineering; John Wiley and Sons: New York, NY, USA, 1999. [Google Scholar] [CrossRef]
- Khoroshun, L.P. Mathematical Models and Methods of the Mechanics of Stochastic Composites. Int. Appl. Mech. 2000, 36, 1284–1316. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).