
Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
2.3. Statistical Analysis
3. Results
3.1. Overall
3.2. Temporal Pattern
3.2.1. Monthly Detection Rate
3.2.2. Diel Cycle
3.3. Spatial Pattern
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bérubé, M.; Aguilar, A.; Dendanto, D.; Larsen, F.; Sciara, G.N.; Sears, R.; Sigurjonsson, J.; Palsbøll, P.J. Population genetic structure of North Atlantic, Mediterranean Sea and Sea of Cortez fin whales, Balaenoptera physalus (Linnaeus 1758): Analysis of mitochondrial and nuclear loci. Mol. Ecol. 1998, 7, 585–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannier, A. Summer distribution of fin whales (Balaenoptera physalus) in the northwestern Mediterranean marine mammals sanctuary. Rev. d’Ecol. 2002, 57, 135–150. [Google Scholar]
- Laran, S.; Gannier, A. Spatial and temporal prediction of fin whale distribution in the northwestern Mediterranean Sea. ICES J. Mar. Sci. 2008, 65, 1260–1269. [Google Scholar] [CrossRef]
- Di Sciara, G.N.; Zanardelli, M.; Jahoda, M.; Panigada, S.; Airoldi, S. The fin whale Balaenoptera physalus (L. 1758) in the Mediterranean Sea. Mammal Rev. 2003, 33, 105–150. [Google Scholar] [CrossRef]
- Panigada, S.; Zanardelli, M.; MacKenzie, M.; Donovan, C.; Mélin, F.; Hammond, P.S. Modelling habitat preferences for fin whales and striped dolphins in the Pelagos Sanctuary (Western Mediterranean Sea) with physiographic and remote sensing variables. Remote Sens. Environ. 2008, 112, 3400–3412. [Google Scholar] [CrossRef]
- Panigada, S.; Pesante, G.; Zanardelli, M.; Capoulade, F.; Gannier, A.; Weinrich, M.T. Mediterranean fin whales at risk from fatal ship strikes. Mar. Pollut. Bull. 2006, 52, 1287–1298. [Google Scholar] [CrossRef]
- Forcada, J.; di Sciara, G.N.; Fabbri, F. Abundance of fin whales and striped dolphins summering in the Corso-Ligurian Basin. Mammalia 1995, 59, 127–140. [Google Scholar] [CrossRef]
- Druon, J.N.; Panigada, S.; David, L.; Gannier, A.; Mayol, P.; Arcangeli, A.; Cañadas, A.; Laran, S.; Méglio, N.; Di Gauffier, P. Potential feeding habitat of fin whales in the western Mediterranean Sea: An environmental niche model. Mar. Ecol. Prog. Ser. 2012, 464, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Cominelli, S.; Moulins, A.; Rosso, M.; Tepsich, P. Fin whale seasonal trends in the Pelagos Sanctuary, Mediterranean Sea. J. Wildl. Manag. 2016, 80, 490–499. [Google Scholar] [CrossRef] [Green Version]
- Di Sciara, G.N. La cetofauna del bacino corso-ligure-provenzale: Rassegna delle attuali conoscenze. Biol. Mar. Mediterr. 1994, 1, 95–98. [Google Scholar]
- Arcangeli, A.; Orasi, A.; Carcassi, S.P.; Crosti, R. Exploring thermal and trophic preference of Balaenoptera physalus in the central Tyrrhenian Sea: A new summer feeding ground? Mar. Biol. 2014, 161, 427–436. [Google Scholar] [CrossRef]
- Goffart, A.; Prieur, L.; Hecq, J.H. Contrôle du phytoplancton du bassin Ligure par le front liguro-provençal (secteur Corse). Biometrics 1995, 51, 888–898. [Google Scholar]
- Esposito, A.; Manzella, G. Current circulation in the ligurian sea. Elsevier Oceanogr. Ser. 1982, 34, 187–203. [Google Scholar]
- Astraldi, M.; Gasparini, G.P.; Sparnocchia, S. The seasonal and interannual variability in the Ligurian-Provençal Basi. J. Geophys. Res. 2011, 46, 93–113. [Google Scholar]
- Astraldi, M.; Bianchi, C.N.; Gasparini, G.P.; Morri, C. Climatic fluctuations, current variability and marine species distribution: A case study in the Ligurian Sea (north-west Mediterranean). Oceanol. Acta 1995, 18, 139–149. [Google Scholar]
- Jacques, G. L’oligotrophie du milieu pélagique de Méditerranée Occidentale: Un paradigme qui s’estompe? Bull. Soc. Zool. Fr. 1989, 114, 17–30. [Google Scholar]
- Arnau, P.; Liquete, C.; Canals, M. River mouth plume events and their dispersal in the northwestern Mediterranean Sea. Oceanography 2004, 17, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Gonella, J.; Lamy, A.; Kartavtseff, A. Rapport II-5. Hydrodynamique côtière en régime d’été. J. L’hydraul. 1977, 14, 1–4. [Google Scholar]
- Aïssi, M.; Celona, A.; Comparetto, G.; Mangano, R.; Würtz, M.; Moulins, A. Large-scale seasonal distribution of fin whales (Balaenoptera physalus) in the central Mediterranean Sea. J. Mar. Biol. Assoc. UK 2008, 88, 1253–1261. [Google Scholar] [CrossRef]
- Castellote, M.; Clark, C.W.; Lammers, M.O. Population Identity and Migration Movements of Fin Whales (Balaenoptera physalus) in the Mediterranean Sea and Strait of Gibraltar; IWC Scientific Committee: Agadir, Morocco, 2010. [Google Scholar]
- Panigada, S.; Donovan, G.P.; Druon, J.N.; Lauriano, G.; Pierantonio, N.; Pirotta, E.; Zanardelli, M.; Zerbini, A.N.; Di Sciara, G.N. Satellite tagging of Mediterranean fin whales: Working towards the identification of critical habitats and the focussing of mitigation measures. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Canese, S.; Cardinali, A.; Fortuna, C.M.; Giusti, M.; Lauriano, G.; Salvati, E.; Greco, S. The first identified winter feeding ground of fin whales (Balaenoptera physalus) in the Mediterranean Sea. J. Mar. Biol. Assoc. UK 2006, 86, 903–907. [Google Scholar] [CrossRef]
- Sciacca, V.; Caruso, F.; Beranzoli, L.; Chierici, F.; De Domenico, E.; Embriaco, D.; Favali, P.; Giovanetti, G.; Larosa, G.; Marinaro, G.; et al. Annual acoustic presence of fin whale (Balaenoptera physalus) offshore Eastern Sicily, central Mediterranean Sea. PLoS ONE 2015, 10, e01411838. [Google Scholar] [CrossRef] [Green Version]
- Geijer, C.K.A.; di Sciara, G.N.; Panigada, S. Mysticete migration revisited: Are Mediterranean fin whales an anomaly? Mammal Rev. 2016, 46, 284–296. [Google Scholar] [CrossRef]
- Marini, L.; Consiglio, C.; Angradi, A.M.; Catalano, B.; Sanna, A.; Valentini, T.; Finoia, M.G.; Villetti, G. Distribution, abundance and seasonality of cetaceans sighted during scheduled ferry crossings in the central tyrrhenian sea: 1989–1992. Ital. J. Zool. 1996, 63, 381–388. [Google Scholar] [CrossRef]
- Laran, S.; Drouot-Dulau, V. Seasonal variation of striped dolphins, fin- and sperm whales’ abundance in the Ligurian Sea (Mediterranean Sea). J. Mar. Biol. Assoc. UK 2007, 87, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Simone, P.; Lauriano, G.; Burt, L.; Pierantonio, N.; Donovan, G. Monitoring winter and summer abundance of cetaceans in the Pelagos Sanctuary (Northwestern Mediterranean sea) through aerial surveys. PLoS ONE 2011, 6, e22878. [Google Scholar]
- Martin, S.W.; Marques, T.A.; Thomas, L.; Morrissey, R.P.; Jarvis, S.; Dimarzio, N.; Moretti, D.; Mellinger, D.K. Estimating minke whale (Balaenoptera acutorostrata) boing sound density using passive acoustic sensors. Mar. Mammal Sci. 2013, 29, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Watkins, W.A.; Daher, M.A.; Reppucci, G.M.; George, J.E.; Martin, D.L.; DiMarzio, N.A.; Gannon, D.P. Seasonahty and distribution of whale calls in the North Pacific. Oceanography 2000, 13, 62–67. [Google Scholar] [CrossRef]
- Castellote, M.; Clark, C.W.; Lammers, M.O. Fin whale (Balaenoptera physalus) population identity in the western Mediterranean Sea. Mar. Mammal Sci. 2012, 28, 325–344. [Google Scholar] [CrossRef] [Green Version]
- Širović, A.; Hildebrand, J.A.; Wiggins, S.M. Blue and fin whale call source levels and propagation range in the Southern Ocean. J. Acoust. Soc. Am. 2007, 122, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.W.; Fristrup, K.M. Whales ’95: A Combined Visual and Acoustic Survey of Blue and Fin Whales off Southern California. Rep. Int. Whal. Comm. 1997, 47, 583–600. [Google Scholar]
- Cerchio, S.; Jacobsen, J.K.; Norris, T.F. Temporal and geographical variation in songs of humpback whales, Megaptera novaeangliae: Synchronous change in Hawaiian and Mexican breeding assemblages. Anim. Behav. 2001, 62, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Charif, R.A.; Mellinger, D.K.; Dunsmore, K.J.; Fristrup, K.M.; Clark, C.W. Estimated source levels of fin whale (Balaenoptera physalus) vocalizations: Adjustments for surface interference. Mar. Mammal Sci. 2002, 18, 81–98. [Google Scholar] [CrossRef]
- Clark, C.W.; Gagnon, G.J. Low-frequency vocal behaviors of baleen whales in the north atlantic: Insights from integrated undersea surveillance system detections, locations, and tracking from 1992 to 1996. J. Underw. Acoust. 2002, 52, 48. [Google Scholar]
- Gedamke, J.; Robinson, S.M. Acoustic survey for marine mammal occurrence and distribution off East Antarctica (30–80° E) in January–February 2006. Deep Res. Part II Top. Stud. Oceanogr. 2010, 57, 968–981. [Google Scholar] [CrossRef]
- Thompson, P.O.; Findley, L.T.; Vidal, O. 20-Hz pulses and other vocalizations of fin whales, Balaenoptera physalus, in the Gulf of California, Mexico. J. Acoust. Soc. Am. 1992, 92, 3051–3057. [Google Scholar] [CrossRef]
- Pereira, A.; Harris, D.; Tyack, P.; Matias, L. Fin whale acoustic presence and song characteristics in seas to the southwest of Portugal. J. Acoust. Soc. Am. 2020, 147, 2235–2249. [Google Scholar] [CrossRef] [PubMed]
- Richardson, W.J.; Greene, C.R.; Malme, C.I.; Thomson, D.H.; Moore, S.E.; Wiirsig, B. Marine Mammals and Noise, 1st ed.; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Charif, R.A.; Clark, C.W. Acoustic monitoring of large whales in deep waters north and west of the British Isles: 1996–2005. Cornell Lab Ornithol. 2009, 08, 40. [Google Scholar]
- Watkins, W.A.; Tyack, P.; Moore, K.E.; &Bird, J.E. The 20-Hz signals of finback whales (Balaenoptera physalus). J. Acoust. Soc. Am. 1987, 82, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Watkins, W.A. Activities and underwater sounds of fin whales (Balaenoptera physalus). Sci. Rep. Whales Res. Inst. 1981, 33, 83–117. [Google Scholar]
- McDonald, M.A.; Fox, C.G. Passive acoustic methods applied to fin whale population density estimation. J. Acoust. Soc. Am. 1999, 105, 2643–2651. [Google Scholar] [CrossRef]
- Tyack, P.L.; Clark, C.W. Communication and acoustic behavior of dolphins and whales. In Hearing by Whales and Dolphins; Au, W.W.L., Popper, A.N., Fay, R.R., Eds.; Springer: New York, NY, USA, 2000; pp. 156–224. [Google Scholar]
- Oleson, E.M.; Širović, A.; Bayless, A.R.; Hildebr, J.A. Synchronous seasonal change in fin whale song in the North Pacific. PLoS ONE 2014, 9, e115678. [Google Scholar] [CrossRef] [Green Version]
- Hatch, L.T.; Clark, C.W. Acoustic Differentiation between Fin Whales in Both the North Atlantic and North Pacific Oceans, and Integration with Genetic Estimates of Divergence; IWC Scientific Committee: Agadir, Morocco, 2004; 37p. [Google Scholar]
- Delarue, J.; Todd, S.K.; Van Parijs, S.M.; Di Iorio, L. Geographic variation in Northwest Atlantic fin whale (Balaenoptera physalus) song: Implications for stock structure assessment. J. Acoust. Soc. Am. 2009, 125, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Širović, A.; Oleson, E.M.; Buccowich, J.; Rice, A.; Bayless, A.R. Fin whale song variability in southern California and the Gulf of California. Sci. Rep. 2017, 7, 1–11. [Google Scholar]
- Simon, M.; Stafford, K.M.; Beedholm, K.; Lee, C.M.; Madsen, P.T. Singing behavior of fin whales in the Davis Strait with implications for mating, migration and foraging. J. Acoust. Soc. Am. 2010, 128, 3200–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Opzeeland, I.; Van Parijs, S.; Kindermann, L.; Burkhardt, E.; Boebel, O. Calling in the cold: Pervasive acoustic presence of humpback whales (Megaptera novaeangliae) in antarctic coastal waters. PLoS ONE 2013, 8, e73007. [Google Scholar]
- Clark, C.W.; Borsani, J.F.; Di Sciara, G.N. Vocal activity of fin whales, Balaenoptera physalus, in the Ligurian Sea. Mar. Mammal Sci. 2002, 18, 286–295. [Google Scholar] [CrossRef]
- Borsani, J.F.; Clark, C.W.; Nani, B.; Scarpiniti, M. Fin whales avoid loud rhythmic low- frequency sounds in the ligurian sea. Bioacoustics 2008, 17, 161–163. [Google Scholar] [CrossRef]
- Lammers, M.O.; Brainard, R.E.; Au, W.W.L.; Mooney, T.A.; Wong, K.B. An ecological acoustic recorder (EAR) for long-term monitoring of biological and anthropogenic sounds on coral reefs and other marine habitats. J. Acoust. Soc. Am. 2008, 123, 1720–1728. [Google Scholar] [CrossRef] [Green Version]
- Giorli, G.; Au, W.W.L.; Ou, H.; Jarvis, S.; Morrissey, R.; Moretti, D. Acoustic detection of biosonar activity of deep diving odontocetes at Josephine Seamount High Seas Marine Protected Area. J. Acoust. Soc. Am. 2015, 137, 2495–2501. [Google Scholar] [CrossRef]
- Giorli, G.; Au, W.W.L.; Neuheimer, A. Differences in foraging activity of deep sea diving odontocetes in the Ligurian Sea as determined by passive acoustic recorders. Deep Res. Part I Oceanogr. Res. Pap. 2016, 107, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sciacca, V.; Viola, S.; Pulvirenti, S.; Riccobene, G.; Caruso, F.; De Domenico, E.; Pavan, G. Shipping noise and seismic airgun surveys in the Ionian Sea: Potential impact on Mediterranean fin whale. Proc. Meet. Acoust. 2016, 27, 1. [Google Scholar]
- EMODnet Bathymetry Consortium: EMODnet Digital Bathymetry (DTM 2018). Eur. Mar. Obs. Data Netw. 2018. [CrossRef]
- QGIS.org. QGIS Geographic Information System. Open SourceGeospatial Foundation Project. 2020. Available online: http://qgis.org (accessed on 24 August 2021).
- Simone, P.; Zanardelli, M.; Canese, S.; Jahoda, M. How deep can baleen whales dive? Mar. Ecol. Prog. Ser. 1999, 187, 309–311. [Google Scholar]
- Goffart, A.; Hecq, J.H.; Legendre, L. Drivers of the winter-spring phytoplankton bloom in a pristine NW Mediterranean site, the Bay of Calvi (Corsica): A long-term study (1979–2011). Prog. Oceanogr. 2015, 137, 121–139. [Google Scholar] [CrossRef]
- Goffart, A.; Hecq, J.H.; Legendre, L. Changes in the development of the winter-spring phytoplankton bloom in the Bay of Calvi (NW Mediterranean) over the last two decades: A response to changing climate? Mar. Ecol. Prog. Ser. 2002, 236, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Macias, D.; Garcia-Gorriz, E.; Stips, A. Deep winter convection and phytoplankton dynamics in the NW Mediterranean Sea under present climate and future (horizon 2030) scenarios. Sci. Rep. 2018, 8, 6626. [Google Scholar] [CrossRef] [PubMed]
- Tepsich, P.; Schettino, I.; Atzori, F.; Azzolin, M.; Campana, I.; Carosso, L.; Cominelli, S.; Crosti, R.; David, L.; Di-Méglio, N.; et al. Trends in summer presence of fin whales in the western mediterranean sea Region: New insights from a long-term monitoring program. PeerJ 2020, 8, e10544. [Google Scholar] [CrossRef]
- Croll, D.A.; Clark, C.W.; Acevedo, A.; Tershy, B.; Flores, S.; Gedamke, J.; Urban, J. Only male fin whales sing loud songs. Nature 2002, 417, 809. [Google Scholar] [CrossRef] [PubMed]
- Weirathmueller, M.J.; Stafford, M.; Wilcock, W.S.D.; Hilmo, R.S.; Dziak, R.P.; Tréhu, A.M. Spatial and temporal trends in fin whale vocalizations recorded in the NE Pacific Ocean between 2003–2013. PLoS ONE 2017, 12, e0186127. [Google Scholar] [CrossRef] [Green Version]
- Onsrud, M.S.R.; Kaartvedt, S. Diel vertical migration of the krill MEganyctiphanes norvegica in relation to physical environment, food and predators. Mar. Ecol. Prog. Ser. 1998, 171, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Kaartvedt, S. Diel Vertical Migration Behaviour of the Northern Krill (Meganyctiphanes norvegica Sars). Adv. Mar. Biol. 2010, 57, 255–275. [Google Scholar] [PubMed]
- Endo, Y.; Yamano, F. Diel vertical migration of Euphausia pacifica (crustacea, euphausiacea) in relation to molt and reproductive processes, and feeding activity. J. Oceanogr. 2006, 62, 693–703. [Google Scholar] [CrossRef]
- Keen, E.M.; Falcone, E.A.; Andrews, R.D.; Schorr, G.S. Diel dive behavior of fin whales (Balaenoptera physalus) in the Southern California Bight. Aquat. Mammal 2019, 45, 233–243. [Google Scholar] [CrossRef]
- Wiggins, S.M.; Hildebrand, J.A. Fin whale 40-Hz calling behavior studied with an acoustic tracking array. Mar. Mammal Sci. 2020, 36, 964–971. [Google Scholar] [CrossRef]
- Azzellino, A.; Gaspari, S.; Airoldi, S.; Nani, B. Habitat use and preferences of cetaceans along the continental slope and the adjacent pelagic waters in the western Ligurian Sea. Deep Res. Part I Oceanogr. Res. Pap. 2008, 55, 296–323. [Google Scholar] [CrossRef]
- Cotté, C.; Guinet, C.; Taupier-Letage, I.; Mate, B.; Petiau, E. Scale-dependent habitat use by a large free-ranging predator, the Mediterranean fin whale. Deep Res. Part I Oceanogr. Res. Pap. 2009, 56, 801–811. [Google Scholar] [CrossRef]
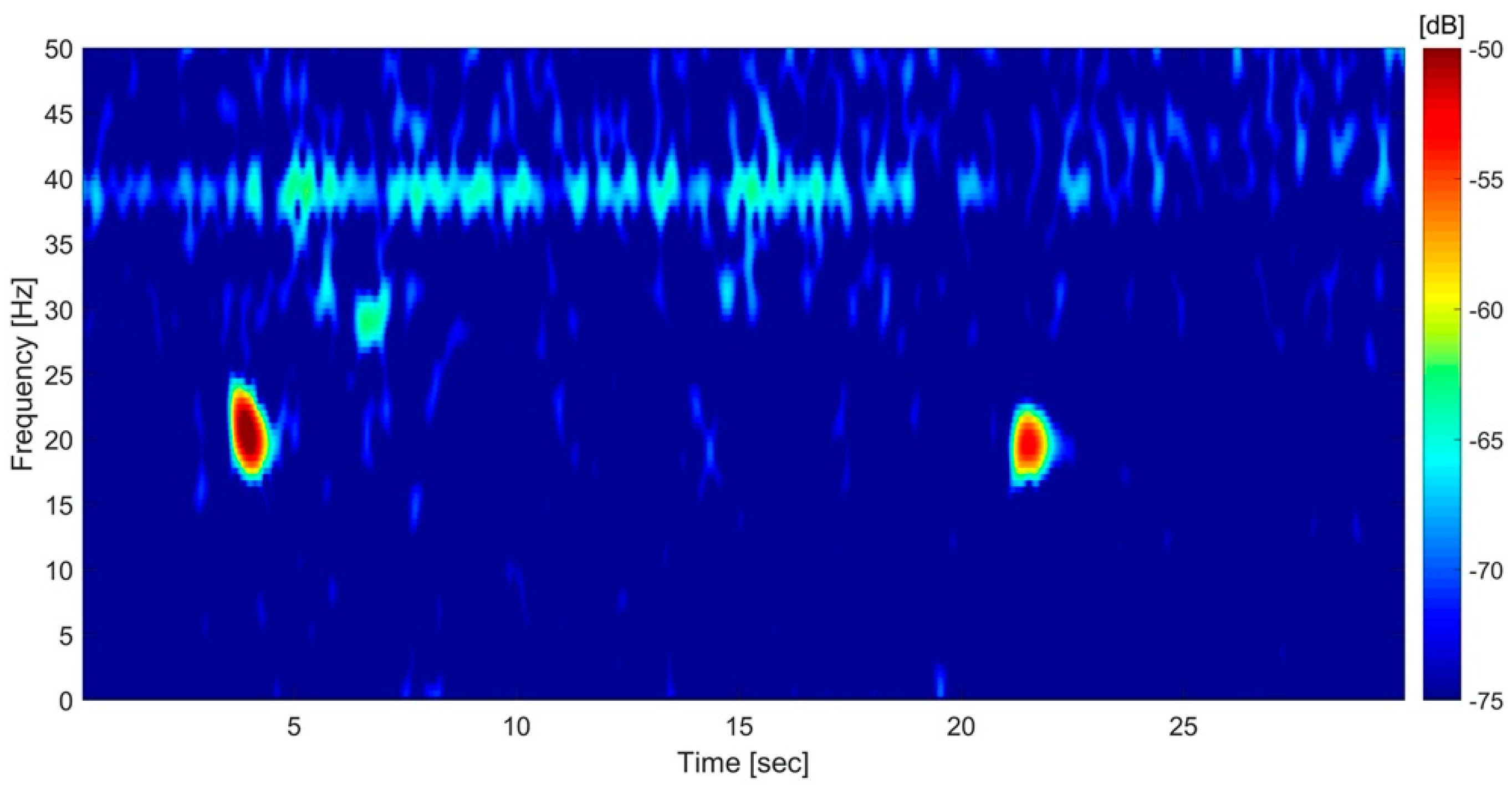







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Rate (Hz) | Duty Cycle (s) | Depth (m) | Latitude (Decimal Deg) | Longitude (Decimal Deg) | Recording Periods | |
|---|---|---|---|---|---|---|
| EAR1 | 80,000 | 120 | 900 | 44.0858 | 8.4601 | 07/22 to 12/8 2011 |
| EAR2 | 80,000 | 120 | 870 | 44.1513 | 8.5795 | 07/22 to 11/3 2011 |
| EAR3 | 80,000 | 240 | 700 | 44.1964 | 8.6157 | 07/22 to 12/9 2011 |
| EAR4 | 80,000 | 240 | 890 | 44.178 | 8.6857 | 07/22 to 09/22 2011 |
| EAR5 | 80,000 | 240 | 880 | 44.0639 | 8.9244 | 07/22 to 11/13 2011 |
| NFD | Latitude (Decimal Deg) | Longitude (Decimal Deg) | Coast Distance (m) | EAR Depth (m) | Seabed Depth (m) | Slope Mean (Deg) | Slope StDv (Deg) | |
|---|---|---|---|---|---|---|---|---|
| EAR1 | 1428 | 44.0858 | 8.4601 | 11,340 | 900 | 1058 | 5.68 | 4.47 |
| EAR2 | 416 | 44.1513 | 8.5795 | 13,320 | 870 | 1095 | 5.46 | 3.83 |
| EAR3 | 1769 | 44.1964 | 8.6157 | 14,400 | 700 | 815 | 5.30 | 3.79 |
| EAR4 | 355 | 44.178 | 8.6587 | 18,360 | 890 | 1099 | 4.28 | 2.96 |
| EAR5 | 727 | 44.0639 | 8.9244 | 33,390 | 880 | 985 | 6.49 | 4.65 |
| EAR1 | EAR2 | EAR3 | EAR4 | EAR5 | TOT | |
|---|---|---|---|---|---|---|
| Number of Files | 59,588 | 23,877 | 61,014 | 22,255 | 42,995 | 209,729 |
| Number of Files with Detections | 1428 | 416 | 1769 | 355 | 727 | 4695 |
| Hourly Detection Rate (Detections/Hours) | 0.0673 | 0.0519 | 0.0591 | 0.1233 | 0.0388 | 0.341 |
| Number of Recording Hours | 2817 | 1277 | 3116 | 1483 | 2534 | 11,227 |
| Kruskall–Wallis | Tamhane Test | ||||
|---|---|---|---|---|---|
| Instrument | N | X2 | p Value | Comparison | p Value |
| EAR1 | 2817 | 43.191 | <0.001 | July–November | 0.017 |
| July–December | 0.009 | ||||
| August–November | 0.018 | ||||
| August–December | 0.013 | ||||
| September–November | 0.045 | ||||
| September–December | 0.020 | ||||
| EAR2 | 1277 | 9.048 | 0.060 | / | / |
| EAR3 | 3116 | 117.797 | <0.001 | July/August/September–October/November/December | <0.001 |
| EAR4 | 1483 | 12.265 | 0.007 | August–September | 0.032 |
| EAR5 | 2534 | 41.505 | <0.001 | July/August/Speptember–October | 0.001 |
| July–November | 0.030 | ||||
| August–November | 0.038 | ||||
| September–November | 0.035 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintore, L.; Sciacca, V.; Viola, S.; Giacoma, C.; Papale, E.; Giorli, G. Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn. J. Mar. Sci. Eng. 2021, 9, 966. https://doi.org/10.3390/jmse9090966
Pintore L, Sciacca V, Viola S, Giacoma C, Papale E, Giorli G. Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn. Journal of Marine Science and Engineering. 2021; 9(9):966. https://doi.org/10.3390/jmse9090966
Chicago/Turabian StylePintore, Laura, Virginia Sciacca, Salvatore Viola, Cristina Giacoma, Elena Papale, and Giacomo Giorli. 2021. "Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn" Journal of Marine Science and Engineering 9, no. 9: 966. https://doi.org/10.3390/jmse9090966
APA StylePintore, L., Sciacca, V., Viola, S., Giacoma, C., Papale, E., & Giorli, G. (2021). Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn. Journal of Marine Science and Engineering, 9(9), 966. https://doi.org/10.3390/jmse9090966