
Decadal Assessment of Sperm Whale Site-Specific Abundance Trends in the Northern Gulf of Mexico Using Passive Acoustic Data

 and
and
Abstract
1. Introduction
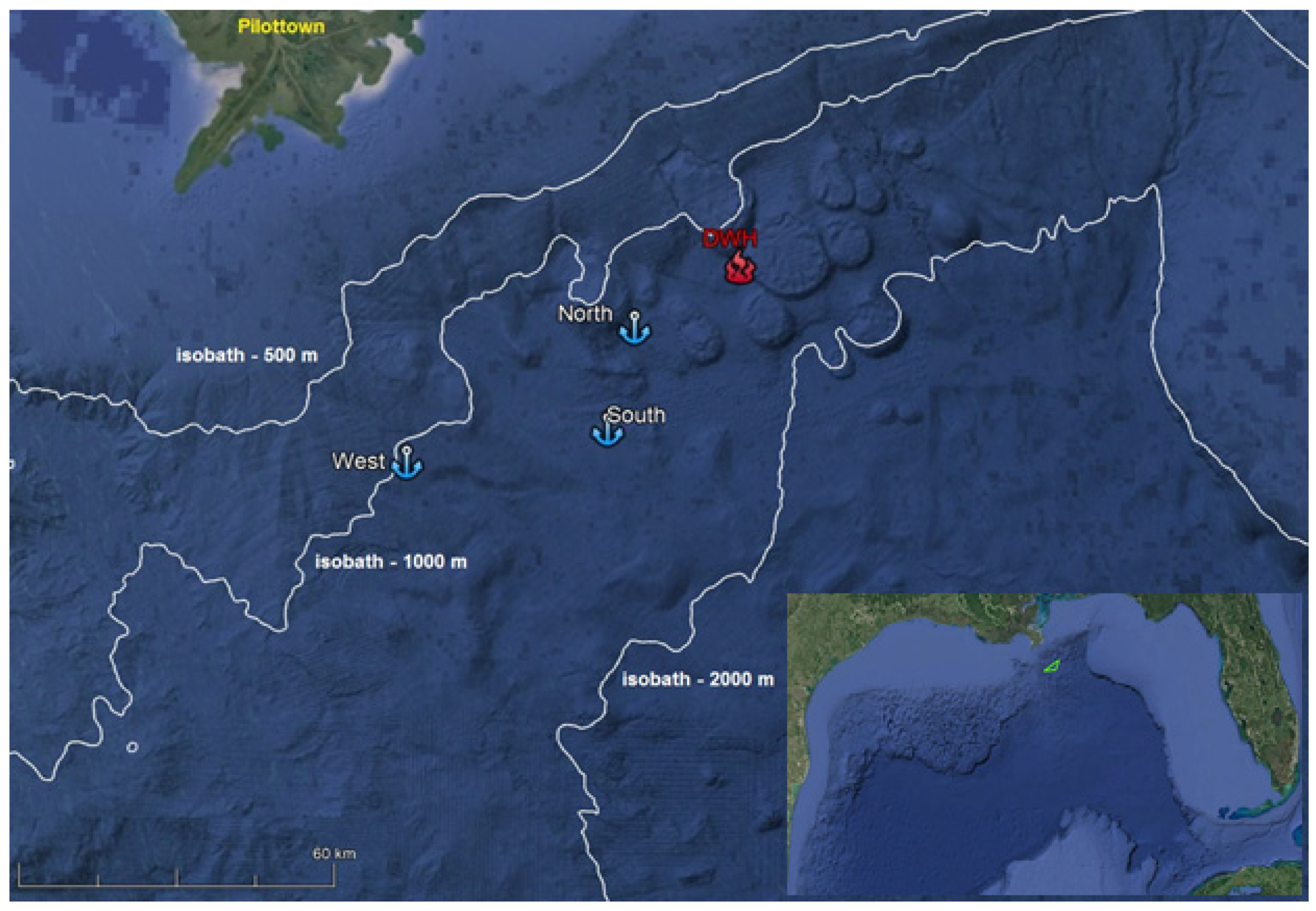
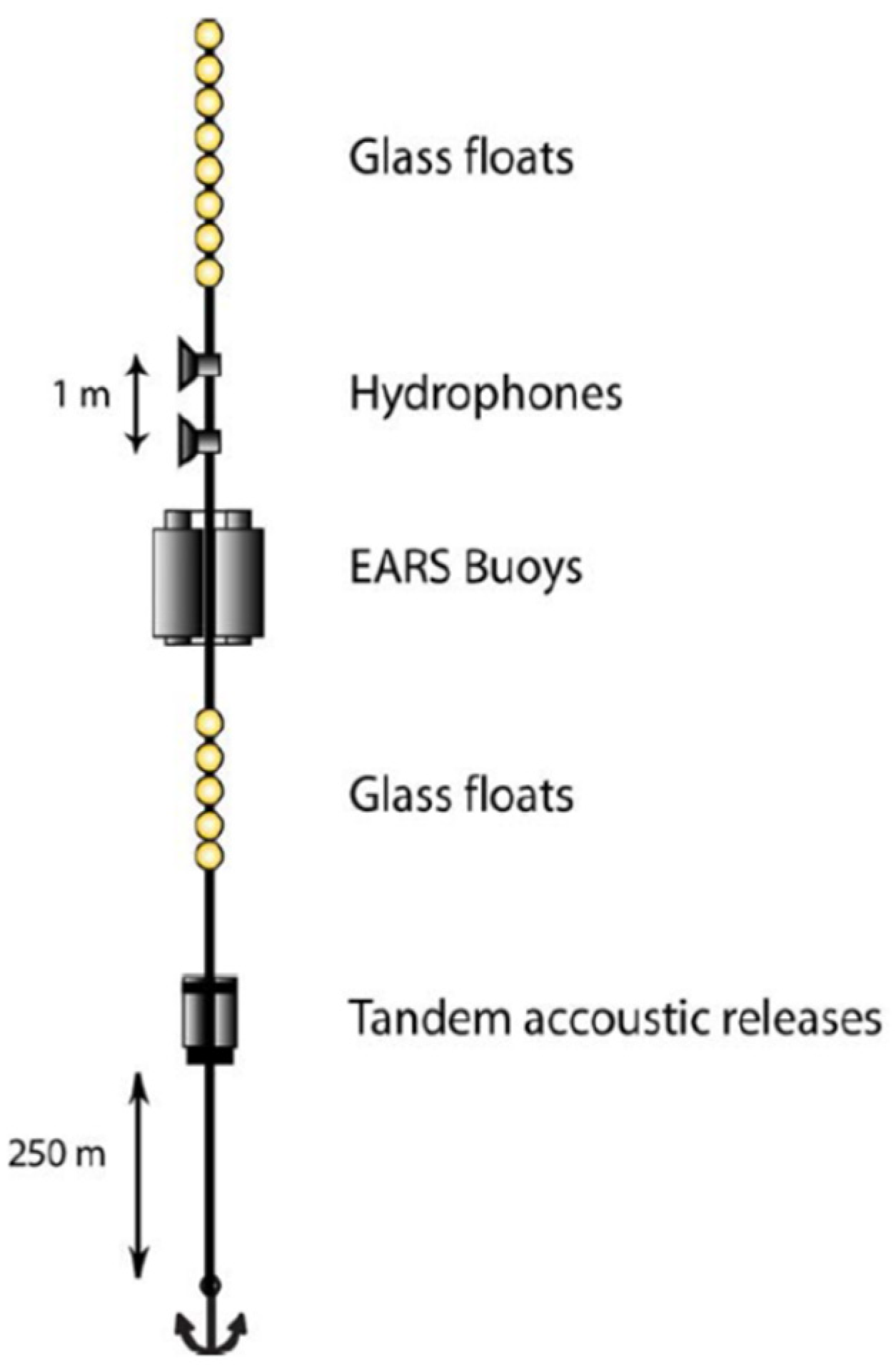
2. Experiment and Data Collection
3. Methods
3.1. Population Density Estimation
3.2. Click Detection
3.3. False Positive Rate
3.4. Probability of Detection
3.5. The Maximum Detection Range
3.6. Production Rate
3.7. Time Period
3.8. Variance of Density Estimates
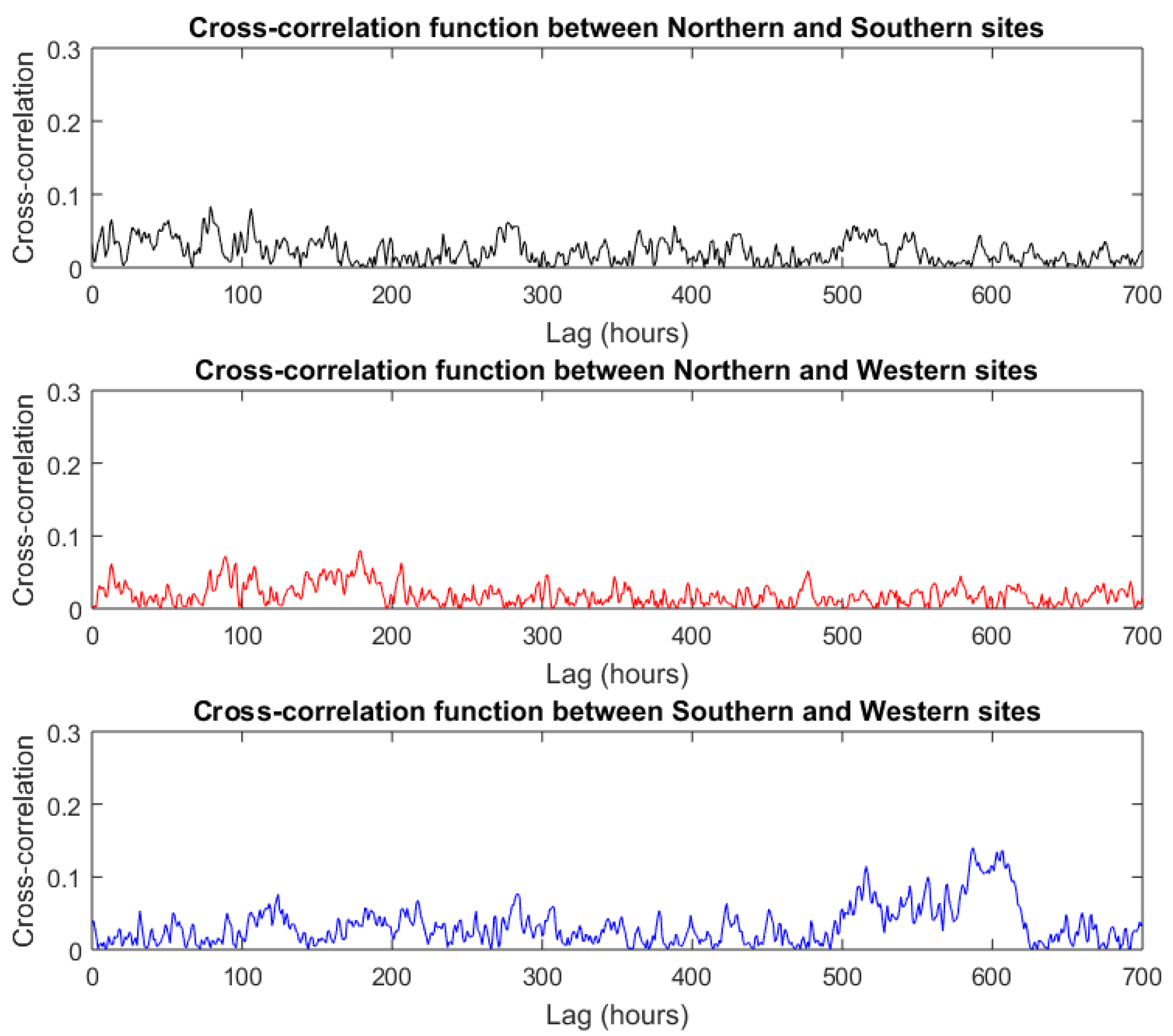
4. Results and Discussion
4.1. Sperm Whale Detections
4.2. Regional Population Density Estimates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaquet, N.; Dawson, S.; Douglas, L. Vocal behavior of male sperm whales: Why do they click? J. Acoust. Soc. Am. 2001, 109, 2254–2259. [Google Scholar] [CrossRef] [PubMed]
- Møhl, B.; Wahlberg, M.; Madsen, P.T.; Heerfordt, A.; Lund, A. The monopulsed nature of sperm whale clicks. J. Acoust. Soc. Am. 2003, 114, 1143–1154. [Google Scholar] [CrossRef]
- Wahlberg, M.; Frantzis, A.; Alexiadou, P.; Madsen, P.T.; Møhl, B. Click production during breathing in a sperm whale (Physeter macrocephalus) (L). J. Acoust. Soc. Am. 2005, 118, 3404–3407. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, W.M.; Tyack, P.L.; Johnson, M.P.; Madsen, P.T. Three-dimensional beam pattern of regular sperm whale clicks confirms bent-horn hypothesis. J. Acoust. Soc. Am. 2005, 117, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Ackleh, A.S.; Ioup, G.E.; Ioup, J.W.; Ma, B.; Newcomb, J.; Pal, N.; Sidorovskaia, N.A.; Tiemann, C. Assessing the Deepwater Horizon oil spill impact on marine mammal population through acoustics: Endangered sperm whales. J. Acoust. Soc. Am. 2012, 131, 2306–2314. [Google Scholar] [CrossRef] [PubMed]
- Baumann-Pickering, S.; McDonald, M.A.; Simonis, A.E.; Berga, A.S.; Merkens, K.P.B.; Oleson, E.M.; Roch, M.A.; Wiggins, S.M.; Rankin, S.; Yack, T.M.; et al. Species-specific beaked whales echolocation signals. J. Acoust. Soc. Am. 2013, 134, 2293–2301. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, J.A.; Baumann-Pickering, S.; Frasier, K.E.; Trickey, J.S.; Merkens, K.P.; Wiggins, S.M.; McDonald, M.A.; Garrison, L.P.; Harris, D.; Marques, T.A.; et al. Passive acoustic monitoring of beaked whale densities in the Gulf of Mexico. Sci. Rep. 2015, 5, 16343. [Google Scholar] [CrossRef]
- Baumann-Pickering, S.; Roch, M.A.; Brownell Jr, R.L.; Simonis, A.E.; McDonald, M.A.; Solsona-Berga, A.; Oleson, E.M.; Wiggins, S.M.; Hildebrand, J.A. Spatio-temporal patterns of beaked whale echolocation signals in the North Parcifc. PLoS ONE 2014, 9, e91383. [Google Scholar] [CrossRef] [PubMed]
- Mellinger, D.K.; Thode, A.M.; Martinez, A. Passive acoustic monitoring of sperm whales in the Gulf of Mexico, with a model of acoustic detection distance. In Proceedings of the Twenty-First Annual Gulf of Mexico Information Transfer Meeting, New Orleans, LA, USA, 1 January 2002; pp. 493–501. [Google Scholar]
- Thode, A.; Mellinger, D.K.; Stienessen, S.; Martinez, A.; Mullin, K. Depth-dependent acoustic features of diving sperm whales (Physeter microcephalus) in the Gulf of Mexico. J. Acoust. Soc. Am. 2002, 112, 308–321. [Google Scholar] [CrossRef]
- Johnson, M.P.; Tyack, P.L. A digital acoustic recording tag for measuring the response of wild marine mammals to sound. IEEE J. Ocean. Eng. 2003, 28, 3–12. [Google Scholar] [CrossRef]
- Zimmer, W.M.X.; Johnson, M.P.; Amico, A.D.; Tyack, P.L. Combining data from a multisensory tag and passive sonar to determine the diving behavior of a sperm whale (Physeter macrocephalus). IEEE J. Ocean. Eng. 2003, 28, 13–28. [Google Scholar] [CrossRef]
- Barlow, J.; Taylor, B.L. Estimates of sperm whale abundance in the northeastern temperate pacific from a combined acoustic and visual survey. Mar. Mamm. Sci. 2005, 21, 429–445. [Google Scholar] [CrossRef]
- Laplanche, C.; Adam, O.; Lopatka, M.; Motsch, J. Male sperm whale acoustic behavior observed from multipaths at a single hydrophone. J. Acoust. Soc. Am. 2005, 118, 2677–2687. [Google Scholar] [CrossRef]
- Morrissey, R.P.; Ward, J.; DiMarzio, N.; Jarvis, S.; Moretti, D.J. Passive acoustic detection and localization of sperm whales (Physeter macrocephalus) in the tongue of the ocean. Appl. Acoust. 2006, 67, 1091–1105. [Google Scholar] [CrossRef]
- Zimmer, W.M.X.; Harwood, J.; Tyack, P.L.; Johnson, M.P.; Madsen, P.T. Passive acoustic detection of deep-diving beaked whales. J. Acoust. Soc. Am. 2008, 124, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.A.; Thomas, L.; Ward, J.; DiMarzio, N.; Tyack, P.L. Estimating cetacean population density using fixed passive acoustic sensors: An example with Blainville’s beaked whales. J. Acoust. Soc. Am. 2009, 125, 1982–1994. [Google Scholar] [CrossRef]
- Mellinger, D.K.; Küsel, E.T.; Thomas, L.; Marques, T.; Moretti, D.; Baggenstoss, P.; Ward, J.J.; DiMarzio, N.; Morrissey, R. Population density of sperm whales in the Bahamas estimated using propagation modeling. J. Acoust. Soc. Am. 2010, 127, 1824. [Google Scholar] [CrossRef]
- Küsel, E.T.; Mellinger, D.K.; Thomas, L.; Marques, T.A.; Moretti, D.; Ward, J. Cetacean population density estimation from single fixed sensors using passive acoustics. J. Acoust. Soc. Am. 2011, 129, 3610–3622. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.A.; Thomas, L.; Jarvis, S.; DiMarzio, N.; Moretti, D.; Marques, T.A.; Tyack, P. Passive acoustic density estimation of sperm whales in the Tongue of the Ocean, Bahamas. Mar. Mamm. Sci. 2012, 28, E444–E455. [Google Scholar] [CrossRef]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.; Tyack, P.L. Estimating animal population density using passive acoustics. Biol. Rev. 2013, 88, 287–309. [Google Scholar] [CrossRef]
- Waring, G.T.; Josephson, E.; Maze-Foley, K.; Rosel, P.E. US Atlantic and Gulf of Mexico Marine Mammal Stock Assessments-2015; NOAA Tech. Memo. NMFS-NE-238; NOAA: Woods Hole, MA, USA, 2016.
- Hildebrand, J.; Merkens, K.; Frasier, K.; Bassett, H.; Baumann-Pickering, S.; Sirovic, A.; Wiggins, S.; McDonald, M.; Marques, T.; Harris, D.; et al. Passive Acoustic Monitoring of Cetaceans in the Northern Gulf of Mexico during 2010–2011. Progress Report for Research Agreement #20105138. July 2012, pp. 1–35. Available online: http://cetus.ucsd.edu/Publications/Reports/HildebrandNRDA-July2012.pdf (accessed on 1 April 2021).
- Hildebrand, J.A.; Frasier, K.E.; Baumann-Pickering, S.; Wiggins, S.M.; Merkens, K.P.; Garrison, L.P.; Soldervilla, M.S.; McDonald, M.A. Assessing seasonality and density from passive acoustic monitoring of signals presumed to be from pygmy and dwarf sperm whales in the Gulf of Mexico. Front. Mar. Sci. 2019, 6, 66. [Google Scholar] [CrossRef]
- Frasier, K.E.; Roch, M.A.; Soldevilla, M.S.; Wiggins, S.M.; Garrison, L.P.; Hildebrand, J.A. Automated classification of dolphin echolocation click types from the Gulf of Mexico. PLoS Comput. Biol. 2017, 13, e1005823. [Google Scholar] [CrossRef]
- Sidorovskaia, N.A.; Ioup, G.E.; Ioup, J.W.; Caruthers, J.W. Acoustic propagation studies for sperm whale phonation analysis during LADC experiments. AIP Conf. Proc. 2004, 728, 288–295. [Google Scholar]
- Sidorovskaia, N.A.; Ioup, G.E.; Ioup, J.W. Studies of waveguide propagation effects on frequency properties of sperm whale communication signals. J. Acoust. Soc. Am. 2004, 116, 2614. [Google Scholar] [CrossRef]
- Ioup, G.E.; Ioup, J.W.; Pflug, L.A.; Tashmukhambetov, A.M.; Sidorovskaia, N.A.; Schexnayder, P.; Tiemann, C.O.; Bernstein, A.; Kuczaj, S.A.; Rayborn, G.H.; et al. EARS Buoy Applications by LADC: I. Marine Mammals. In Proceedings of the OCEANS 2009, Biloxi, MS, USA, 26–29 October 2009; pp. 1–9. [Google Scholar]
- Ioup, G.E.; Ioup, J.W.; Sidorovskaia, N.A.; Walker, R.T.; Kuczaj, S.A.; Walker, C.D.; Fisher, R. Analysis of bottom-moored hydrophone measurements of Gulf of Mexico sperm whale phonations. In Proceedings of the 23rd Annual Gulf of Mexico Information Transfer Meeting, New Orleans, LA, USA, January 2005; U.S. Department of the Interior, Minerals Management Service, Gulf of Mexico OCS Region: New Orleans, LA, USA, 2005; pp. 109–136. [Google Scholar]
- Li, K.; Sidorovskaia, N.A.; Tiemann, C.O. Model-based unsupervised clustering for distinguishing Cuvier’s and Gervais’ beaked whales in acoustic data. Ecol. Inform. 2020, 58, 101094. [Google Scholar] [CrossRef]
- Watwood, S.L.; Miller, P.J.; Johnson, M.; Madsen, P.T.; Tyack, P.L. Deep-diving foraging behavior of sperm whales. J. Anim. Ecol. 2006, 75, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Rhinelander, M.Q.; Dawson, S.M. Measuring sperm whales from their clicks: Stability of interpulse inter-vals and validation that they indicate whale length. J. Acoust. Soc. Am. 2004, 115, 1826–1831. [Google Scholar] [CrossRef]
- Antunes, R.; Rendell, L.; Gordon, J. Measuring inter-pulse intervals in sperm whale clicks: Consistency of automatic estimation methods. J. Acoust. Soc. Am. 2010, 127, 3239–3247. [Google Scholar] [CrossRef]
- Growcott, A.; Miller, B.; Sirguey, P.; Slooten, E.; Dawson, S. Measuring body length of male sperm whales from their clicks: The relationship between inter-pulse intervals and photogrammetrically measured lengths. J. Acoust. Soc. Am. 2011, 130, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.S.; Growcott, A.; Slooten, E.; Dawson, S.M. Acoustically derived growth rates of sperm whales (Physeter macrocephalus) in Kaikoura, New Zealand. J. Acoust. Soc. Am. 2013, 134, 2438–2445. [Google Scholar] [CrossRef]
- Zimmer, W.M.; Madsen, P.T.; Teloni, V.; Johnson, M.P.; Tyack, P.L. Off-axis effects on the multipulse structure of sperm whale usual clicks with implications for sound production. J. Acoust. Soc. Am. 2005, 118, 3337–3345. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.C. Evaluation of a method for determining the length of sperm whales (Physeter catodon) from their vocalizations. J. Zool. Lond. 1991, 224, 301–314. [Google Scholar] [CrossRef]
- Goold, J.C. Signal processing techniques for acoustic measurement of sperm whale body lengths. J. Acoust. Soc. Am. 1996, 100, 3431–3441. [Google Scholar] [CrossRef]
- Pavan, G.; Priano, M.; Manghi, M.; Fossati, C. Software tools for real-time IPI measurements on sperm whale sounds. In Proceedings of the Underwater Bio-Sonar and Bioacoustics Symposium, Loughborough, UK, 16–17 December 1997; Loughborough University: Loughborough, UK, 1997; pp. 157–164. [Google Scholar]
- Caruso, F.; Sciacca, V.; Bellia, G.; De Domenico, E.; Larosa, G.; Papale, E.; Pellegrino, C.; Pulvirenti, S.; Riccobene, G.; Simeone, F.; et al. Size Distribution of Sperm Whales Acoustically Identified During Long Term Deep-Sea Monitoring in the Ionian Sea. PLoS ONE 2015, 10, e0144503. [Google Scholar] [CrossRef]
- Beslin, W.A.M.; Whitehead, H.; Gero, S. Automatic acoustic estimation of sperm whale size distributions achieved through machine recognition of on-axis clicks. J. Acoust. Soc. Am. 2018, 144, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Lohrasbipeydeh, H.; Dakin, D.T.; Gulliver, T.A.; Amindavar, H.; Zielinski, A. Adaptive energy-based acoustic sperm whale echolocation click detection. IEEE J. Ocean. Eng. 2015, 40, 957–968. [Google Scholar] [CrossRef]
- Urick, R.J. Principles of Underwater Sound; McGraw-Hill Book Co.: New York, NY, USA, 1983. [Google Scholar]
- Zimmer, W.M.X.; Johnson, M.P.; Madsen, P.T.; Tyack, P.L. Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). J. Acoust. Soc. Am. 2005, 117, 3919–3927. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.B.; Kuperman, W.A.; Porter, M.B.; Schmidt, H. Computational Ocean Acoustics; Springer Science & Business Media: New York, NY, USA, 2011. [Google Scholar]
- Mathias, D.; Thode, A.M.; Straley, J.; Andrews, R.D. Acoustic tracking of sperm whales in the Gulf of Alaska using a two-element vertical array and tags. J. Acoust. Soc. Am. 2013, 134, 2446–2461. [Google Scholar] [CrossRef] [PubMed]
- Warren, V.E.; Marques, T.A.; Harris, D.; Thomas, L.; Tyack, P.L.; Aguilar de Soto, N.; Hickmott, L.S.; Johnson, M.P. Spatio-temporal variation in click production rates of beaked whales: Implications for passive acoustic density estimation. J. Acoust. Soc. Am. 2017, 141, 1962–1974. [Google Scholar] [CrossRef]
- Whitehead, H.; Weilgart, L. Click rates from sperm whales. J. Acoust. Soc. Am. 1990, 87, 1798–1806. [Google Scholar] [CrossRef]
- Goold, J.C.; Jones, S.E. Time and frequency domain characteristics of sperm whale clicks. J. Acoust. Soc. Am. 1995, 98, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Fais, A.; Johnson, M.; Wilson, M.; Aguilar Soto, N.; Madsen, P.T. Sperm whale predator-prey interactions involve chasing and buzzing, but no acoustic stunning. Sci. Rep. 2016, 6, 28562. [Google Scholar] [CrossRef] [PubMed]
- Winsor, M.H.; Irvine, L.M.; Mate, B.R. Analysis of the Spatial Distribution of Satellite-Tagged Sperm Whales (Physeter macrocephalus) in Close Proximity to Seismic Surveys in the Gulf of Mexico. Aquat. Mamm. 2017, 43, 439–446. [Google Scholar] [CrossRef]
- Sullivan, L.; Brosnan, T.; Rowles, T.; Schwacke, L.; Simeone, C.; Collier, T.K. Guidelines for Assessing Exposure and Impacts of Oil Spills on Marine Mammals; NOAA Tech. Memo. NMFS-OPR-62; NOAA: Woods Hole, MA, USA, 2019.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year/Station | Latitude (N) | Longitude (W) | Water Depth (m) | Start | End |
|---|---|---|---|---|---|
| 2007 North | 28°38.99′ | 88°31.56′ | 1560 | 6 July | 16 July |
| 2007 South | 28°25.23′ | 88°37.07′ | 1465 | 7 July | 16 July |
| 2010 North | 28°39.00′ | 88°31.53′ | 1545 | 10 Sept | 22 Sept |
| 2010 South | 28°24.61′ | 88°34.26′ | 1550 | 11 Sept | 23 Sept |
| 2010 West | 28°23.06′ | 88°59.52′ | 1000 | 12 Sept | 24 Sept |
| 2015 North | 28°39.07′ | 88°31.05′ | 1570 | 26 June | 22 Oct |
| 2015 South | 28°25.28′ | 88°37.11′ | 1500 | 25 June | 21 Oct |
| 2015 West | 28°24.04′ | 88°59.69′ | 1000 | 24 June | 21 Oct |
| 2016 South | 28°27.71′ | 88°36.30′ | 1500 | 10 Sept | 30 Jan |
| 2017 North | 28°38.09′ | 88°33.17′ | 1500 | 09 June | 09 Oct |
| 2017 South | 28°27.63′ | 88°36.16′ | 1500 | 11 June | 09 Oct |
| 2017 West | 28°23.84′ | 88°59.23′ | 1000 | 10 June | 09 Oct |
| Parameter | Value |
|---|---|
| Reference source level | 235 1 |
| (dB re 1 µPa) | |
| Piston radius a (cm) | 80 1 |
| Sound speed c (m/s) | 1500 |
| Frequency range f (kHz) | 3–20 |
| Center frequency f0 (kHz) | 13.4 1 |
| RMS bandwidth b (kHz) | 4.1 1 |
| Year | Northern Site | Southern Site | Western Site | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2007 | 421.72 | 0.090 | 0.106 | 205.32 | 0.217 | 0.108 | n/a | n/a | n/a |
| 2010 | 85.49 | 0.156 | 0.081 | 316.60 | 0.071 | 0.056 | 232.43 | 0.071 | 0.055 |
| 2015 | 159.81 | 0.115 | 0.085 | 121.24 | 0.117 | 0.072 | 189.91 | 0.137 | 0.054 |
| 2016 | n/a | n/a | n/a | 264.70 | 0.070 | 0.067 | n/a | n/a | n/a |
| 2017 | 288.54 | 0.120 | 0.072 | 231.90 | 0.080 | 0.056 | 213.94 | 0.069 | 0.062 |
| Year | Northern Site | Southern Site | Western Site | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ICI | Std | ICI | Std | ICI | Std | ||||
| 2007 | 0.7017 | 0.4160 | 1.1396 | 0.7085 | 0.4186 | 1.1286 | n/a | n/a | n/a |
| 2010 | 0.7373 | 0.4397 | 1.0845 | 0.7169 | 0.4236 | 1.1154 | 0.7439 | 0.4074 | 1.0749 |
| 2015 | 0.7697 | 0.4288 | 1.0389 | 0.7924 | 0.4152 | 1.0091 | 0.7488 | 0.4214 | 1.0679 |
| 2016 | n/a | n/a | n/a | 0.7492 | 0.4138 | 1.0673 | n/a | n/a | n/a |
| 2017 | 0.7516 | 0.4276 | 1.0639 | 0.7481 | 0.4236 | 1.0689 | 0.7349 | 0.4222 | 1.0881 |
| Year | Northern Site | Southern Site | Western Site | |||
|---|---|---|---|---|---|---|
(/1000 km2) | CV | (/1000 km2) | CV | (/1000 km2) | CV | |
| 2007 | 0.599 | 0.021 | 0.249 | 0.034 | n/a | n/a |
| 2010 | 0.128 | 0.068 | 0.888 | 0.027 | 0.698 | 0.028 |
| 2015 | 0.302 | 0.010 | 0.275 | 0.013 | 0.518 | 0.009 |
| 2016 | n/a | n/a | 0.651 | 0.009 | n/a | n/a |
| 2017 | 0.640 | 0.008 | 0.671 | 0.011 | 0.536 | 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Sidorovskaia, N.A.; Guilment, T.; Tang, T.; Tiemann, C.O. Decadal Assessment of Sperm Whale Site-Specific Abundance Trends in the Northern Gulf of Mexico Using Passive Acoustic Data. J. Mar. Sci. Eng. 2021, 9, 454. https://doi.org/10.3390/jmse9050454
Li K, Sidorovskaia NA, Guilment T, Tang T, Tiemann CO. Decadal Assessment of Sperm Whale Site-Specific Abundance Trends in the Northern Gulf of Mexico Using Passive Acoustic Data. Journal of Marine Science and Engineering. 2021; 9(5):454. https://doi.org/10.3390/jmse9050454
Chicago/Turabian StyleLi, Kun, Natalia A. Sidorovskaia, Thomas Guilment, Tingting Tang, and Christopher O. Tiemann. 2021. "Decadal Assessment of Sperm Whale Site-Specific Abundance Trends in the Northern Gulf of Mexico Using Passive Acoustic Data" Journal of Marine Science and Engineering 9, no. 5: 454. https://doi.org/10.3390/jmse9050454
APA StyleLi, K., Sidorovskaia, N. A., Guilment, T., Tang, T., & Tiemann, C. O. (2021). Decadal Assessment of Sperm Whale Site-Specific Abundance Trends in the Northern Gulf of Mexico Using Passive Acoustic Data. Journal of Marine Science and Engineering, 9(5), 454. https://doi.org/10.3390/jmse9050454