
Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea


Abstract
1. Introduction
2. Materials and Methods
2.1. Cruise Information
2.2. Survey Stations and Zones
2.3. Hydrological Data
2.4. Chlorophyll a and Nutrients
2.5. Phytoplankton Community
2.6. Meteorological Data
2.7. Statistical Analysis
3. Results
3.1. Distribution of Surface Temperature and Salinity
3.2. Meteorological Features
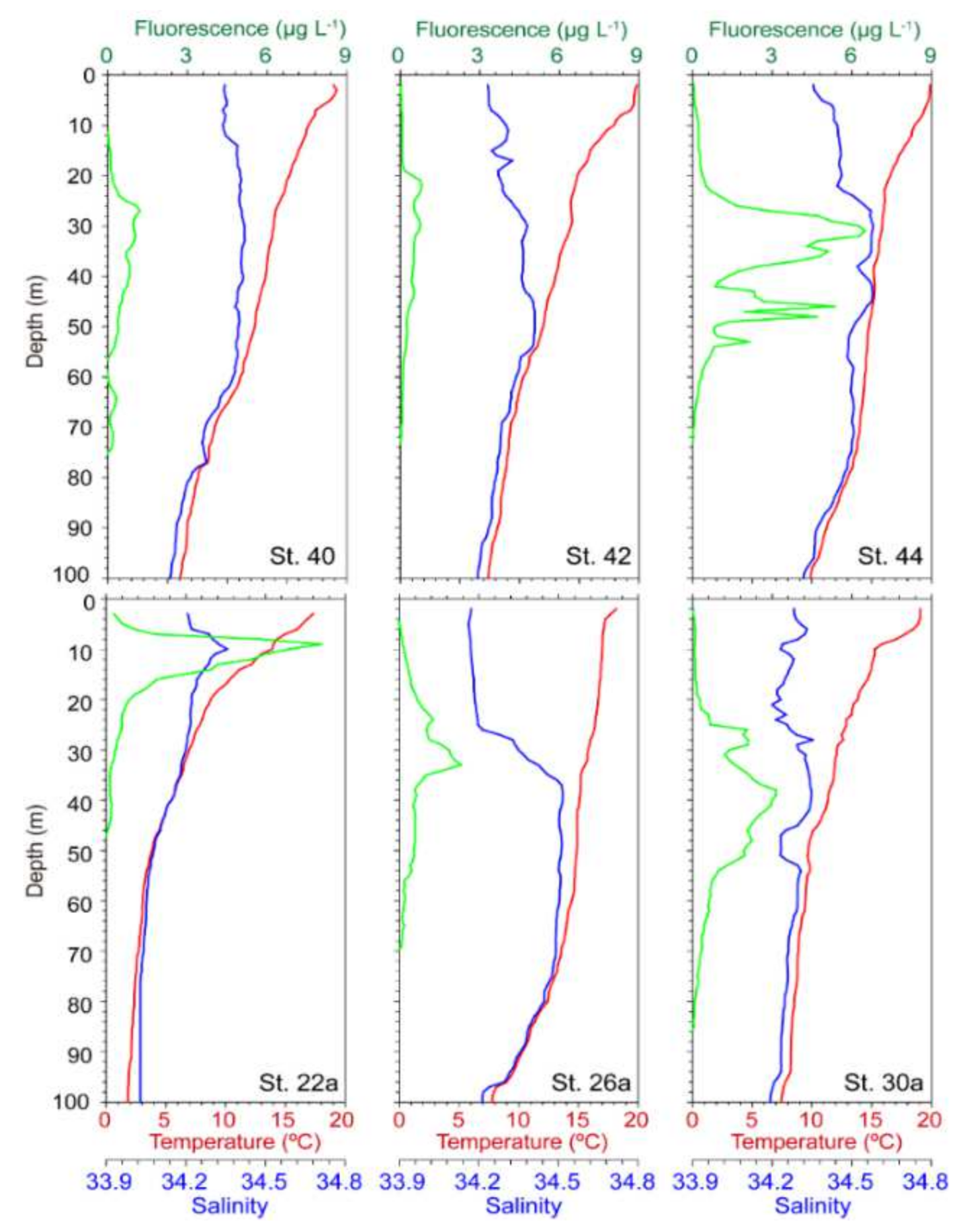
3.3. Vertical Profiles
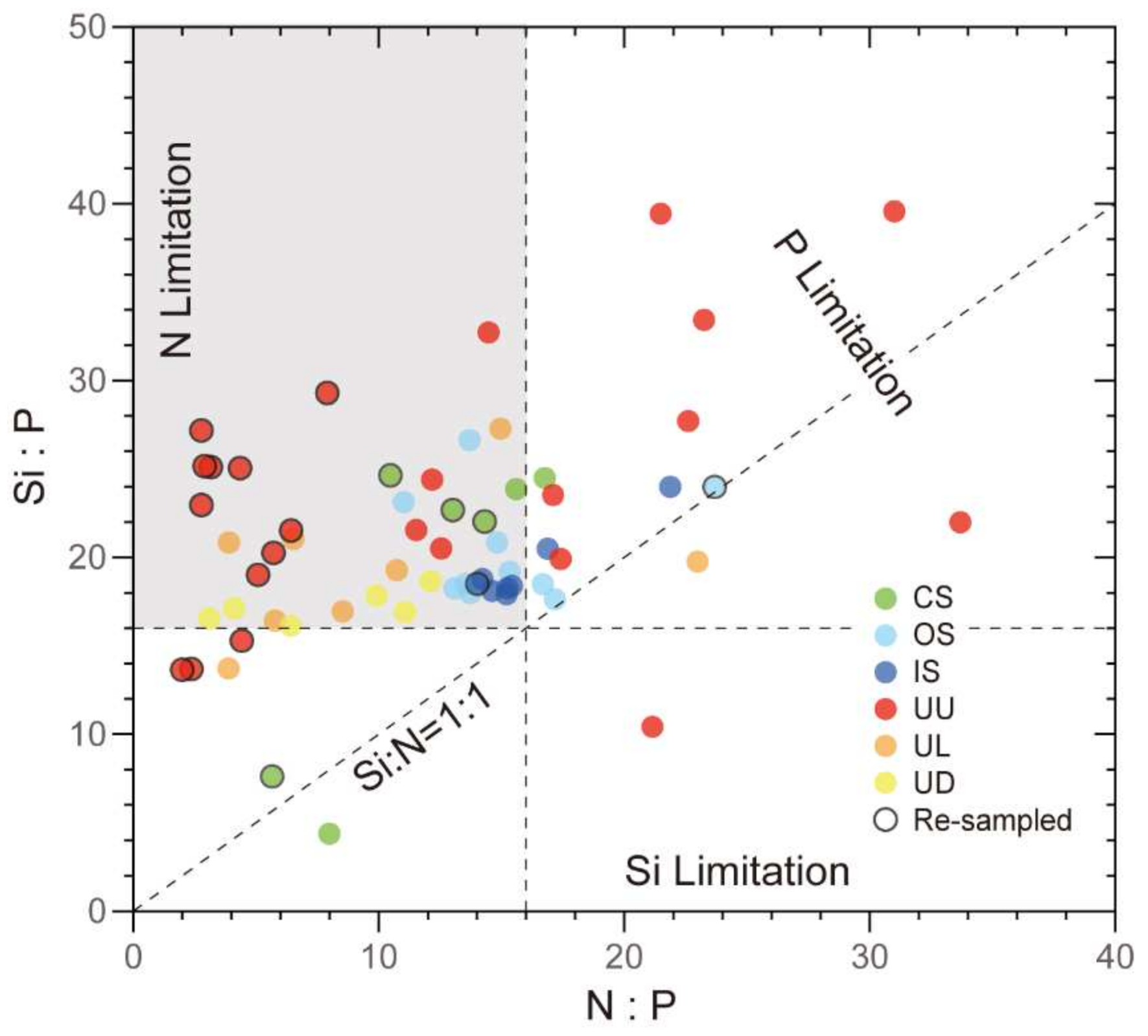
3.4. Distribution of Nutrients
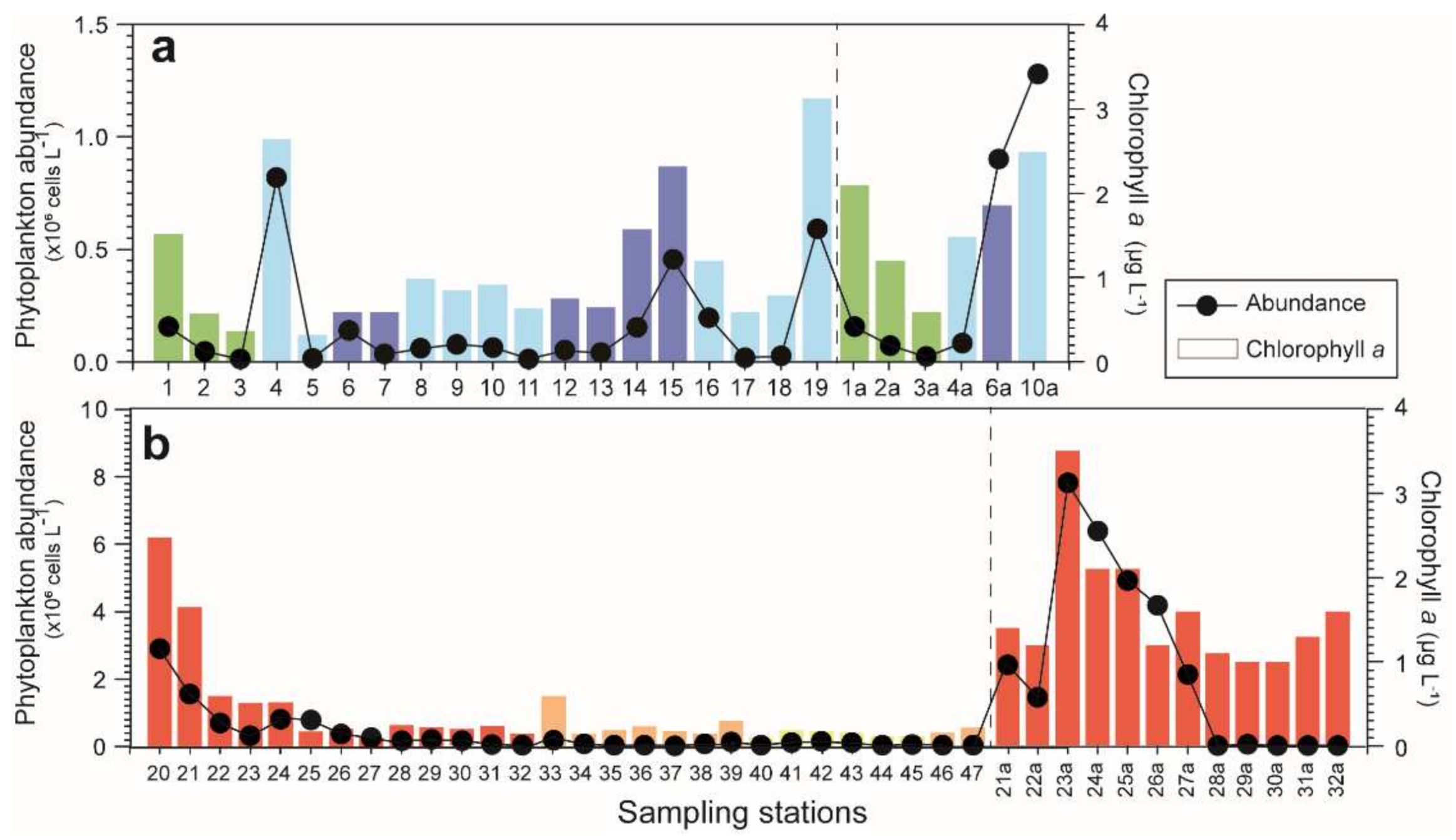
3.5. Chlorophyll a
3.6. Phytoplankton Abundance
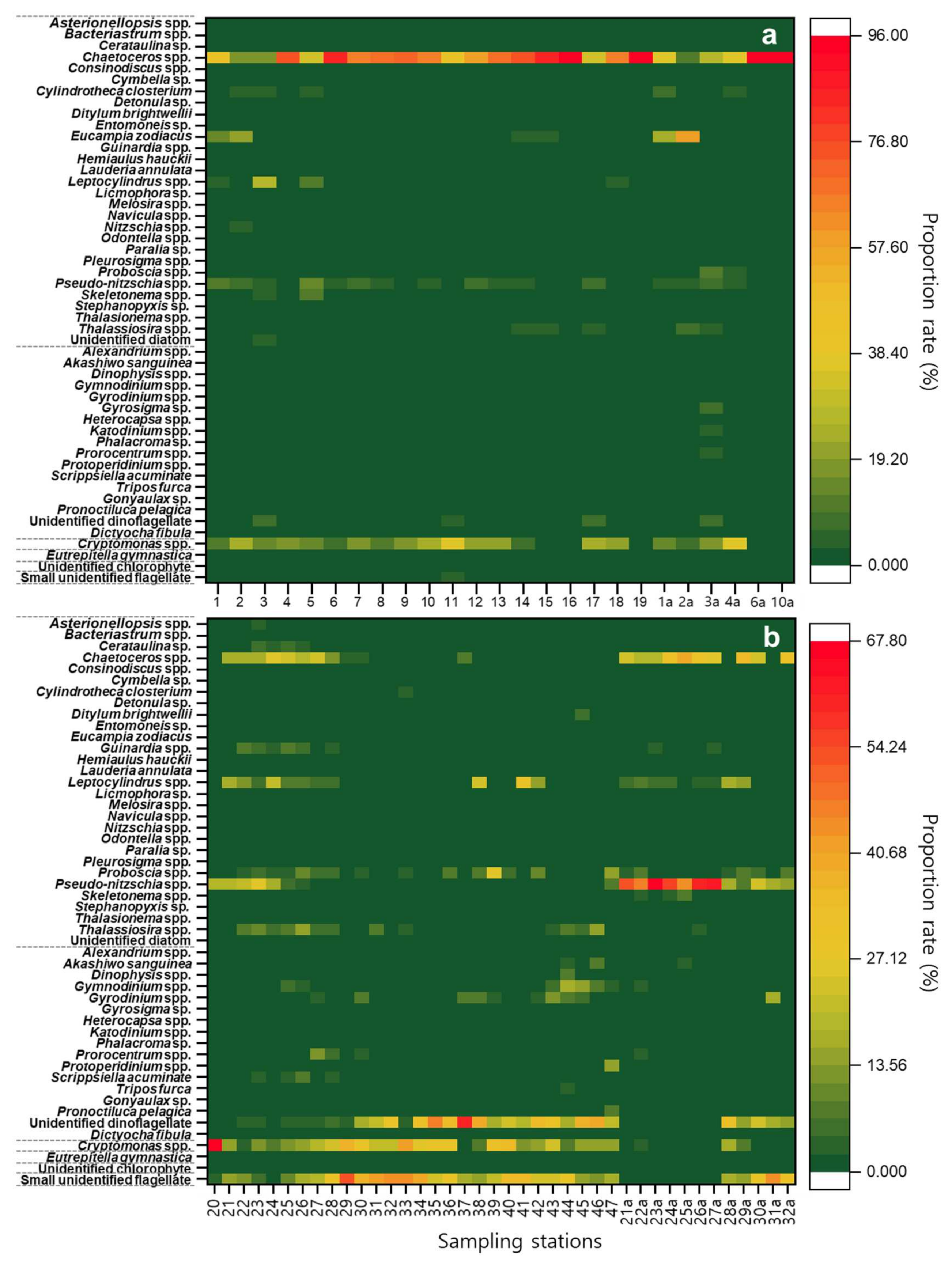
3.7. Phytoplankton Community
3.8. Comparison of Environment in Early Spring and Late Spring
3.9. Canonical Correspondence Analysis
4. Discussion
4.1. Environmental Characteristics and Phytoplankton Dynamics during Early Spring
4.2. Environmental Characteristics and Phytoplankton Dynamics during Late Spring
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lancelot, C.; Billen, G.; Sournia, A.; Weisse, T.; Colijn, F.; Veldhuis, M.J.W.; Davies, A.; Wassman, P. Phaeocystis Blooms and Nutrient Enrichment in the Continental Coastal Zones of the North Sea. Ambio 1987, 16, 38–46. [Google Scholar]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Anderson, G.C.; Zeutschel, R.P. Release of dissolved organic matter by marine phytoplankton in coastal and offshore areas of the Northeast Pacific ocean. Limnol. Oceanogr. 1970, 15, 402–407. [Google Scholar] [CrossRef]
- Hyun, J.-H.; Kim, D.; Shin, C.-W.; Noh, J.-H.; Yang, E.-J.; Mok, J.-S.; Kim, S.-H.; Kim, H.-C.; Yoo, S. Enhanced phytoplankton and bacterioplankton production coupled to coastal upwelling and an anticyclonic eddy in the Ulleung basin, East Sea. Aquat. Microb. Ecol. 2009, 54, 45–54. [Google Scholar] [CrossRef]
- Isobe, A.; Isoda, Y. Circulation in the Japan Basin, the Northern Part of the Japan Sea. J. Oceanogr. 1997, 53, 373. [Google Scholar]
- Isoda, Y.; Saitoh, S. The northward intruding eddy along the East coast of Korea. J. Oceanogr. 1993, 49, 443–458. [Google Scholar] [CrossRef]
- Gong, Y.; Son, S. The study of the study on the marine thermo-front in the east sea of Korea. Rep. Nat. Fish. Res. Dep. Inst. 1982, 28, 24–25. [Google Scholar]
- Martin, S.; Kawase, M. The southern flux of sea ice in the Tatarskiy Strait, Japan Sea and the generation of the Liman Current. J. Mar. Res. 1998, 56, 141–155. [Google Scholar] [CrossRef]
- Chung, C.S.; Shim, J.H.; Park, Y.G.; Park, S.-G. Primary Productivity and Nitrogenous Nutrient Dynamics in the East Sea of Korea. J. Oceanol. Soc. Korea 1989, 24, 52–61. [Google Scholar]
- Moon, C.-H.; Yang, S.-R.; Yang, H.-S.; Cho, H.-J.; Lee, S.-Y.; Kim, S.-Y. Regeneration Processes of Nutrients in the Polar Front Area of the last Sea IV. Chlorophyll a Distribution, New Production and the Vertical Diffusion of Nitrate. Korean J. Fish. Aquat. Sci. 1998, 31, 259–266. [Google Scholar]
- Kwak, J.H.; Hwang, J.; Choy, E.J.; Park, H.J.; Kang, D.J.; Lee, T.; Chang, K.I.; Kim, K.R.; Kang, C.K. High primary productivity and f-ratio in summer in the Ulleung basin of the East/Japan Sea. Deep Sea Res. Part. I Oceanogr. Res. Pap. 2013, 79, 74–85. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, D.J.; Kim, I.N.; Rho, T.; Lee, T.; Kang, C.K.; Kim, K.R. Spatial and temporal variability in the pelagic ecosystem of the East Sea (Sea of Japan): A review. J. Mar. Syst. 2009, 78, 288–300. [Google Scholar] [CrossRef]
- Yoo, S.; Park, J. Why is the southwest the most productive region of the East Sea/Sea of Japan? J. Mar. Syst. 2009, 78, 301–315. [Google Scholar] [CrossRef]
- Ji, R.; Jin, M.; Li, Y.; Kang, Y.H.; Kang, C.K. Variability of primary production among basins in the East/Japan Sea: Role of water column stability in modulating nutrient and light availability. Prog. Oceanogr. 2019, 178, 102173. [Google Scholar] [CrossRef]
- Lie, H.J.; Cho, C.H. Seasonal circulation patterns of the Yellow and East China Seas derived from satellite-tracked drifter trajectories and hydrographic observations. Prog. Oceanogr. 2016, 146, 121–141. [Google Scholar] [CrossRef]
- Ryan, J.P.; Polito, P.S.; Strutton, P.G.; Chavez, F.P. Unusual large-scale phytoplankton blooms in the equatorial Pacific. Prog. Oceanogr. 2002, 55, 263–285. [Google Scholar] [CrossRef]
- Zhao, H.; Tang, D.; Wang, Y. Comparison of phytoplankton blooms triggered by two typhoons with different intensities and translation speeds in the South China Sea. Mar. Ecol. Prog. Ser. 2008, 365, 57–65. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Yoshiki, T.; Nakajima, R.; Miyaguchi, H.; Kuwahara, V.S.; Taguchi, S.; Kikuchi, T.; Toda, T. Typhoon-driven variations in primary production and phytoplankton assemblages in Sagami Bay, Japan: A case study of typhoon Mawar (T0511). Plankt. Benthos Res. 2013, 8, 74–87. [Google Scholar] [CrossRef][Green Version]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Baek, S.H.; Shimode, S.; Kim, H.C.; Han, M.S.; Kikuchi, T. Strong bottom–up effects on phytoplankton community caused by a rainfall during spring and summer in Sagami Bay, Japan. J. Mar. Syst. 2009, 75, 253–264. [Google Scholar] [CrossRef]
- Baek, S.H.; Kim, D.; Son, M.; Yun, S.M.; Kim, Y.O. Seasonal distribution of phytoplankton assemblages and nutrient-enriched bioassays as indicators of nutrient limitation of phytoplankton growth in Gwangyang Bay, Korea. Estuar. Coast. Shelf Sci. 2015, 163, 265–278. [Google Scholar] [CrossRef]
- Lee, M.; Kim, Y.-B.; Kang, J.H.; Park, C.H.; Baek, S.H. Seasonal phytoplankton dynamics in oligotriphic offshore water of Dokdo, 2018. Korean J. Environ. Biol. 2019, 37, 19–30. [Google Scholar] [CrossRef]
- Kim, H.C.; Yoo, S.; Oh, I.S. Relationship between phytoplankton bloom and wind stress in the sub-polar frontal area of the Japan/East Sea. J. Mar. Syst. 2007, 67, 205–216. [Google Scholar] [CrossRef]
- Lee, M.; Kim, J.H.; Kim, Y.-B.; Park, C.H.; Shin, K.; Baek, S.H. Specific oceanographic characteristics and phytoplankton responses influencing the primary production around the Ulleung Basin area in spring. Acta Oceanol. Sin. 2020, 39, 107–122. [Google Scholar] [CrossRef]
- Kang, Y.-S.; Choi, H.-C.; Lim, J.-H.; Jeon, I.-S.; Seo, J.-H. Dynamics of the Phytoplankton Community in the Coastal Waters of Chuksan Harbor, East Sea. Algae 2005, 20, 345–352. [Google Scholar] [CrossRef]
- Oh, H.J.; Suh, Y.S.; Heo, S. The Relationship Between Phytoplankton Distribution and Environmental Conditions of the Upwelling Cold Water in the Eastern Coast of the Korean Peninsula. J. Korean Assoc. Geogr. Inf. Stud. 2004, 7, 166–173. [Google Scholar]
- Baek, S.H.; Lee, M.; Kim, Y.B. Spring phytoplankton community response to an episodic windstorm event in oligotrophic waters offshore from the Ulleungdo and Dokdo islands, Korea. J. Sea Res. 2018, 132, 1–14. [Google Scholar] [CrossRef]
- Baek, S.H.; Lee, M.; Park, B.S.; Lim, Y.K. Variation in Phytoplankton Community Due to an Autumn Typhoon and Winter Water Turbulence in Southern Korean Coastal Waters. Sustainability 2020, 12, 2781. [Google Scholar] [CrossRef]
- Ahn, Y.-H.; Shanmugam, P.; Ryu, J.-H.; Jeong, J.-C. Satellite detection of harmful algal bloom occurrences in Korean waters. Harmful Algae 2006, 5, 213–231. [Google Scholar] [CrossRef]
- Kim, H.-C.; Yamaguchi, H.; Yoo, S.; Zhu, J.; Okamura, K.; Kiyomoto, Y.; Tanaka, K.; Kim, S.-W.; Park, T.; Oh, I.S.; et al. Distribution of Changjiang Diluted Water detected by satellite chlorophyll a and its interannual variation during 1998–2007. J. Oceanogr. 2009, 65, 129–135. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical & Biological Methods for Seawater Analysis; Elsevier: Amsterdam, The Netherlands, 1984; ISBN 978-0-08-030287-4. [Google Scholar]
- Sournia, A. Phytoplankton Manual; United Nations Educational, Scientific and Cultural Organization: Paris, France, 1978; ISBN 92-3-101572-9. [Google Scholar]
- Baek, S.H.; Kim, D.; Kim, Y.O.; Son, M.; Kim, Y.J.; Lee, M.; Park, B.S. Seasonal changes in abiotic environmental conditions in the Busan coastal region (South Korea) due to the Nakdong River in 2013 and effect of these changes on phytoplankton communities. Cont. Shelf Res. 2019, 175, 116–126. [Google Scholar] [CrossRef]
- Chung, C.-C.; Gong, G.-C.; Hung, C.-C. Effect of Typhoon Morakot on microphytoplankton population dynamics in the subtropical Northwest Pacific. Mar. Ecol. Prog. Ser. 2012, 448, 39–49. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Kuwahara, V.S.; Yoshiki, T.; Nakajima, R.; Miyaguchi, H.; Kumekawa, N.; Kikuchi, T.; Toda, T. Phytoplankton community response and succession in relation to typhoon passages in the coastal waters of Japan. J. Plankton Res. 2014, 36, 424–438. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E. Marine Diatoms. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 5–386. [Google Scholar]
- Degerlund, M.; Huseby, S.; Zingone, A.; Sarno, D.; Landfald, B. Functional diversity in cryptic species of Chaetoceros socialis Lauder (Bacillariophyceae). J. Plankton Res. 2012, 34, 416–431. [Google Scholar] [CrossRef]
- Gaonkar, C.C.; Kooistra, W.H.C.F.; Lange, C.B.; Montresor, M.; Sarno, D. Two new species in the Chaetoceros socialis complex (Bacillariophyta): C. sporotruncatus and C. dichatoensis, and characterization of its relatives, C. radicans and C. cinctus. J. Phycol. 2017, 53, 889–907. [Google Scholar] [CrossRef]
- Lee, J.B.; Jwa, J.; Kang, D.; Go, Y.B.; Oh, B. Bioecological Characteristics of Coral Habitats around Moonsom, Cheju Island, Korea: Ⅱ. Community Dynamics of Phytoplankton and Primary Productivity. ALGAE 2000, 15, 37. [Google Scholar]
- Kim, H.-J.; Park, J.Y.; Son, M.H.; Moon, C.-H. Long-term Variations of Phytoplankton Community in Coastal Waters of Kyoungju City Area. J. Fish. Mar. Sci. Educ. 2016, 28, 1417–1434. [Google Scholar] [CrossRef][Green Version]
- Kang, Y.; Choi, J. Ecological characteristics of phytoplankton communities in the coastal waters of Gori, Wulseong, Uljin and Youngkwang I. Species Compostion and Distribution (1992~1996). Algae 2001, 16, 85. [Google Scholar]
- Nishikawa, T.; Hori, Y.; Tanida, K.; Imai, I. Population dynamics of the harmful diatom Eucampia zodiacus Ehrenberg causing bleachings of Porphyra thalli in aquaculture in Harima-Nada, the Seto Inland Sea, Japan. Harmful Algae 2007, 6, 763–773. [Google Scholar] [CrossRef]
- Rumyantseva, A.; Henson, S.; Martin, A.; Thompson, A.F.; Damerell, G.M.; Kaiser, J.; Heywood, K.J. Phytoplankton spring bloom initation: The impact of the atmospheric forcing anf light in the temperate North Atlantic Ocean. Prog. Oceanogr. 2019, 178, 102202. [Google Scholar] [CrossRef]
- Lim, Y.K.; Park, B.S.; Kim, J.H.; Baek, S.S.; Baek, S.H. Effect of marine heatwaves on bloom formation of the harmful dinoflagellate Cochlodinium polykrikoides: Two sides of the same coin? Harmful Algae 2021, 104, 102029. [Google Scholar] [CrossRef]
- Onitsuka, G.; Yanagi, T.; Yoon, J.H. A numerical study on nutrient sources in the surface layer of the Japan Sea using a coupled physical-ecosystem model. J. Geophys. Res. Oceans 2007, 112. [Google Scholar] [CrossRef]
- Kwak, J.H.; Lee, S.H.; Park, H.J.; Choy, E.J.; Jeong, H.D.; Kim, K.R.; Kang, C.K. Monthly measured primary and new productivities in the Ulleung Basin as a biological “hot spot” in the East/Japan Sea. Biogeosciences 2013, 10, 4405–4417. [Google Scholar] [CrossRef]
- Lee, T.; Rho, T.K. Seawater N/P ratio of the East Sea. Sea 2015, 20, 199–205. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 230A-221. [Google Scholar]
- Gilmartin, M.; Revelante, N. The ‘island mass’ effect on the phytoplankton and primary production of the Hawaiian Islands. J. Exp. Mar. Bio. Ecol. 1974, 16, 181–204. [Google Scholar] [CrossRef]
- Palacios, D.M. Factors influencing the island-mass effect of the Galápagos Archipelago. Geophys. Res. Lett. 2002, 29, 49-1–49-4. [Google Scholar] [CrossRef]
- Hasegawa, D.; Yamazaki, H.; Ishimaru, T.; Nagashima, H.; Koike, Y. Apparent phytoplankton bloom due to island mass effect. J. Mar. Syst. 2008, 69, 238–246. [Google Scholar] [CrossRef]
- Hernández-León, S. Accumulation of mesozooplankton in a wake area as a causative mechanism of the “island-mass effect”. Mar. Biol. 1991, 109, 141–147. [Google Scholar] [CrossRef]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Jang, P.-G.; Shin, H.; Baek, S.; Jang, M.; Lee, T.; Shin, K. Nutrient distribution and effects on phytoplankton assemblages in the western Korea/Tsushima Strait. N. Z. J. Mar. Freshw. Res. 2013, 47, 21–37. [Google Scholar] [CrossRef]
- Du, X.; Peterson, W.T. Seasonal Cycle of Phytoplankton Community Composition in the Coastal Upwelling System Off Central Oregon in 2009. Estuaries Coasts 2014, 37, 299–311. [Google Scholar] [CrossRef]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Du, X.; Peterson, W.; Fisher, J.; Hunter, M.; Peterson, J. Initiation and development of a toxic and persistent Pseudo-nitzschia bloom off the Oregon coast in spring/summer 2015. PLoS ONE 2016, 11, e0163977. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early Spring | CS | OS | IS | F-Value |
| Temperature (°C) | 11.10 ± 3.88 a | 8.88 ± 0.68 a | 9.17 ± 0.84 a | 2.437 N.S |
| Salinity | 34.39 ± 0.70 a | 34.51 ± 0.09 a | 34.47 ± 0.18 a | 0.248 N.S |
| Dissolved oxygen (mg L−1) | 8.92 ± 1.09 a | 8.74 ± 0.48 a | 8.66 ± 0.32 a | 0.219 N.S |
| Nitrate + Nitrite (μM) | 3.79 ± 2.68 a | 5.38 ± 1.11 a | 5.78 ± 1.04 a | 2.140 N.S |
| Ammonium (μM) | 1.12 ± 0.25 a | 0.61 ± 0.51 a | 0.64 ± 0.41 a | 1.536 N.S |
| Phosphate (μM) | 0.34 ± 0.12 a | 0.40 ± 0.06 a | 0.40 ± 0.05 a | 1.347 N.S |
| Silicate (μM) | 6.81 ± 5.19 a | 7.98 ± 0.95 a | 7.60 ± 1.03 a | 0.384 N.S |
| Chl a (μg L−1) | 0.82 ± 0.61 a | 1.20 ± 0.92 a | 0.98 ± 0.83 a | 0.281 N.S |
| Phytoplankton abundance (×105 cells L−1) | 0.71 ± 0.77 a | 1.89 ± 2.83 a | 1.46 ± 1.60 a | 0.308 N.S |
| Late Spring | UU | UL | UD | F-Value |
| Temperature (°C) | 18.56 ± 0.80 a | 17.95 ± 1.64 a | 18.34 ± 0.57 a | 0.800 N.S |
| Salinity | 34.27 ± 0.09 a | 34.33 ± 0.22 ab | 34.46 ± 0.04 b | 3.720 * |
| Dissolved oxygen (mg L−1) | 7.93 ± 0.25 a | 7.83 ± 0.21 a | 7.79 ± 0.10 a | 1.071 N.S |
| Nitrate + Nitrite (μM) | 1.87 ± 1.38 a | 1.03 ± 0.60 a | 0.87 ± 0.53 a | 2.655 N.S |
| Ammonium (μM) | 0.95 ± 1.88 a | 0.47 ± 0.41 a | 0.20 ± 0.07 a | 0.754 N.S |
| Phosphate (μM) | 0.13 ± 0.13 a | 0.15 ± 0.05 a | 0.14 ± 0.01 a | 0.090 N.S |
| Silicate (μM) | 3.16 ± 1.77 a | 3.30 ± 1.24 a | 2.41 ± 0.26 a | 0.797 N.S |
| Chl a (μg L−1) | 0.58 ± 0.70 a | 0.25 ± 0.14 a | 0.16 ± 0.04 a | 1.922 N.S |
| Phytoplankton abundance (×105 cells L−1) | 7.17 ± 9.28 b | 0.42 ± 0.80 a | 0.43 ± 0.18 a | 3.834 * |
| Early Spring | Day 1 | Day 7 | t-Value |
| Temperature (°C) | 9.72 ± 2.91 | 10.21 ± 0.80 | −0.496 N.S |
| Salinity | 34.45 ± 0.45 | 34.50 ± 0.12 | −0.285 N.S |
| Dissolved oxygen (mg L−1) | 8.97 ± 0.69 | 8.31 ± 0.43 | 2.036 N.S |
| Nitrate + Nitrite (μM) | 4.76 ± 2.11 | 4.23 ± 1.78 | 0.353 N.S |
| Ammonium (μM) | 0.98 ± 0.40 | 0.72 ± 0.70 | 0.691 N.S |
| Phosphate (μM) | 0.36 ± 0.08 | 0.35 ± 0.06 | 0.115 N.S |
| Silicate (μM) | 7.84 ± 3.48 | 7.39 ± 3.10 | 0.219 N.S |
| Chl a (μg L−1) | 1.00 ± 0.95 | 1.62 ± 0.68 | −0.998 N.S |
| Phytoplankton abundance (105 cells L−1) | 2.05 ± 3.05 | 4.18 ± 5.35 | −0.746 * |
| Late Spring | Day 1 | Day 6 | t-Value |
| Temperature (°C) | 18.52 ± 0.82 | 18.83 ± 0.57 | −0.873 N.S |
| Salinity | 34.28 ± 0.07 | 34.43 ± 0.10 | −3.723 ** |
| Dissolved oxygen (mg L−1) | 7.98 ± 0.15 | 8.04 ± 0.36 | −0.431 N.S |
| Nitrate + Nitrite (μM) | 1.61 ± 1.08 | 0.52 ± 0.19 | 3.592 ** |
| Ammonium (μM) | 0.43 ± 0.29 | 0.23 ± 0.17 | 1.777 N.S |
| Phosphate (μM) | 0.09 ± 0.02 | 0.18 ± 0.02 | −10.12 *** |
| Silicate (μM) | 2.93 ± 1.64 | 3.82 ± 0.96 | −1.677 N.S |
| Chl a (μg L−1) | 0.41 ± 0.42 | 1.16 ± 1.45 | −1.728 N.S |
| Phytoplankton abundance (105 cells L−1) | 5.11 ± 5.85 | 24.44 ± 27.57 | −2.69 * |
| Early Spring | Late Spring | t-Value | |
|---|---|---|---|
| Temperature (°C) | 8.95 ± 1.01 | 18.2 ± 1.17 | −13.40 *** |
| Salinity | 34.5 ± 0.09 | 34.4 ± 0.08 | 2.011 N.S |
| Dissolved oxygen (mg L−1) | 8.86 ± 0.18 | 7.81 ± 0.20 | 12.14 *** |
| Nitrate + Nitrite (μM) | 6.17 ± 0.43 | 0.8 ± 0.48 | 19.84 *** |
| Ammonium (μM) | 0.82 ± 0.47 | 0.30 ± 0.21 | 3.286 * |
| Phosphate (μM) | 0.43 ± 0.03 | 0.14 ± 0.01 | 26.00 *** |
| Silicate (μM) | 8.32 ± 0.41 | 2.43 ± 0.32 | 41.48 *** |
| Chl a (μg L−1) | 0.67 ± 0.31 | 0.18 ± 0.06 | 4.329 ** |
| Phytoplankton abundance (104 cells L−1) | 6.12 ± 3.97 | 4.08 ± 1.64 | 1.052 N.S |
| Diatom (104 cells L−1) | 5.00 ± 3.82 | 0.81 ± 0.67 | 3.196 * |
| Dinoflagellate (104 cells L−1) | 0.12 ± 0.06 | 1.56 ± 0.64 | −5.684 *** |
| Cryptophyte (104 cells L−1) | 0.96 ± 0.38 | 0.76 ± 0.54 | 0.364 N.S |
| Others (104 cells L−1) | 0.05 ± 0.03 | 0.95 ± 0.54 | −5.974 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Ro, H.; Kim, Y.-B.; Park, C.-H.; Baek, S.-H. Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea. J. Mar. Sci. Eng. 2021, 9, 1455. https://doi.org/10.3390/jmse9121455
Lee M, Ro H, Kim Y-B, Park C-H, Baek S-H. Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea. Journal of Marine Science and Engineering. 2021; 9(12):1455. https://doi.org/10.3390/jmse9121455
Chicago/Turabian StyleLee, Minji, Hyejoo Ro, Yun-Bae Kim, Chan-Hong Park, and Seung-Ho Baek. 2021. "Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea" Journal of Marine Science and Engineering 9, no. 12: 1455. https://doi.org/10.3390/jmse9121455
APA StyleLee, M., Ro, H., Kim, Y.-B., Park, C.-H., & Baek, S.-H. (2021). Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea. Journal of Marine Science and Engineering, 9(12), 1455. https://doi.org/10.3390/jmse9121455