A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens


Abstract
1. Introduction
2. Effects of Various Seaweeds on Poultry Production
2.1. Broiler Health
2.1.1. Green Seaweeds
2.1.2. Brown Seaweeds
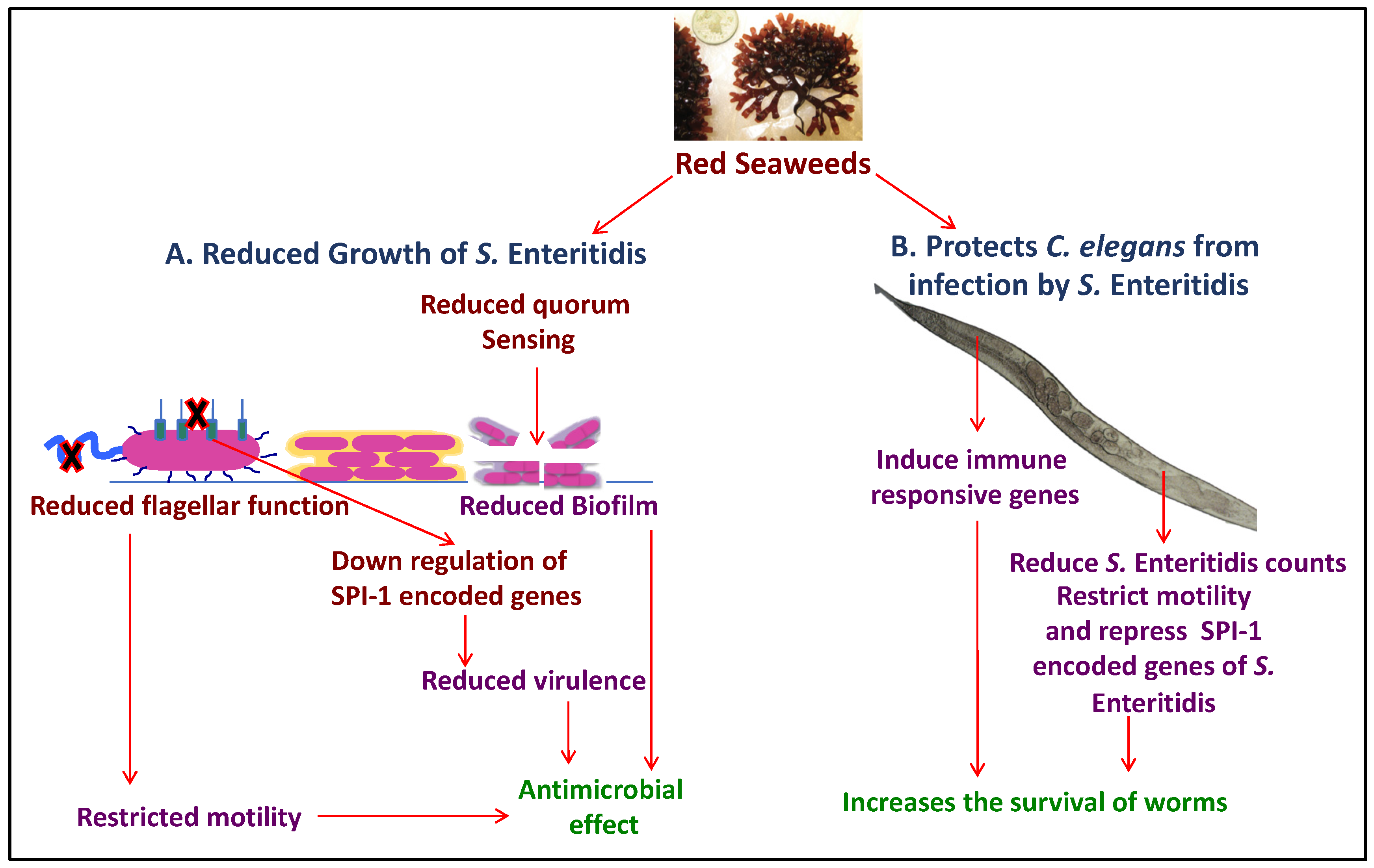
2.1.3. Red Seaweeds
2.2. Health of Laying Chickens
2.2.1. Green Seaweeds
2.2.2. Brown Seaweeds
2.2.3. Red Seaweeds
3. Novel Formulations of Seaweeds for Poultry Health
3.1. Mechanical Approach
3.1.1. Size Reduction
3.1.2. Extrusion
3.2. Additive/Synergistic Approaches
3.2.1. Biological Treatment by Fermentation
3.2.2. Botanical Blends
3.2.3. Algal Clay
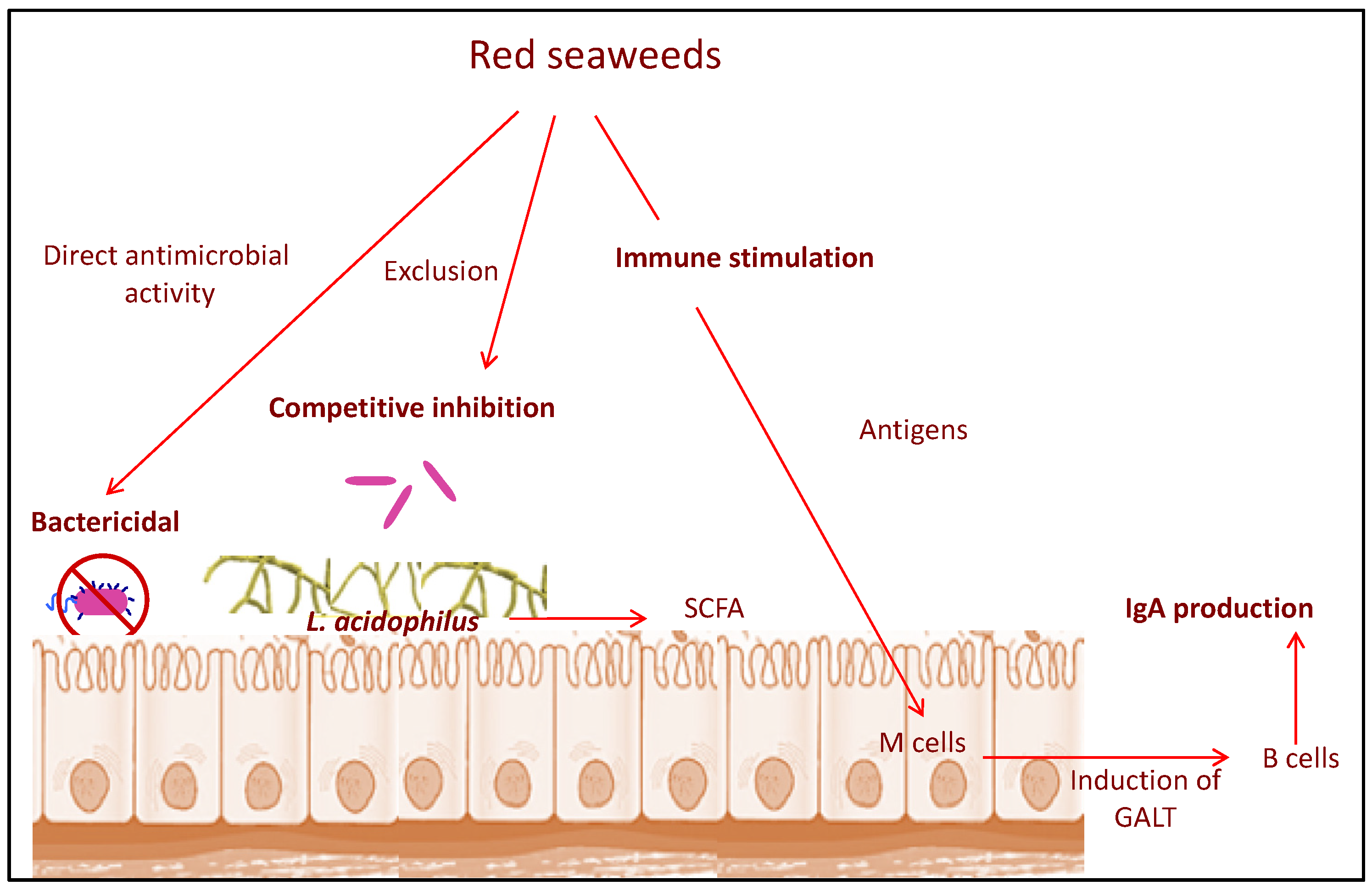
4. Anti-Bacterial and Anti-Viral Effects of Various Seaweeds on Disease in Poultry Production
4.1. Mechanism of Anti-Microbial Activity of Seaweeds
4.1.1. Anti-Bacterial Mode of Action
4.1.2. Anti-Viral Modes of Action
5. Use of Prebiotics in Poultry Production
- (1)
- They should resist digestion by acid and enzymatic hydrolysis in the upper gastrointestinal tract (GIT).
- (2)
- They must have a selective function as a substrate for the growth of beneficial bacteria.
- (3)
- They must be capable of altering the profile of the microflora.
- (4)
- They must induce beneficial effects that boost the host immune system and overall health.
6. Effect of Selected Seaweeds on the Gut Microbiome
7. Reduced Use of Antibiotics in Combination with Dietary Seaweeds
8. Commercialization of Various Seaweeds for Animal Feeds
Challenges and Future Prospects
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guiry, M.D.; Guiry, G.M.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 6 July 2020).
- Piconi, P. Edible Seaweed Market Analysis. Available online: http://www.islandinstitute.org/edible-seaweed-market-analysis-2020 (accessed on 25 January 2020).
- Lorbeer, A.J.; Tham, R.; Zhang, W. Potential products from the highly diverse and endemic macroalgae of Southern Australia and pathways for their sustainable production. J. Appl. Phycol. 2013, 25, 717–732. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Recent developments in the application of seaweeds or seaweed extracts as a means for enhancing the safety and quality attributes of foods. Innov. Food Sci. Emerg. Technol. 2011, 12, 600–609. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018–Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Comtex. Animal Feed Supplements Market Demand Status with Industry Growth 2020 Latest Trends, Top Manufacturers, Analysis by Market Size and Global Share and Forecast to 2026. Available online: https://www.marketwatch.com/press-release/animal-feed-supplements-market-demand-status-with-industry-growth-2020-latest-trends-top-manufacturers-analysis-by-market-size-and-global-share-and-forecast-to-2026-2020-05-15 (accessed on 25 May 2020).
- Allied Market Research. Animal Feed Additives Market by Additive Type (Amino Acids, Antioxidants, Feed Enzymes, Feed Acidifiers, Vitamins, Minerals, Binders, Antibiotics, and Others), Livestock (Swine, Ruminants, Poultry, Aquatic Animals, and Others), Form (Dry, Liquid, and Others), and Function (Single Function and Multifunction): Global Opportunity Analysis and Industry Forecast, 2018–2025. Available online: https://www.alliedmarketresearch.com/animal-feed-additives-market (accessed on 10 September 2019).
- Evans, F.D.; Critchley, A.T. Seaweeds for animal production use. J. Appl. Phycol. 2014, 26, 891–899. [Google Scholar] [CrossRef]
- Mišurcová, L. Chemical composition of seaweeds. In Handbook of Marine Macroalgae: Biotechnology and Applied Phycology; John Wiley & Sons: West Sussex, UK, 2011; pp. 173–192. [Google Scholar]
- Peng, Y.; Hu, J.; Yang, B.; Lin, X.P.; Zhou, X.F.; Yang, X.W.; Liu, Y. Chemical composition of seaweeds. In Seaweed Sustainability: Food and Non-Food Applications; Academic Press: Cambridge, MA, USA, 2015; pp. 79–124. [Google Scholar]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Amerah, A.M.; Ravindran, V.; Lentle, R.G.; Thomas, D.G. Feed particle size: Implications on the digestion and performance of poultry. Worlds Poult. Sci. J. 2007, 63, 439–455. [Google Scholar] [CrossRef]
- El-Deek, A.A.; Brikaa, M.A. Nutritional and biological evaluation of marine seaweed as a feedstuff and as a pellet binder in poultry diet. Int. J. Poult. Sci. 2009, 8, 875–881. [Google Scholar] [CrossRef]
- Abudabos, A.M.; Okab, A.B.; Aljumaah, R.S.; Samara, E.M.; Abdoun, K.A.; Al-Haidary, A.A. Nutritional value of green seaweed (Ulva lactuca) for broiler chickens. Ital. J. Anim. Sci. 2013, 12, e28. [Google Scholar] [CrossRef]
- Marković, R.; Šefer, D.; Krstić, M.; Petrujkić, B. Effect of different growth promoters on broiler performance and gut morphology. Arch. Med. Vet. 2009, 41, 163–169. [Google Scholar] [CrossRef]
- Cañedo-Castro, B.; Piñón-Gimate, A.; Carrillo, S.; Ramos, D.; Casas-Valdez, M. Prebiotic effect of Ulva rigida meal on the intestinal integrity and serum cholesterol and triglyceride content in broilers. J. Appl. Phycol. 2019, 1–9. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, S.R.; Oh, J.W. Effects of dietary fermented seaweed and seaweed fusiforme on growth performance, carcass parameters and immunoglobulin concentration in broiler chicks. Asian-Australas J. Anim. Sci. 2014, 27, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Gumus, R.; Urcar Gelen, S.; Koseoglu, S.; Ozkanlar, S.; Ceylan, Z.G.; Imik, H. The effects of fucoxanthin dietary inclusion on the growth performance, antioxidant metabolism and meat quality of broilers. Rev. Bras. Cienc. Avic. 2018, 20, 487–496. [Google Scholar] [CrossRef]
- Surai, P.F. Selenium in poultry nutrition 1. Antioxidant properties, deficiency and toxicity. Worlds Poult. Sci. J. 2002, 58, 333–347. [Google Scholar] [CrossRef]
- Sobotik, E.; Nelson, J.; Archer, G. The effect of feeding a seaweed extract during heat stress on broiler production and stress. In Proceedings of the Poultry Science Association Annual Meeting, San Antonio, TX, USA, 23–26 July 2018. [Google Scholar]
- El-Deek, A.A.; Al-Harthi, M.A.; Abdalla, A.A.; Elbanoby, M.M. The use of brown algae meal in finisher broiler diets. Egypt. Poult. Sci. 2011, 3, 767–781. [Google Scholar]
- Kumar, A.K. Effect of Sargassum wightii on growth, carcass and serum qualities of broiler chickens. Vet. Sci. Res. 2018, 3, 156. [Google Scholar]
- Wiseman, M. Evaluation of Tasco® as A Candidate Prebiotic in Broiler Chickens. Master’s Thesis, Dalhousie University, Halifax, NS, Canada, 2012. [Google Scholar]
- Walk, C.L.; Addo-Chidie, E.K.; Bedford, M.R.; Adeola, O. Evaluation of a highly soluble calcium source and phytase in the diets of broiler chickens. Poult. Sci. 2012, 91, 2255–2263. [Google Scholar] [CrossRef]
- Bradbury, E.J.; Wilkinson, S.J.; Cronin, G.M.; Walk, C.L.; Cowieson, A.J. The effect of marine calcium source on broiler leg integrity. In Proceedings of the 23rd Annual Australian Poultry Science Symposium, Sydney, Australia, 19–22 February 2012; pp. 85–88. [Google Scholar]
- Bradbury, E.J.; Wilkinson, S.J.; Cronin, G.M.; Thomson, P.; Walk, C.L.; Cowieson, A.J. Evaluation of the effect of a highly soluble calcium source in broiler diets supplemented with phytase on performance, nutrient digestibility, foot ash, mobility and leg weakness. Anim. Prod. Sci. 2017, 57, 2016. [Google Scholar] [CrossRef]
- Karimi, S.H. Effects of Red Seaweed (Palmaria palmata) Supplemented Diets Fed to Broiler Chickens Raised under Normal or Stressed Conditions. Master’s Thesis, Dalhousie University, Halifax, NS, Canada, 2015. [Google Scholar]
- Qadri, S.S.N.; Biswas, A.; Mandal, A.B.; Kumawat, M.; Saxena, R.; Nasir, A.M. Production performance, immune response and carcass traits of broiler chickens fed diet incorporated with Kappaphycus alvarezii. J. Appl. Phycol. 2019, 31, 753–760. [Google Scholar] [CrossRef]
- Balasubramanian, B.; Koo, J.S.; Deun, S.K.; Park, J.H.; Recharla, N.; Park, S.; Kim, I.H. Influence of marine red seaweed supplementation on growth performance, blood metabolites, breast muscle meat quality, fecal consistency score, excreta microbial shedding and noxious gas emission in broilers. In Proceedings of the Poultry Science Association Annual Meeting, San Antonio, TX, USA, 23–26 July 2019. [Google Scholar]
- Bach, S.J.; Wang, Y.; McAllister, T.A. Effect of feeding sun-dried seaweed (Ascophyllum nodosum) on fecal shedding of Escherichia coli O157:H7 by feedlot cattle and on growth performance of lambs. Anim. Feed Sci. Technol. 2008, 142, 17–32. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Dobrzański, Z.; Górecki, H.; Zielińska, A.; Korczyński, M.; Opaliński, S. Effect of macroalgae enriched with microelements on egg quality parameters and mineral content of eggs, eggshell, blood, feathers and droppings. J. Anim. Physiol. Anim. Nutr. 2011, 95, 374–387. [Google Scholar] [CrossRef]
- Wang, S.B.; Jia, Y.H.; Wang, L.H.; Zhu, F.H.; Lin, Y.T. Enteromorpha prolifera supplemental level: Effects on laying performance, egg quality, immune function and microflora in feces of laying hens. Chin. J. Anim. Nutr. 2013, 25, 1346–1352. [Google Scholar]
- Li, Q.; Luo, J.; Wang, C.; Tai, W.; Wang, H.; Zhang, X.; Liu, K.; Jia, Y.; Lyv, X.; Wang, L.; et al. Ulvan extracted from green seaweeds as new natural additives in diets for laying hens. J. Appl. Phycol. 2018, 30, 2017–2027. [Google Scholar] [CrossRef]
- Carrillo, S.; López, E.; Casas, M.M.; Avila, E.; Castillo, R.M.; Carranco, M.E.; Calvo, C.; Pérez-Gil, F. Potential use of seaweeds in the laying hen ration to improve the quality of n-3 fatty acid enriched eggs. J. Appl. Phycol. 2008, 20, 721–728. [Google Scholar] [CrossRef]
- Al-Harthi, M.A.; El-Deek, A.A. Effect of different dietary concentrations of brown marine algae (Sargassum dentifebium) prepared by different methods on plasma and yolk lipid profiles, yolk total carotene and lutein plus zeaxanthin of laying hens. Ital. J. Anim. Sci. 2012, 11, e64. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, E.C.; Na, Y.; Lee, S.R. Effects of dietary supplementation with fermented and non-fermented brown algae by-products on laying performance, egg quality, and blood profile in laying hens. Asian-Australas. J. Anim. Sci. 2018, 31, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Stupart, C. Supplementation of Red Seaweed (Chondrus crispus) and Tasco®(Ascophyllum nodosum) in Laying Hen Diets. Master’s Thesis, Dalhousie University, Halifax, NS, Canada, 2019. [Google Scholar]
- Kulshreshtha, G.; Rathgeber, B.; Stratton, G.; Thomas, N.; Evans, F.; Critchley, A.; Hafting, J.; Prithiviraj, B. Immunology, health, and disease: Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poult. Sci. 2014, 93. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, B.; Davood, S.S.; Mohammadi-Sangcheshmeh, A. Dietary supplementation of Gracilariopsis persica is associated with some quality related sera and egg yolk parameters in laying quails. J. Sci. Food Agric. 2015, 95, 643–648. [Google Scholar] [CrossRef]
- Mandal, A.B.; Biswas, A.; Mir, N.A.; Tyagi, P.K.; Kapil, D.; Biswas, A.K. Effects of dietary supplementation of Kappaphycus alvarezii on productive performance and egg quality traits of laying hens. J. Appl. Phycol. 2019, 31, 2065–2072. [Google Scholar] [CrossRef]
- Kiarie, E.G.; Mills, A. Role of feed processing on gut health and function in pigs and poultry: Conundrum of optimal particle size and hydrothermal regimens. Front. Vet. Sci. 2019, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Drake, K.; Swick, R.A.; Perez-Maldonado, R.A.; Ruhnke, I. Feed particle selection and nutrient intake altered by pecking stone consumption and beak length in free-range laying hens. Anim. Nutr. 2019, 5, 140–147. [Google Scholar] [CrossRef]
- Jacob, J.; Pescatore, T. Avian Digestive System. Available online: http://www2.ca.uky.edu/agcomm/pubs/ASC/ASC203/ASC203.pdf (accessed on 6 November 2019).
- Missotten, J.A.; Michiels, J.; Dierick, N.; Ovyn, A.; Akbarian, A.; De Smet, S. Effect of fermented moist feed on performance, gut bacteria and gut histo-morphology in broilers. Br. Poult. Sci. 2013, 54, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.J.; Huang, L.X.; Zhang, C.H.; Zhang, Y.L. Nutrient assessment of olive leaf residues processed by solid-state fermentation as an innovative feedstuff additive. J. Appl. Microbiol. 2016, 121, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.T.V.; Lu, W.J.; Tsai, G.J.; Chou, C.T.; Hsiao, H.I.; Hwang, P.A. Enhanced anti-inflammatory activity of brown seaweed Laminaria japonica by fermentation using Bacillus subtilis. Process Biochem. 2016, 51, 1945–1953. [Google Scholar] [CrossRef]
- Canibe, N.; Jensen, B.B. Fermented liquid feed-Microbial and nutritional aspects and impact on enteric diseases in pigs. Anim. Feed Sci. Technol. 2012, 173, 17–40. [Google Scholar] [CrossRef]
- Carrillo, S.; Ríos, V.H.; Calvo, C.; Carranco, M.E.; Casas, M.; Pérez-Gil, F. N-3 Fatty acid content in eggs laid by hens fed with marine algae and sardine oil and stored at different times and temperatures. J. Appl. Phycol. 2012, 24, 593–599. [Google Scholar] [CrossRef]
- Rizk, Y.S.; Ismail, I.I.; Hafsa, S.H.A.; Eshera, A.A.; Tawfeek, F.A. Effect of dietary green tea and dried seaweed on productive and physiological performance of laying hens during late phase of production. Egypt. Poult. Sci. J. 2017, 37, 685–706. [Google Scholar] [CrossRef]
- Bai, J.; Wang, R.; Yan, L.; Feng, J. Co-supplementation of dietary seaweed powder and antibacterial peptides improves broiler growth performance and immune function. Braz. J. Poult. Sci. 2019, 21. [Google Scholar] [CrossRef]
- Brennan, K.M.; Araujo, L.; Encina, C.; Salvá, B.; Flores, R. Effect of an algae-based antioxidant supplement containing selenium yeast on chicken meat quality. In Proceedings of the Poultry Science Association Annual Meeting, San Antonio, TX, USA, 23–26 July 2013. [Google Scholar]
- Bortoluzzi, C.; Schmidt, J.M.; Bordignon, H.L.F.; Fülber, L.M.; Layter, J.R.; Fernandes, J.I.M. Efficacy of yeast derived glucomannan or algae-based antioxidant or both as feed additives to ameliorate mycotoxicosis in heat stressed and unstressed broiler chickens. Livest. Sci. 2016, 193, 20–25. [Google Scholar] [CrossRef]
- Habold, C.; Reichardt, F.; Le Maho, Y.; Angel, F.; Liewig, N.; Lignot, J.H.; Oudart, H. Clay ingestion enhances intestinal triacylglycerol hydrolysis and non-esterified fatty acid absorption. Br. J. Nutr. 2009, 102, 249–257. [Google Scholar] [CrossRef]
- Frobose, H.L.; Erceg, J.A.; Fowler, S.Q.; Tokach, M.D.; DeRouchey, J.M.; Woodworth, J.C.; Dritz, S.S.; Goodband, R.D. The progression of deoxynivalenol-induced growth suppression in nursery pigs and the potential of an algae-modified montmorillonite clay to mitigate these effects. J. Anim. Sci. 2016, 94, 3746–3759. [Google Scholar] [CrossRef]
- Suarez, M.G.; Gallissot, M.; Cierpinski, P. Effect of an algae-clay mix on the use by broiler chickens of a diet containing corn DDGS. In Proceedings of the 4th International Poultry Meat Congress, Antalya, Turkey, 26–30 April 2017; pp. 321–325. [Google Scholar]
- Fumonisins, J.; Laurain, M.; Gallissot, M.; Tavares, M.N. Combating the most occurring mycotoxin in South America. In Proceedings of the Poultry Science Association Latin American Annual Meeting, Sao Paulo, Brazil, 6–8 November 2018. [Google Scholar]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Safety and efficacy of a preparation of algae interspaced bentonite as a feed additive for all animal species. EFSA J. 2016, 14. [Google Scholar] [CrossRef]
- Ventura, M.R.; Castañon, J.I.R.; McNab, J.M. Nutritional value of seaweed (Ulva rigida) for poultry. Anim. Feed Sci. Technol. 1994, 49, 87–92. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; Mcsorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs. 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed]
- Gudiel-Urbano, M.; Goñi, I. Effect of edible seaweeds (Undaria pinnatifida and Porphyra ternera) on the metabolic activities of intestinal microflora in rats. Nutr. Res. 2002, 22, 323–331. [Google Scholar] [CrossRef]
- Khosravi, M.; Gharibi, D.; Kaviani, F.; Mohammadidust, M. The antibacterial and immunomodulatory effects of carbohydrate fractions of the seaweed gracilaria persica. J. Med. Microbiol. Infect. Dis. 2018, 6, 57–61. [Google Scholar] [CrossRef]
- Ford, L.; Stratakos, A.C.; Theodoridou, K.; Dick, J.T.; Sheldrake, G.N.; Linton, M.; Corcionivoschi, N.; Walsh, P.J. Polyphenols from brown seaweeds as a potential antimicrobial agent in animal feeds. ACS Omega 2020, 5, 9093–9103. [Google Scholar] [CrossRef]
- Alghazeer, R.; Whida, F.; Abduelrhman, E.; Gammoudi, F.; Azwai, S. Screening of antibacterial activity in marine green, red and brown macroalgae from the western coast of Libya. Nat. Sci. 2013, 5, 7–14. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, P.; Yu, G.L.; Li, C.X.; Hao, C.; Qi, X.; Zhang, L.J.; Guan, H.S. Preparation and anti-influenza A virus activity of κ-carrageenan oligosaccharide and its sulphated derivatives. Food Chem. 2012, 133, 880–888. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Scolaro, L.A.; Noseda, M.D.; Cerezo, A.S.; Damonte, E.B. Protective effect of a natural carrageenan on genital herpes simplex virus infection in mice. Antivir. Res. 2004, 64, 137–141. [Google Scholar] [CrossRef]
- Talarico, L.B.; Damonte, E.B. Interference in dengue virus adsorption and uncoating by carrageenans. Virology 2007, 363, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Damonte, E.; Matulewicz, M.; Cerezo, A. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2012, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Trejo-Avila, L.M.; Morales-Martínez, M.E.; Ricque-Marie, D.; Cruz-Suarez, L.E.; Zapata-Benavides, P.; Morán-Santibañez, K.; Rodríguez-Padilla, C. In vitro anti-canine distemper virus activity of fucoidan extracted from the brown alga Cladosiphon okamuranus. VirusDisease 2014, 25, 474–480. [Google Scholar] [CrossRef]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Critchley, A.T. The COVID 19 novel coronavirus pandemic 2020: Seaweeds to the rescue? Why does substantial, supporting research about the antiviral properties of seaweed polysaccharides seem to go unrecognized by the pharmaceutical community in these desperate times? J. Appl. Phycol. 2020, 1–3. [Google Scholar] [CrossRef]
- Leonel pereire antiviral activity of seaweeds and their extracts. In Therapeutic and Nutritional Uses of Algae; CRC Press: Boca Raton, FL, USA, 2018; pp. 175–211.
- Park, H.J.; Kurokawa, M.; Shiraki, K.; Nakamura, N.; Choi, J.S.; Hattori, M. Antiviral activity of the marine alga Symphyocladia latiuscula against herpes simplex virus (HSV-1) in Vitro and its therapeutic efficacy against HSV-1 infection in mice. Biol. Pharm. Bull. 2005, 28, 2258–2262. [Google Scholar] [CrossRef]
- Diogo, J.V.; Novo, S.G.; González, M.J.; Ciancia, M.; Bratanich, A.C. Antiviral activity of lambda-carrageenan prepared from red seaweed (Gigartina skottsbergii) against BoHV-1 and SuHV-1. Res. Vet. Sci. 2015, 98, 142–144. [Google Scholar] [CrossRef]
- Sweeney, T.; Meredith, H.; Ryan, M.T.; Gath, V.; Thornton, K.; O’Doherty, J.V. Effects of Ascophyllum nodosum supplementation on Campylobacter jejuni colonisation, performance and gut health following an experimental challenge in 10 day old chicks. Innov. Food Sci. Emerg. Technol. 2016, 37, 247–252. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Rathgeber, B.; MacIsaac, J.; Boulianne, M.; Brigitte, L.; Stratton, G.; Thomas, N.A.; Critchley, A.T.; Hafting, J.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, reduce Salmonella Enteritidis in laying hens. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and comparison of the structural features, immune-modulatory and anti-avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs. 2016, 14, 4. [Google Scholar] [CrossRef]
- Aguilar-Briseño, J.A.; Cruz-Suarez, L.E.; Sassi, J.F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated polysaccharides from Ulva clathrata and Cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion, in NDV infection. Mar. Drugs. 2015, 13, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Paiva, D.M.; Walk, C.L.; McElroy, A.P. Influence of dietary calcium level, calcium source, and phytase on bird performance and mineral digestibility during a natural necrotic enteritis episode. Poult. Sci. 2013, 92, 3125–3133. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, S.; Ramanathan, G.; Jacob Inbaneson, S.; Ramu, A. Antiplasmodial activity of two marine polyherbal preparations from Chaetomorpha antennina and Aegiceras corniculatum against Plasmodium falciparum. Parasitol. Res. 2011, 108, 107–113. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D.; Jahnke, L.S. Seasonal effects of sun exposure and emersion on intertidal seaweed physiology: Fluctuations in antioxidant contents, photosynthetic pigments and photosynthetic efficiency in the red alga Porphyra umbilicalis Kützing (Rhodophyta, Bangiales). J. Exp. Mar. Bio. Ecol. 2008, 361, 83–91. [Google Scholar] [CrossRef]
- O’Connor, D.O.; Rubino, J.R. Phenolic compounds. In Disinfection, Sterilization and Preservation; Block, S.S., Ed.; Lea and Febiger: Philadelphia, PA, USA, 1991; pp. 204–224. [Google Scholar]
- Potin, P.; Bouarab, K.; Küpper, F.; Kloareg, B. Oligosaccharide recognition signals and defence reactions in marine plant-microbe interactions. Curr. Opin. Microbiol. 1999, 2, 276–283. [Google Scholar] [CrossRef]
- Choi, J.; Shin, D.; Kim, M.; Park, J.; Lim, S.; Ryu, S. LsrR-mediated quorum sensing controls invasiveness of salmonella typhimurium by regulating SPI-1 and flagella genes. PLoS ONE 2012, 7, e37059. [Google Scholar] [CrossRef] [PubMed]
- Jesudhasan, P.R.; Cepeda, M.L.; Widmer, K.; Dowd, S.E.; Soni, K.A.; Hume, M.E.; Zhu, J.; Pillai, S.D. Transcriptome analysis of genes controlled by luxS/Autoinducer-2 in salmonella enterica serovar typhimurium. Foodborne Pathog. Dis. 2010, 7, 399–410. [Google Scholar] [CrossRef]
- Janssens, J.C.A.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated furanones inhibit biofilm formation by Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef]
- Manefield, M.; De Nys, R.; Kumar, N.; Read, R.; Givskov, M.; Steinberg, P.; Kjelleberg, S. Evidence that halogenated furanones from Delisea pulchra inhibit acylated homoserine lactone (AHL)-mediated gene expression by displacing the AHL signal from its receptor protein. Microbiology 1999, 145, 283–291. [Google Scholar] [CrossRef]
- Kulshreshtha, G. The Use of Selected Red Macroalgae (Seaweeds) for the Reduction of Salmonella Enteritidis in Poultry. Ph.D. Thesis, Dalhousie University, Halifax, NS, Canada, 2016. [Google Scholar]
- Ofek, I.; Mirelman, D.; Sharon, N. Adherence of escherichia coli to human mucosal cells mediated by mannose receptors. Nature 1977, 265, 623–625. [Google Scholar] [CrossRef]
- Karbassi, E.; Asadinezhad, A.; Lehocký, M.; Humpolíček, P.; Vesel, A.; Novák, I.; Sáha, P. Antibacterial performance of alginic acid coating on polyethylene film. Int. J. Mol. Sci. 2014, 15, 14684–14696. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Borza, T.; Rathgeber, B.; Stratton, G.S.; Thomas, N.A.; Critchley, A.; Hafting, J.; Prithiviraj, B. Red seaweeds Sarcodiotheca gaudichaudii and chondrus crispus down regulate virulence factors of salmonella enteritidis and induce immune responses in Caenorhabditis elegans. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Hirayama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-mannose specific lectin and its recombinants from a carrageenophyta kappaphycus alvarezii represent a potent anti-HIV activity through high-affinity binding to the viral envelope glycoprotein gp120. Mar. Biotechnol. 2016, 18, 215–231. [Google Scholar] [CrossRef]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs. 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wang, A.; Lu, Z.; Qin, C.; Hu, J.; Yin, J. Overview on the antiviral activities and mechanisms of marine polysaccharides from seaweeds. Carbohydr. Res. 2017, 453–454, 1–9. [Google Scholar] [CrossRef]
- Patterson, J.A.; Burkholder, K. Application of prebiotics and probiotics in poultry production. Poult. Sci. 2003, 82, 627–631. [Google Scholar] [CrossRef]
- Hajati, H.; Rezaei, M. The application of prebiotics in poultry production mitochondrial energetics in response to different fat types in heat-stressed broilers view project effects of fat sources on energetics and redox statues of liver mitochondria in heat-stressd broilers Vi. Artic. Int. J. Poult. Sci. 2010, 9, 298–304. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Ricke, S.C. Impact of prebiotics on poultry production and food safety. Yale J. Biol Med. 2018, 91, 151–159. [Google Scholar] [PubMed]
- Cummings, J.H.; Macfarlane, G.T. Gastrointestinal effects of prebiotics. Br. J. Nutr. 2002, 87, S145–S151. [Google Scholar] [CrossRef]
- Reddy, B.S. Possible mechanisms by which pro- and prebiotics influence colon carcinogenesis and tumor growth. J. Nutr. 1999, 129, 1478S–1482S. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Apajalahti, J. Comparative gut microflora, metabolic challenges, and potential opportunities. J. Appl. Poult. Res. 2005, 14, 444–453. [Google Scholar] [CrossRef]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 15, 344–354. [Google Scholar] [CrossRef]
- Liu, J.; Kandasamy, S.; Zhang, J.; Kirby, C.W.; Karakach, T.; Hafting, J.; Critchley, A.T.; Evans, F.; Prithiviraj, B. Prebiotic effects of diet supplemented with the cultivated red seaweed Chondrus crispus or with fructo-oligo-saccharide on host immunity, colonic microbiota and gut microbial metabolites. BMC Complement. Altern. Med. 2015, 15, 279. [Google Scholar] [CrossRef]
- Ramnani, P.; Chitarrari, R.; Tuohy, K.; Grant, J.; Hotchkiss, S.; Philp, K.; Campbell, R.; Gill, C.; Rowland, I. Invitro fermentation and prebiotic potential of novel low molecular weight polysaccharides derived from agar and alginate seaweeds. Anaerobe 2012, 18, 1–6. [Google Scholar] [CrossRef]
- Hu, B.; Gong, Q.; Wang, Y.; Ma, Y.; Li, J.; Yu, W. Prebiotic effects of neoagaro-oligosaccharides prepared by enzymatic hydrolysis of agarose. Anaerobe 2006, 12, 260–266. [Google Scholar] [CrossRef]
- Ai, C.; Jiang, P.; Liu, Y.; Duan, M.; Sun, X.; Luo, T.; Jiang, G.; Song, S. The specific use of alginate from: Laminaria japonica by Bacteroides species determined its modulation of the Bacteroides community. Food Funct. 2019, 10, 4304–4314. [Google Scholar] [CrossRef]
- Nakata, T.; Kyoui, D.; Takahashi, H.; Kimura, B.; Kuda, T. Inhibitory effects of laminaran and alginate on production of putrefactive compounds from soy protein by intestinal microbiota in vitro and in rats. Carbohydr. Polym. 2016, 143, 61–69. [Google Scholar] [CrossRef]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of: Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Conlon, M.A.; Methacanon, P.; Franco, C.M.M.; Su, P.; Zhang, W. Gut health benefits of brown seaweed Ecklonia radiata and its polysaccharides demonstrated in vivo in a rat model. J. Funct. Foods 2017, 37, 676–684. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; Mcmullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Antimicrobial Resistance. Available online: https://www.fda.gov/animal-veterinary/safety-health/antimicrobial-resistance (accessed on 25 May 2020).
- Maday, J. Princeton Study: Antibiotic Resistance Increasing Globally. Available online: https://www.drovers.com/article/princeton-study-antibiotic-resistance-increasing-globally (accessed on 16 November 2019).
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- And middle-income countries. Science 2019, 365. [Google Scholar] [CrossRef]
- Hussin, W.A.; El-Sayed, W.M. Synergic interactions between selected botanical extracts and tetracycline against gram positive and gram negative bacteria. J. Biol. Sci. 2011, 11, 433–441. [Google Scholar] [CrossRef]
- Chovanová, R.; Mezovská, J.; Vaverková, Š.; Mikulášová, M. The inhibition the Tet(K) efflux pump of tetracycline resistant Staphylococcus epidermidis by essential oils from three Salvia species. Lett. Appl. Microbiol. 2015, 61, 58–62. [Google Scholar] [CrossRef]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming drug resistance with alginate oligosaccharides able to potentiate the action of selected antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.-J.; Lin, H.-J.; Hsu, P.-H.; Lai, M.; Chiu, J.-Y.; Lin, H.-T.V. Brown and red seaweeds serve as potential efflux pump inhibitors for drug-resistant escherichia coli. Evid.–Based Complement. Altern. Med. 2019. [Google Scholar] [CrossRef]
- Prithiviraj, B.; Kulshreshtha, G. Use of Floridoside or Isethionic Acid to Potentiate Antimicrobial Activity of Antibiotics. Available online: https://patents.google.com/patent/US20190038651A1/en (accessed on 20 January 2020).
- Fortune Business Insight. Commercial Seaweed Market Size, Share and Global Trend by Product Type (Red, Brown, Green), by Form (Flakes, Powder, Liquid), by END user (Food and Beverage, Agriculture Fertilizer, Animal Feed Additives, Pharmaceuticals, Cosmetics, and Personal Care) and Regional Forecast Till 2025. Available online: http://web.archive.org/web/20200510003328/https://www.fortunebusinessinsights.com/industry-reports/commercial-seaweed-market-100077 (accessed on 24 September 2019).
- Grand View Research Inc. Commercial Seaweeds Market Size, Share & Trends Analysis Report By Product (Brown Seaweeds, Red Seaweeds, Green Seaweeds), By Form (Liquid, Powdered, Flakes), By Application, By Region, And Segment Forecasts, 2020–2027. Available online: https://www.grandviewresearch.com/industry-analysis/commercial-seaweed-market (accessed on 25 May 2020).
- Mac Monagail, M.; Cornish, L.; Morrison, L.; Araújo, R.; Critchley, A.T. Sustainable harvesting of wild seaweed resources. Eur. J. Phycol. 2017, 52, 371–390. [Google Scholar] [CrossRef]
- Cornish, L. The animal kingdom and seaweeds. J. Mar. Sci. Pers. Comm. 2020. in preparation. [Google Scholar]
- Lerat, Y.; Cornish, M.L.; Critchley, A.T. Applications of algal biomass in global food and feed markets: From traditional usage to the potential for functional products. In Blue Biotechnology: Production and Use of Marine Molecules; Wiley YHC Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; pp. 143–189. [Google Scholar]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Ratcliff, J.J.; Wan, A.H.L.; Edwards, M.D.; Soler-Vila, A.; Johnson, M.P.; Abreu, M.H.; Morrison, L. Metal content of kelp (Laminaria digitata) co-cultivated with Atlantic salmon in an Integrated Multi-Trophic Aquaculture system. Aquaculture 2016, 450, 234–243. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, T.; Shi, H.; Gao, K.; Huang, W.; Xu, J.; Wang, J.; Wang, R.; Li, J.; Gao, G. Microplastics in bloom-forming macroalgae: Distribution, characteristics and impacts. J. Hazard. Mater. 2020, 397, 122752. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhang, T.; Wang, J.; Huang, W.; Wang, R.; Xu, J.; Fu, G.; Gao, G. Spatio-temporal features of microplastics pollution in macroalgae growing in an important mariculture area, China. Sci. Total Environ. 2020, 719, 137490. [Google Scholar] [CrossRef]
- Mac Monagail, M.; Morrison, L. Arsenic speciation in a variety of seaweeds and associated food products. Compr. Anal. Chem. 2019, 85, 267–310. [Google Scholar]
- Rajauria, G. Seaweeds: A sustainable feed source for livestock and aquaculture. Seaweed Sustain. 2015, 389–420. [Google Scholar]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macroalgae B = Brown R = Red G = Green | Level of Inclusion in Feed | Anti-Microbial Response/Poultry Disease | Reference |
|---|---|---|---|
| Laminaria japonica (LJP) (B) and anti-microbial peptide cecropin | Laminaria japonica LJP:1%, 3% and 5%; Cecropin: 0.03% | Anti-bacterial and anti-viral activities were observed with dietary supplementation of broiler diets with LJP + cecropin, which increased feed conversion ratio (FCR), and serum Newcastle disease antibody titers and lymphocyte numbers. In addition, birds fed with LJP showed significant inhibition of E. coli counts and increase in Lactobacillus counts in ceca. | [51] |
| Ascophyllum nodosum (B) | 0.05% and 0.1% | Anti-bacterial activity. A. nodosum reduced C. jejuni counts in the caecum of chicks (10 days old), at both concentrations, but decreased the growth parameters (disruptive effect on gut morphology in ileum). Significant increases in the expression of tight-junction genes OCLN and CLND-1 alongside increases in MUC2 and CCND1 expression. | [76] |
| Chondrus crispus and Sarcodiotheca gaudichaudii (R) | 2% and 4% | Anti-bacterial activity. The incorporation of SW in the diets of Lohmann Lite laying hens reduced the negative effects of Salmonella enteritidis (SE) infection on body weight and egg production. | [77] |
| Grateloupia filicina, (R) Ulva pertusa (G) and Sargassum qingdaoense (B) | In vitro, 20–500 mg/mL of sulphated polysaccharides SPs; in vivo mouse model, 0.001% and 0.005% of SPs per day | Anti-viral activity. Sulphated polysaccharide extracts from all three species showed immune-modulatory activities, both in vitro and in vivo; S. qingdaoense showed the best activity. All three SPs significantly inhibited the activity of activated AIV (H9N2 subtype) in vitro and inactivated avian influenza virus (AIV) in vivo. Sulphated polysaccharides from G. filicina showed the strongest anti-AIV response. | [78] |
| Ulva clathrata (G) and fucoidan | In vitro 0.1–1000 μg/mL | Anti-viral activity. The ulvan and fucoidan extracts inhibited Newcastle disease virus (NDV) in vitro and showed no cytotoxicity at effective concentrations. Ulvan inhibited viral fusion by interacting with the intact F0 protein. Ulvan exhibited better anti-cell–cell spread activity than fucoidans, but a combination showed more potent (synergistic) responses. | [79] |
| Highly soluble calcified seaweed (HSC) (R) | 0.6% and 0.9% | Anti-bacterial activity. Broilers fed HSC diets had significantly higher feed conversion/total weight than control birds. Lower dietary Ca (0.6% vs. 0.9%) showed lower mortality associated with necrotic enteritis (NE) as compared to higher dose (0.9%) and on bird performance. | [80] |
| Chaetomorpha antennina (G) in combination with mangrove species Aegiceras corniculatum (land plant) | In vitro: 0.5, 1.0 and 1.5 mg/mL In vivo rat model: 0.02% per day | Parasite inhibition. The extract mixture showed 60% suppression of parasitaemia against Plasmodium falciparum at 1.5 mg/mL. Anti-plasmodial activity (50%) against Plasmodium berghei was observed in vivo. | [81] |
| Seaweed Source Red (R), Brown (B) or Green (G) | Component | Type of Study/Level of Inclusion | Microbiome Modulation | Metabolome Modulation | Other Responses | Reference |
|---|---|---|---|---|---|---|
| Palmaria palmata (R) | whole seaweed | 0.6%, 1.2%, 1.8%, 2.4% and 3%. In vitro in broiler chickens | Method: Microbiology culture techniques, 16SRNA amplicon sequencing. ↑Bifidobacterium ↑ Lactobacillus (ileum) ↓ Clostridium perfringens | Not measured | Increasing trend in the size of villus height, width, villus surface area and mucosal depth ↑ Plasma immunoglobulin (IgA and IgG) Best response: 1.8% | [28] |
| Chondrus crispus and Sarcodiotheca gaudichaudii (R) | whole seaweed | 0.5%,1% and 2% | Method: Real-time PCR. ↑Bifidobacterium longum, ↑Lactobacillus acidophilus ↑Streptococcus salivarius ↓Clostridium perfringens | ↑Acetic, ↑propionic acid, ↑n-butyric acid ↑i-butyric | Increase in the size of villus height, width, villus surface area and mucosal depth Increase in ceca weight | [39] |
| Chondrus crispus and Sarcodiotheca gaudichaudii (R) | whole seaweeds | 2% and 4% | Method: Microbiology culture techniques, 16SRNA amplicon sequencing. ↑ Firmicutes and Bacteroidetes ↑Bifidobacterium longum, ↑Lactobacillus acidophilus, ↑Streptococcus salivarius ↓ Clostridium perfringens ↓ Salmonella Enteritidis | ↑ Propionic acid | ↑ Plasma immunoglobulin (IgA and IgG) | [77] |
| Chondrus crispus (R) | whole seaweeds | 0.5 and 2.5% In vivo mouse model | Method: 16S rRNA sequencing-based Phylochip array of fecal samples. ↑ Bifidobacterium breve ↓ Clostridium septicum and Streptococcus pneumonia | ↑Acetic, ↑propionic and ↑butyric acids in faecal samples | Improvements in proximal colon histo-morphology ↑ Plasma immunoglobulin (IgA and IgG) | [105] |
| Gracilaria spp. (R), Gelidium sesquipidale (R) and Ascophyllum nodosum (B) | agar and alginate | Anaerobic fermentation | Method: Fluorescent in-situ hybridization. ↑Bifidogenic effect | ↑Acetic, propionic | Not measured | [106] |
| Red seaweed | neo-agaro-oligosaccharides (NAOS) from enzymatic hydrolysis of agarose | 2.5% and 5% In vivo mouse model | Method: Microbiology culture techniques using cecal and fecal samples. ↑Bifidobacteria and ↑Lactobacilli Reduced putrefactive microorganisms. | Not measured | No side effects, such as eructation and bloating, were observed | [107] |
| Saccharina (Laminiaria) japonica (B) | alginate | 4 mg/mL, simulated oral, gastric and small intestinal digestion | Method: 16S rRNA sequencing-based high throughput sequencing, MALDI-TOF/MS. Bacteroides (Bacteroides finegoldii) | ↑Acetic acid and ↑propionic acid | Specific modulation of Bacteroides by alginates | [108] |
| Eisenia bicyclis (B) | laminarin | 2% (w/w) In vivo in rats | Method: 16S rDNA-DGGE and Pyrosequencing. ↑ Catabacter hongkongensis ↑ Stomatobaculum longum ↓ Adlercreuzia ↓ Helicobacter | ↓ Indole | Not measured | [109] |
| Ascophyllum nodosum and Laminaria japonica | Fucoidan | 0.01% day−1 In vivo in mice | Method: 16S rRNA sequencing-based high throughput sequencing. ↑ Lactobacillus ↑ Ruminococcaceae ↓ Peptococcus | Not measured | Reduced the antigen load and the inflammatory response | [110] |
| Ecklonia radiata (B) | Whole seaweed and polysaccharide fraction (fucoidan and alginate) | 5% (w/w) WS 5% (w/w) PF in vivo in rats | Method: Real time Q-PCR. ↑F. prausnitzii ↑E. coli (PF) ↓ Enterococcus (WS) ↓Lactobacillus ↓Bifidobacterium ↓Firmicutes: Bacteroidetes | ↑ Acetate ↑ Propionate ↑ Butyrate (PF) ↓ Valerate ↓ Hexanoate ↑ Total SCFA ↓ i-Butyrate ↓ i-Valerate ↓ phenol ↓ p-cresol | ↑ Butyrate (PF) Linked to anti-inflammatory action | [111] |
| Business Organization | Product/Source/ Description | Stage of Development/ Operation Level | Product Function/ Claims | Animal Health Sector | Web Address |
|---|---|---|---|---|---|
| Ekogea, UK | -BCxF®, prebiotic poultry water additive (comprised of Ascophyllum Nodosum). -BCxS, animal housing sanitizer, 100% Ascophyllum Nodosum | Commercialized market, industrial scale | -Reduces mortality; -Improves feed conversion, weight gain and overall bird health, gut health; -Reduces ammonia emission and Campylobacter levels. | Poultry, pigs and other animals | http://www.ekogea.co.uk/ |
| Ocean Harvest Technology, Ireland | -OceanFeedTM -Ocean Poultry contains a complex blend of seaweeds (red, green and brown) based feed additive | Commercialized market, industrial scale | -Improves body weight gain; -Increases breast meat yield increased. | Poultry, pigs and canines (dogs) | https://www.oceanharvesttechnology.com |
| Olmix Group | Mycotoxin risk: -MT.X+ -MMI.S Digestive efficiency: -MFeed+ -DigestSea Immunity: -Searup -Algimun | Commercialized market, industrial scale | -Reduces mycotoxins in feed; -Immune modulation; -Improves feed efficiency. | Poultry, pigs and canines | https://www.olmix.com |
| Algea, The Arctic Company, Kristiansund - Omagata | AlgaeFeed 1.4 AlgaeFeed 3.5 Brown seaweed meal containing mineral macro and micro elements | Commercialized market, industrial scale | -Improves animal metabolism and performance; -Improves eggshell quality and production of eggs. | Poultry, pigs, cattle, fish and equines | https://www.algea.com/ |
| FutureFeed, Australia | Asparagopsis seaweed-based diet | Conducting trials for commercialization/ pilot scale | -Anti-bacterial properties; -Methane reduction in livestock digestive fermentation. | Livestock | https://www.csiro.au/en/Research/AF/Areas/Food-security/FutureFeed |
| SeaLac, Seaweed production | Ascophyllum nodosum-based seaweed supplement- organic dried seaweed-food grade | Commercialized market, industrial scale | -Improves feed absorption, weight gain in broilers; -Natural alternative to antibiotics; -Improves egg production, egg quality and immune response. | Poultry, pigs and cattle | http://www.sealac.eu/ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulshreshtha, G.; Hincke, M.T.; Prithiviraj, B.; Critchley, A. A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens. J. Mar. Sci. Eng. 2020, 8, 536. https://doi.org/10.3390/jmse8070536
Kulshreshtha G, Hincke MT, Prithiviraj B, Critchley A. A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens. Journal of Marine Science and Engineering. 2020; 8(7):536. https://doi.org/10.3390/jmse8070536
Chicago/Turabian StyleKulshreshtha, Garima, Maxwell T. Hincke, Balakrishnan Prithiviraj, and Alan Critchley. 2020. "A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens" Journal of Marine Science and Engineering 8, no. 7: 536. https://doi.org/10.3390/jmse8070536
APA StyleKulshreshtha, G., Hincke, M. T., Prithiviraj, B., & Critchley, A. (2020). A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens. Journal of Marine Science and Engineering, 8(7), 536. https://doi.org/10.3390/jmse8070536