Zooplankton Distribution and Community Structure in the Pacific and Atlantic Sectors of the Southern Ocean during Austral Summer 2017–18: A Pilot Study Conducted from Ukrainian Long-Liners


 and
and
Abstract
1. Introduction
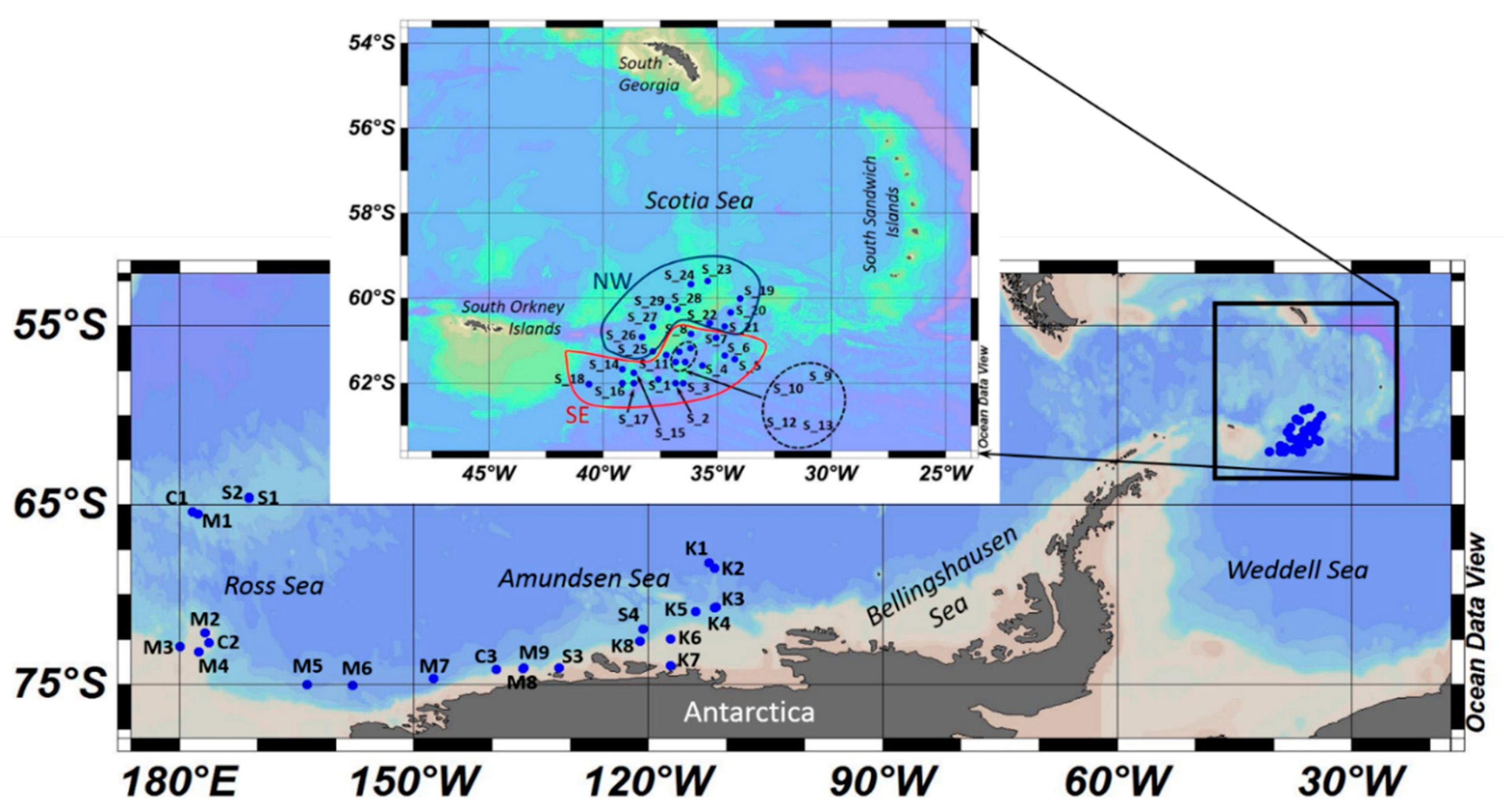
2. Materials and Methods
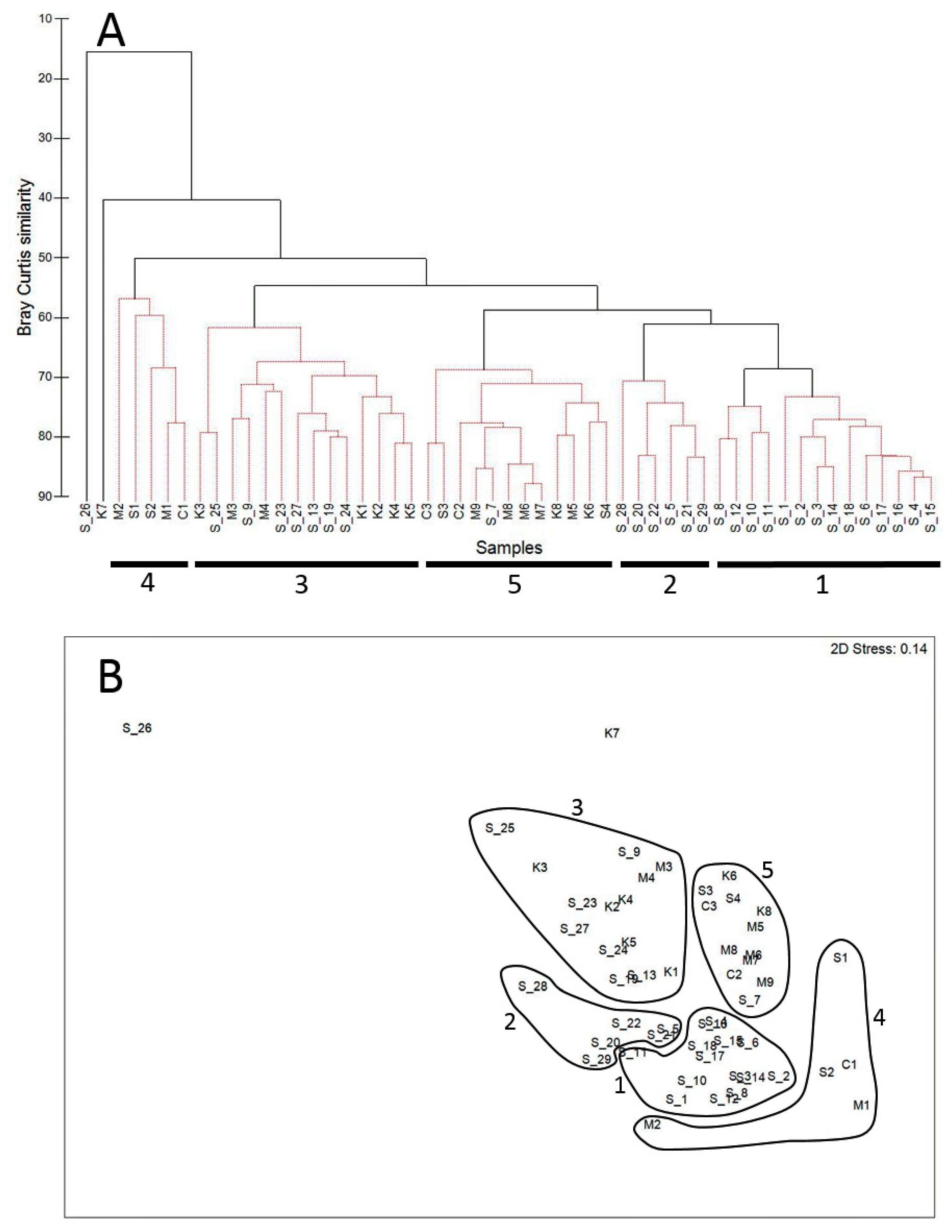
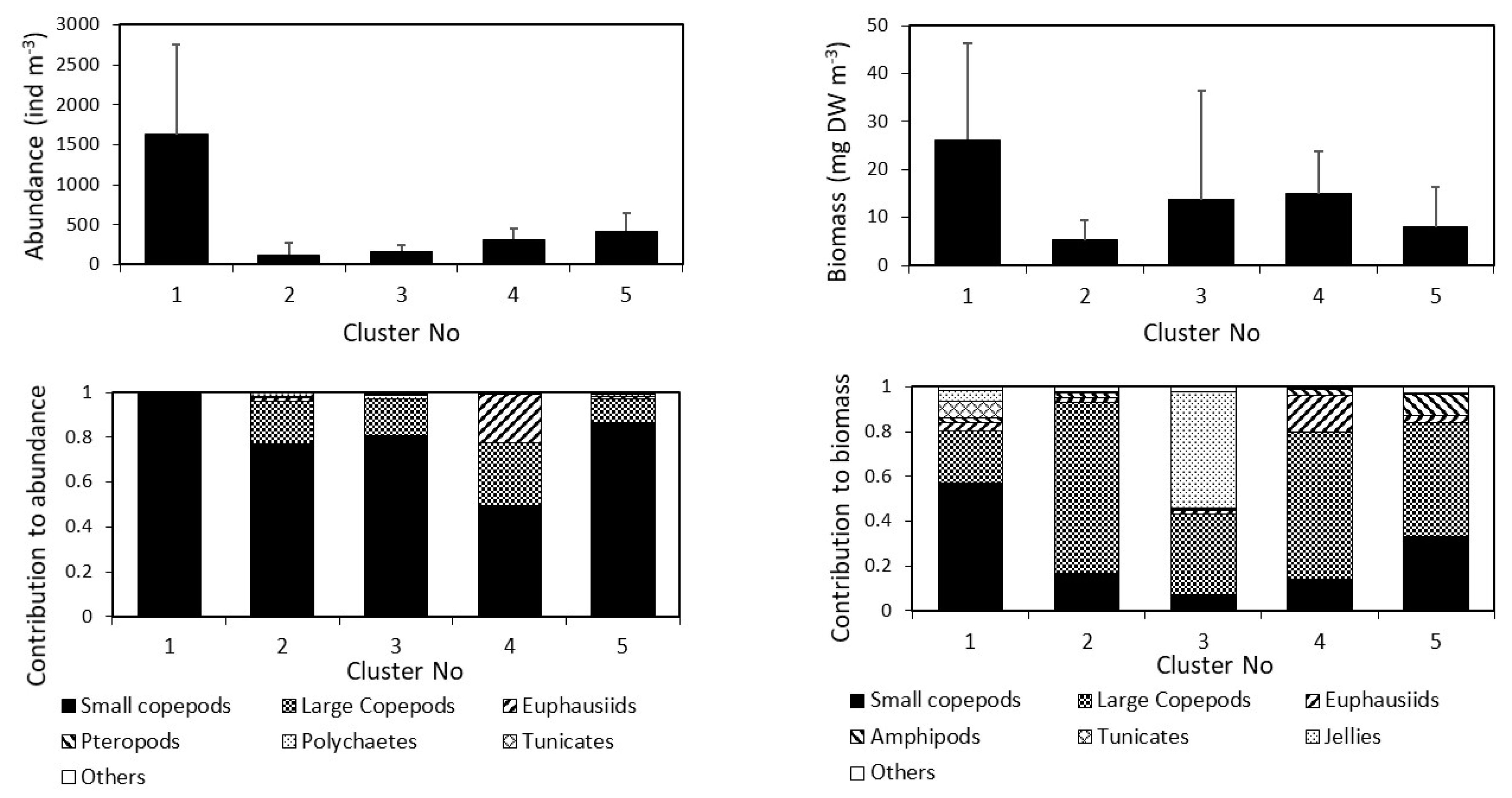
3. Results
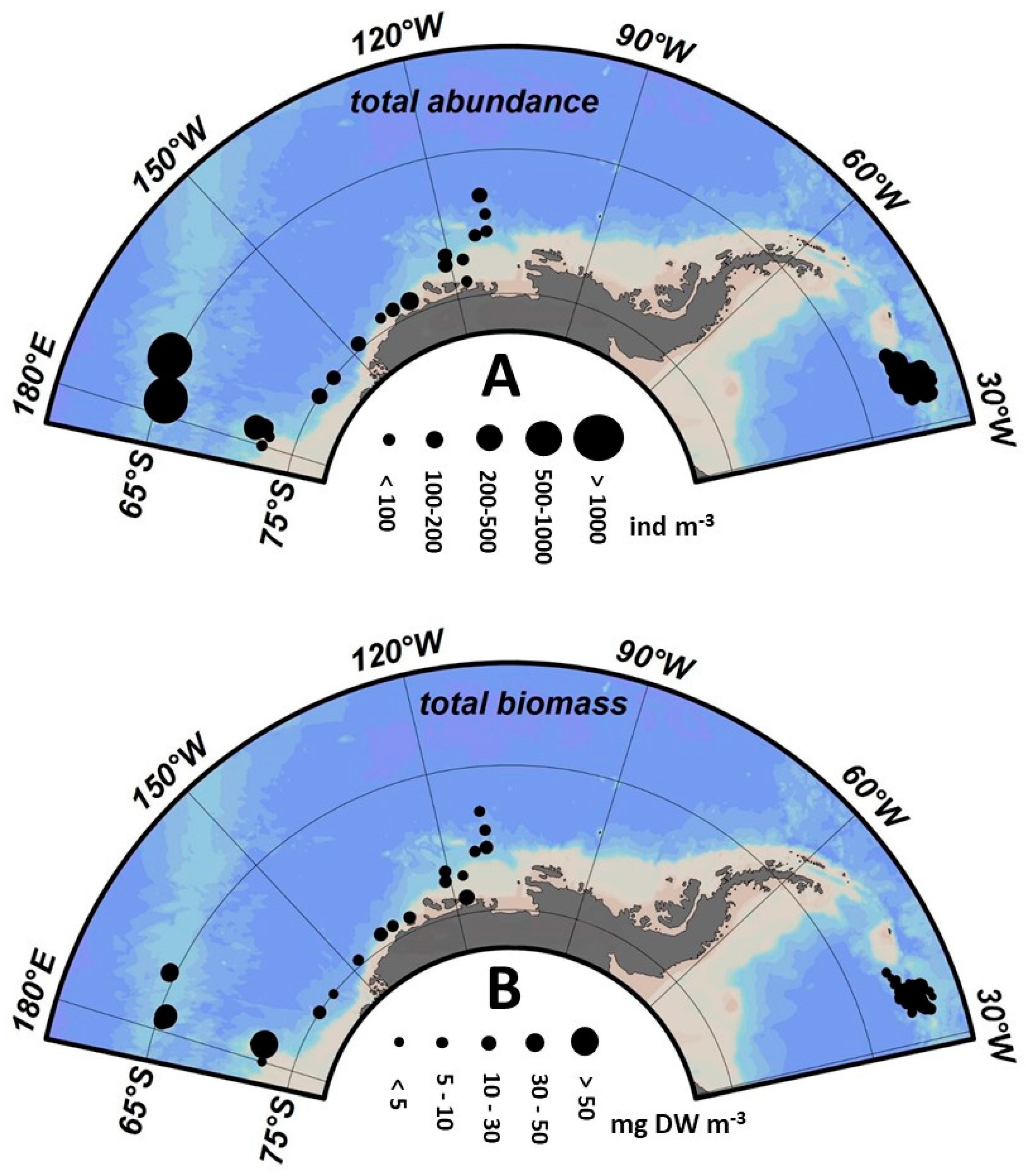
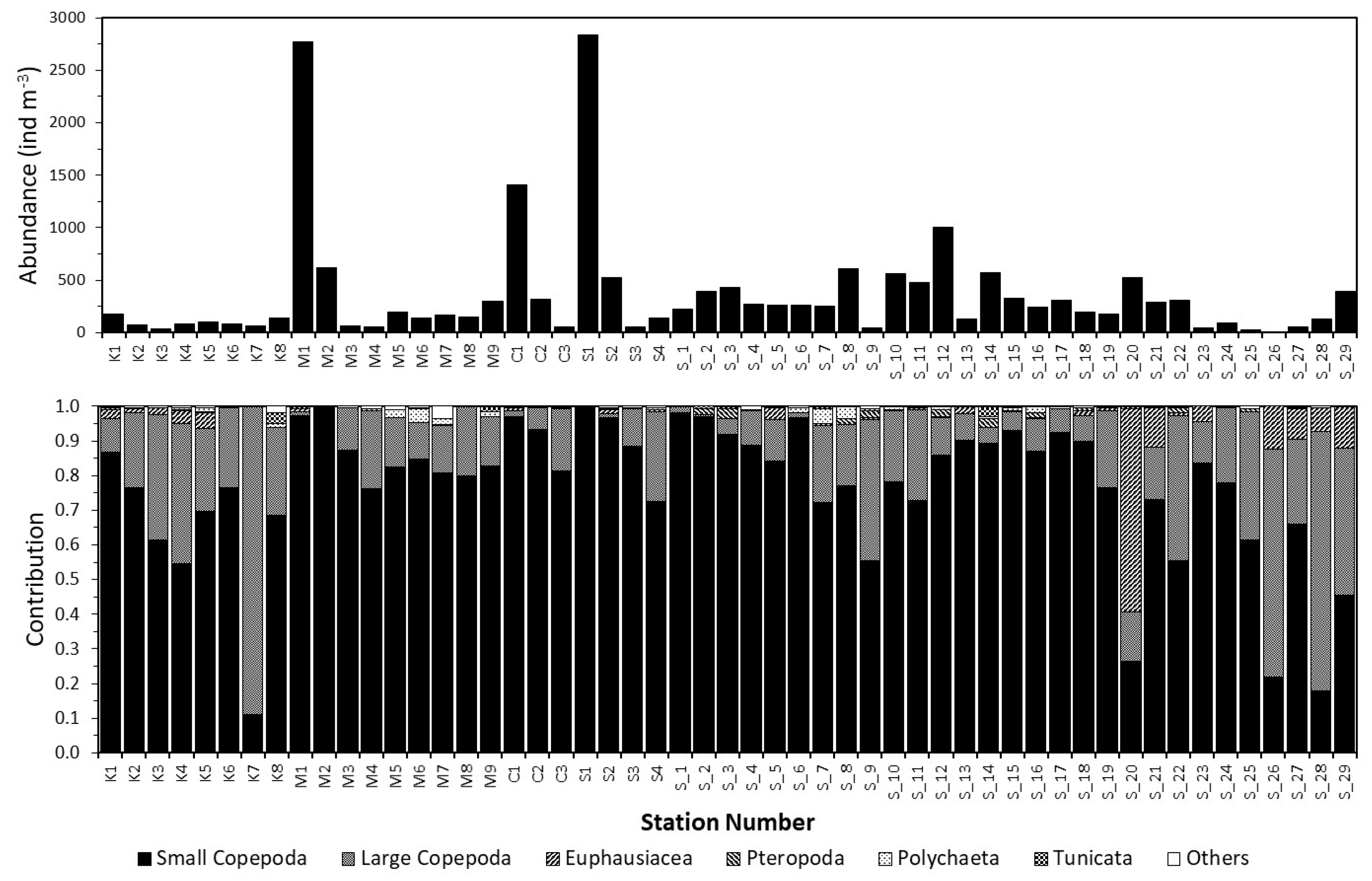
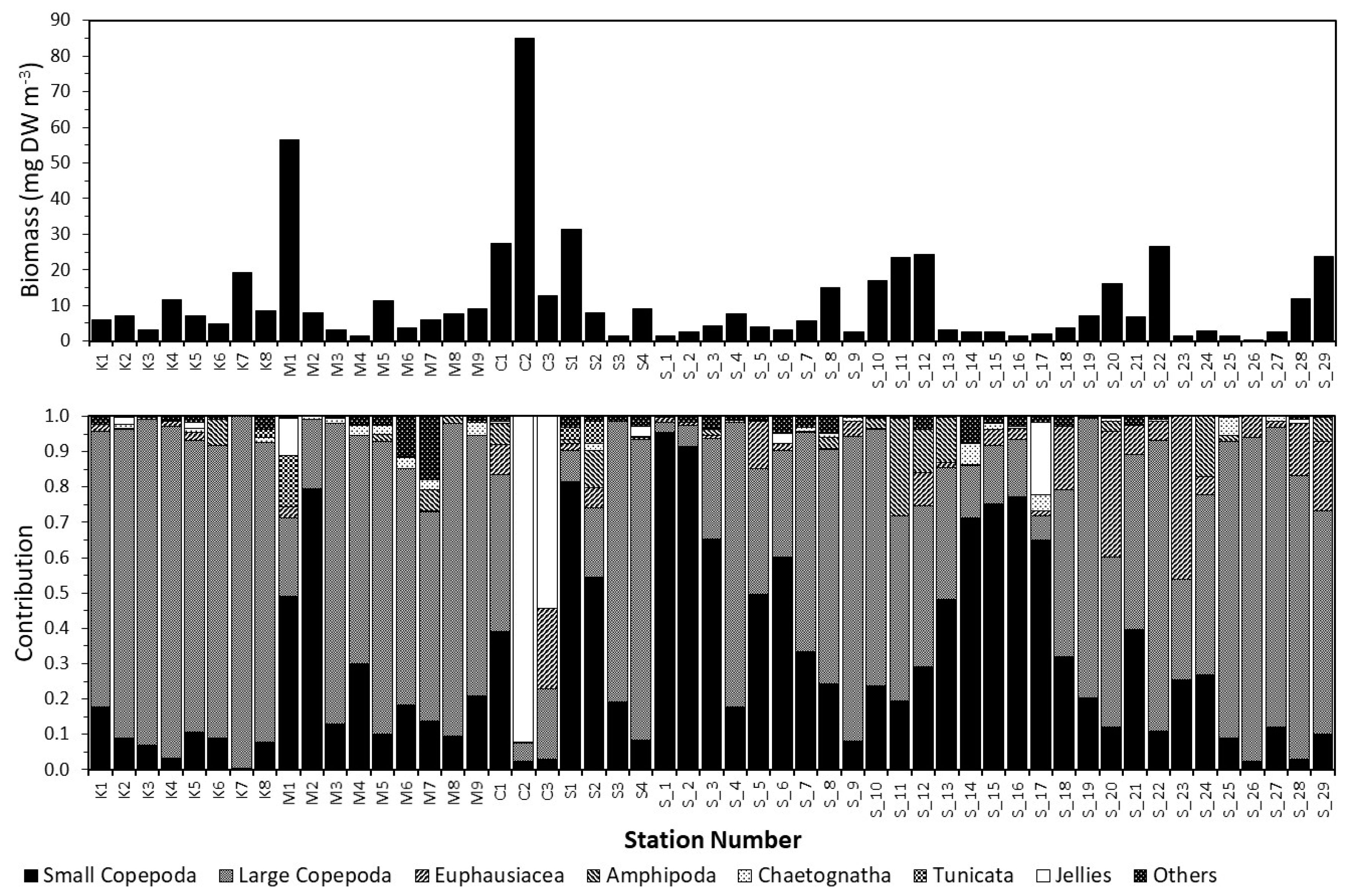
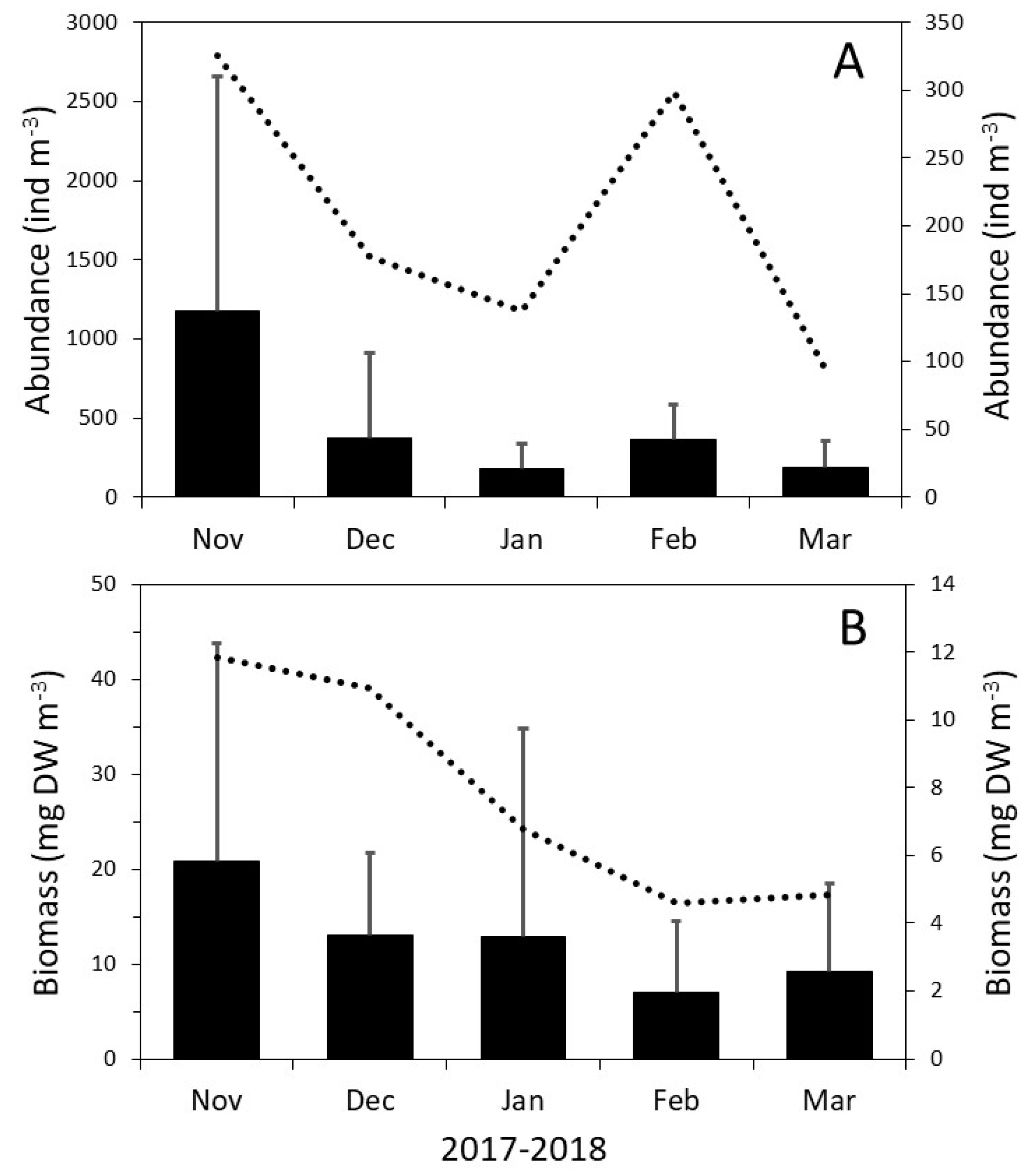
3.1. Spatial Patterns in the Zooplankton Density and Composition
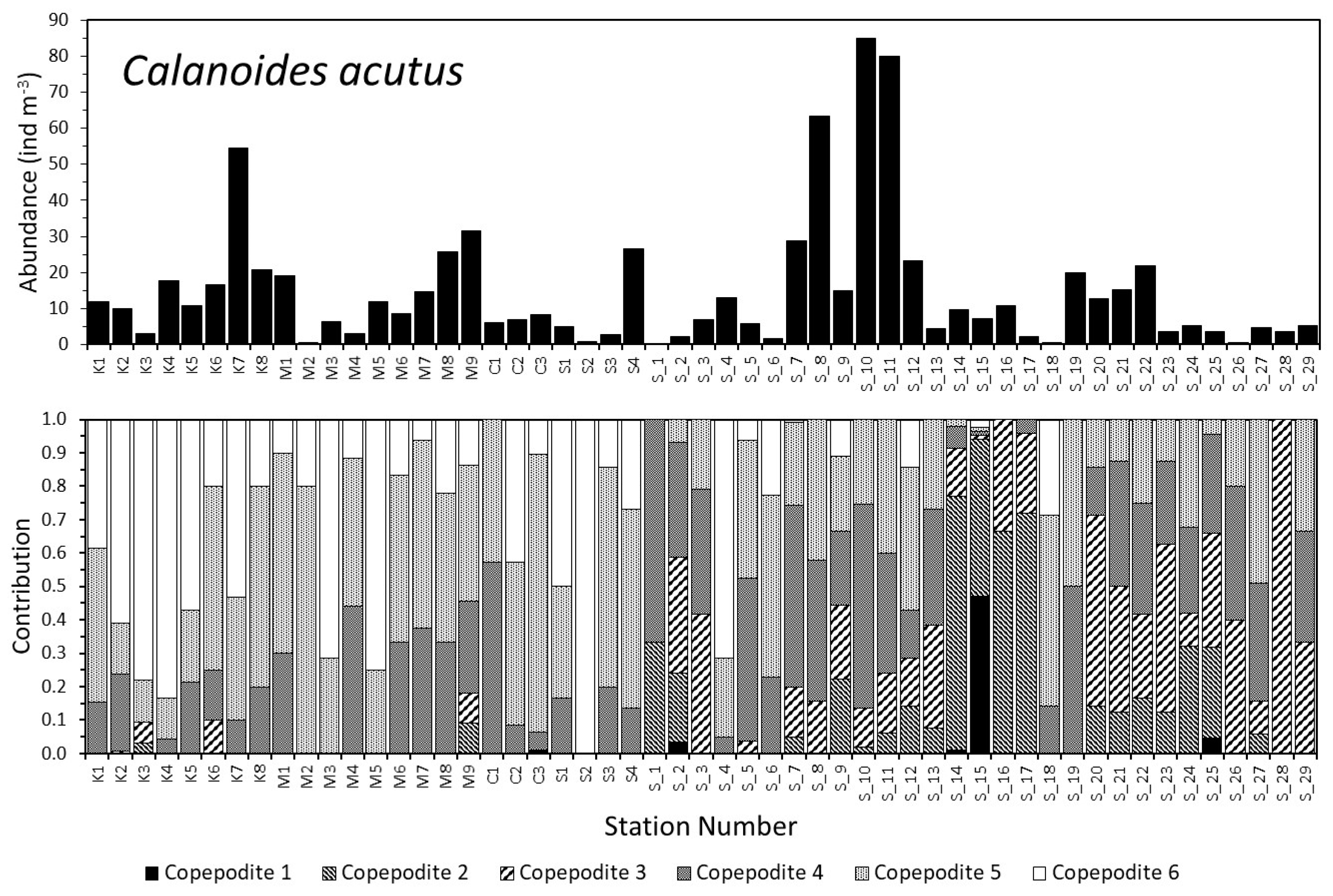
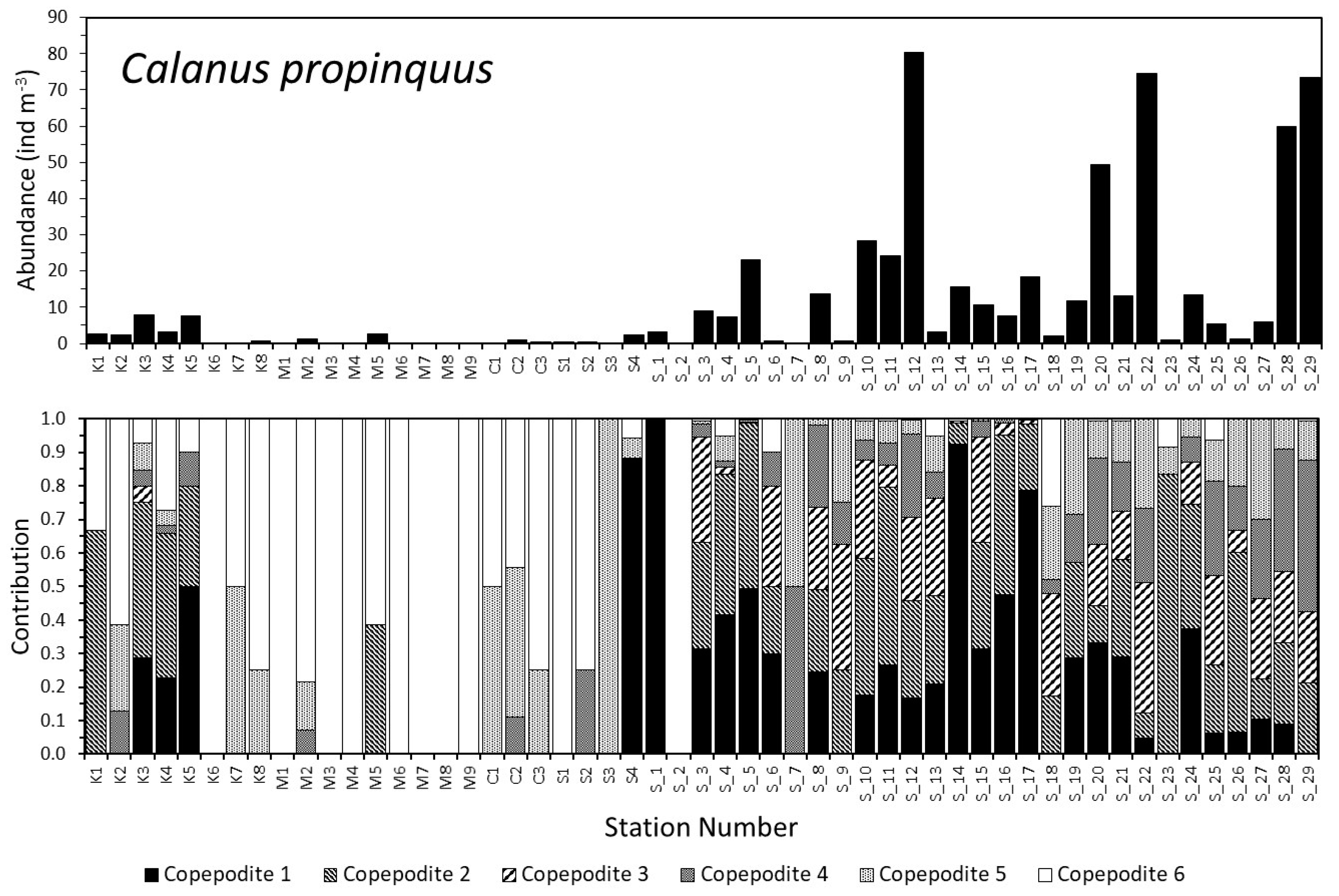
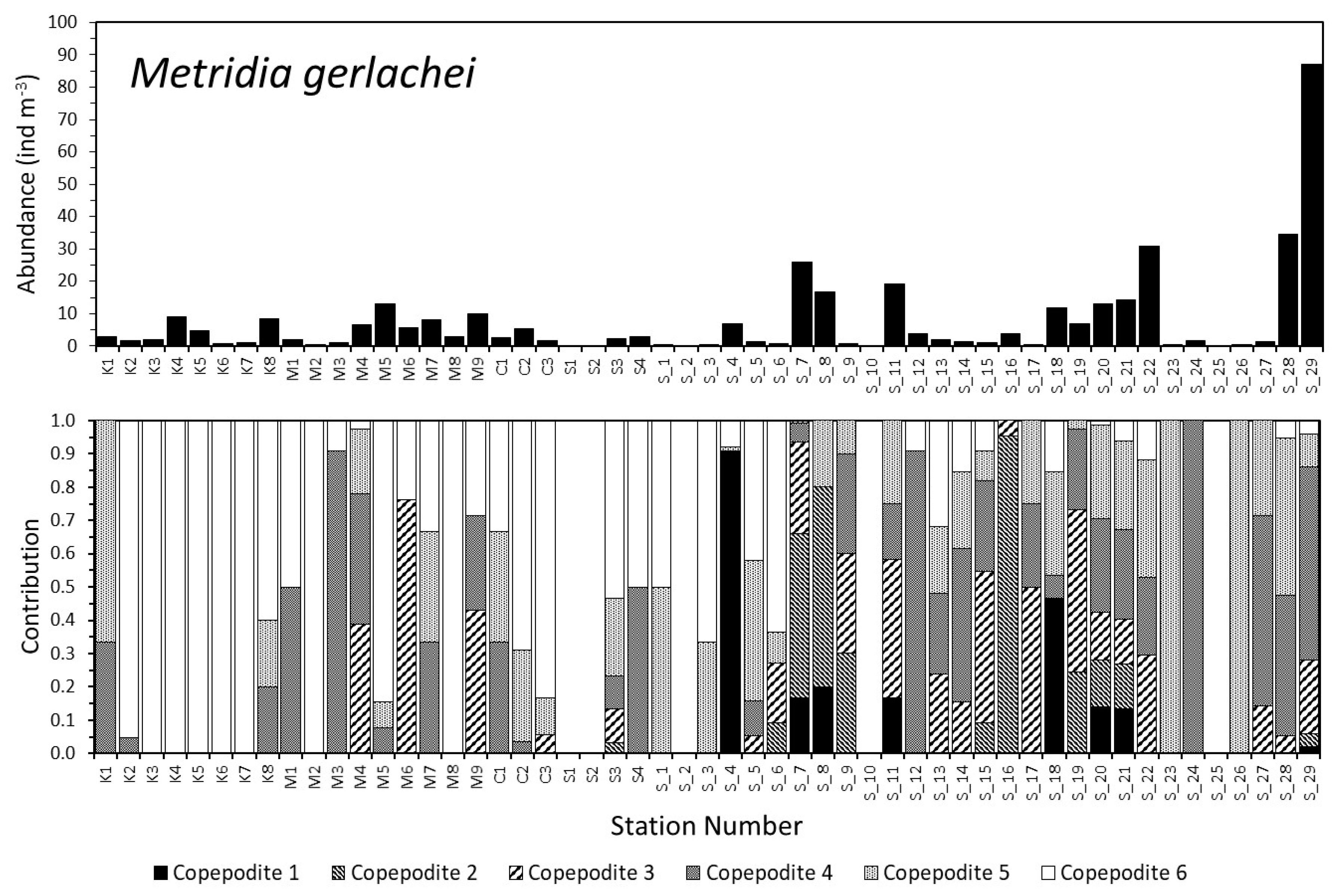
3.2. Dynamics of the Copepod Community
3.3. Community Composition Dynamics
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Perry, R.I.; Batchelder, H.P.; Mackas, D.L.; Chiba, S.; Durbin, E.; Greve, W.; Verheye, H.M. Identifying global synchronies in marine zooplankton populations: Issues and opportunities. ICES J. Mar. Sci. 2004, 61, 445–456. [Google Scholar] [CrossRef]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Robert, D.; Murphy, H.M.; Jenkins, G.P.; Fortier, L. Poor taxonomical knowledge of larval fish prey preference is impeding our ability to assess the existence of a “critical period” driving year-class strength. ICES J. Mar. Sci. 2013, 71, 2042–2052. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Landry, M.R. Zooplankton and the Ocean Carbon Cycle. Annu. Rev. Mar. Sci. 2017, 9, 413–444. [Google Scholar] [CrossRef]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of Southern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- Mantua, N.J.; Hare, S.R.; Zhang, Y.; Wallace, J.M.; Francis, R.C. A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteorol. Soc. 1997, 78, 1069–1080. [Google Scholar] [CrossRef]
- Rogers, A.; Frinault, B.; Barnes, D.; Bindoff, N.; Downie, R.; Ducklow, H.; Friedlaender, A.; Hart, T.; Hill, S.; Hofmann, E.; et al. Antarctic Futures: An Assessment of Climate-Driven Changes in Ecosystem Structure, Function, and Service Provisioning in the Southern Ocean. Annu. Rev. Mar. Sci. 2020, 12, 87–120. [Google Scholar] [CrossRef]
- Baker, A.D.C. The circumpolar continuity of Antarctic plankton species. Discov. Rep. 1954, 27, 201–2018. [Google Scholar]
- Makabe, R.; Tanimura, A.; Tamura, T.; Hirano, D.; Shimada, K.; Hashihama, F.; Fukuchi, M. Meso-zooplankton abundance and spatial distribution off Lützow-Holm Bay during austral summer 2007–2008. Polar Sci. 2017, 12, 25–33. [Google Scholar] [CrossRef]
- Smith, W.O.; Delizo, L.M.; Herbolsheimer, C.; Spencer, E. Distribution and abundance of mesozooplankton in the Ross Sea, Antarctica. Polar Biol. 2017, 40, 2351–2361. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Ruck, K.E.; Gleiber, M.R.; Garzio, L.M.; Cope, J.S.; Bernard, K.S.; Stammerjohn, S.E.; Schofield, O.M.E.; Quetin, L.B.; Ross, R.M. Long-term (1993–2013) changes in macrozooplankton off the Western Antarctic Peninsula. Deep Sea Res. Part I Oceanogr. Res. Pap. 2015, 101, 54–70. [Google Scholar] [CrossRef]
- Moriarty, R.; O’Brien, T.D. Distribution of mesozooplankton biomass in the global ocean. Earth Syst. Sci. Data 2013, 5, 45–55. [Google Scholar] [CrossRef]
- Yang, E.J.; Lee, Y.; Lee, S. Trophic interactions of micro- and mesozooplankton in the Amundsen Sea polynya and adjacent sea ice zone during austral late summer. Prog. Oceanogr. 2019, 174, 117–130. [Google Scholar] [CrossRef]
- Mizdalski, E. Weight and length data of zooplankton in the Weddell Sea in austral spring 1986 (ANT V/3). Ber. Polarforsch. 1988, 55, 1–72. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMAR-E: Plymouth, UK, 2001. [Google Scholar]
- Field, J.; Clarke, K.R.; Warwick, R. A Practical Strategy for Analysing Multispecies Distribution Patterns. Mar. Ecol. Prog. Ser. 1982, 8, 37–52. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Stevens, C.J.; Pakhomov, E.A.; Robinson, K.V.; Hall, J.A. Mesozooplankton biomass, abundance and community composition in the Ross Sea and the Pacific sector of the Southern Ocean. Polar Biol. 2014, 38, 275–286. [Google Scholar] [CrossRef]
- Swadling, K.M.; Kawaguchi, S.; Hosie, G.W. Antarctic mesozooplankton community structure during BROKE-West (30° E–80° E), January–February 2006. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 887–904. [Google Scholar] [CrossRef]
- Takahashi, K.T.; Hosie, G.W.; Odate, T. Intra-annual seasonal variability of surface zooplankton distribution patterns along a 110° E transect of the Southern Ocean in the austral summer of 2011/12. Polar Sci. 2017, 12, 46–58. [Google Scholar] [CrossRef]
- Takahashi, K.T.; Ojima, M.; Tanimura, A.; Odate, T.; Fukuchi, M. The vertical distribution and abundance of copepod nauplii and other micro- and mesozooplankton in the seasonal ice zone of Lützow-Holm Bay during austral summer 2009. Polar Biol. 2016, 40, 79–93. [Google Scholar] [CrossRef]
- Voronina, N.M.; Kosobokova, K.N.; Pakhomov, E.A. Size structure of Antarctic metazoan plankton according to united net, trawl and water bottle data. Russ. J. Aquat. Ecol. 1994, 3, 137–142. [Google Scholar]
- Voronina, N.M.; Kosobokova, K.N.; Pakhomov, E.A. Composition and biomass of summer metazoan plankton in the 0–200 m layer of the Atlantic sector of the Antarctic. Polar Biol. 1994, 14, 91–95. [Google Scholar] [CrossRef]
- Errhif, A.; Razouls, C.; Mayzaud, P. Composition and community structure of pelagic copepods in the Indian sector of the Antarctic Ocean during the end of the austral summer. Polar Biol. 1997, 17, 418–430. [Google Scholar] [CrossRef]
- Boysen-Ennen, E.; Hagen, W.; Hubold, G.; Piatkowski, U. Zooplankton biomass in the ice-covered Weddell Sea, Antarctica. Mar. Biol. 1991, 111, 227–235. [Google Scholar] [CrossRef]
- Gallienne, C.P.; Robins, D.B. Is Oithona the most important copepod in the world’s oceans? J. Plankton Res. 2001, 23, 1421–1432. [Google Scholar] [CrossRef]
- Makarov, R.R.; Men’shenina, L.L.; Spiridonov, V.A. Plankton distribution and its seasonal biological activity in the central and western parts of the Antarctic Pacific Sector. In Oceanographic and Biological Investigations of the Pacific Antarctic Sector (Biologo-Okeanographicheskie Issledovanija Tihookenaskogo Sektora Antaktiki); Makarov, R.R., Dolzhenkov, V.N., Ljubimova, T.G., Spiridonov, V.A., Tarverdieva, T.G., Eds.; VNIRO Publishers: Moscow, Russia, 1987; pp. 90–110. (In Russian) [Google Scholar]
- Pane, L.; Feletti, M.; Francomacaro, B.; Mariottini, G.L. Summer coastal zooplankton biomass and copepod community structure near the Italian Terra Nova Base (Terra Nova Bay, Ross Sea, Antarctica). J. Plankton Res. 2004, 26, 1479–1488. [Google Scholar] [CrossRef]
- Hunt, B.; Pakhomov, E.; Hosie, G.; Siegel, V.; Ward, P.; Bernard, K. Pteropods in Southern Ocean ecosystems. Prog. Oceanogr. 2008, 78, 193–221. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Torres, J.J. The zooplankton community in the vicinity of the ice edge, western Weddell Sea, March 1986. Polar Biol. 1988, 9, 79–87. [Google Scholar] [CrossRef]
- Hosie, G.W.; Cochran, T.G.; Pauly, T.; Beaumont, K.L.; Wright, S.W.; Kitchener, J.A. The zooplankton community structure of Prydz bay, January–February 1993. Proc. NIPR Symp. Polar Biol. 1997, 10, 90–133. [Google Scholar]
- Yang, G.; Li, C.; Sun, S. Inter-annual variation in summer zooplankton community structure in Prydz Bay, Antarctica, from 1999 to 2006. Polar Biol. 2011, 34, 921–932. [Google Scholar] [CrossRef]
- Voronina, N.M. Pelagic Ecosystems of the Southern Ocean; Nauka Press: Moscow, Russia, 1984; p. 206. (In Russian) [Google Scholar]
- Fedotova, V.V.; Gorlanova, O.L. Peculiarities of the mesoplankton distribution in the Ross Sea in summer 1983. In Oceanographic and Biological Investigations of the Antarctic Pacific Sector (Biologo-Okeanographicheskie Issledovanija Tihookenaskogo Sektora Antaktiki); Makarov, R.R., Dolzhenkov, V.N., Ljubimova, T.G., Spiridonov, V.A., Tarver-dieva, T.G., Eds.; VNIRO Publishers: Moscow, Russia, 1987; pp. 80–90. (In Russian) [Google Scholar]
- Makarov, R.R.; Solyankin, E.V. Abundant copepods and regional peculiarities of their seasonal development in the east area of the Weddell Gyre. In Investigations of the Weddell Gyre. Oceanographic Conditions and Peculiarities of the Development of Plankton Communities; Solyankin, E.V., Ed.; VNIRO Press: Moscow, Russia, 1990; pp. 99–125. (In Russian) [Google Scholar]
- Bundichenko, E.V.; Khromov, N.S. Mesoplankton biomass, age composition and distribution of dominant species in relation to water structure in the Commonwealth and the Cosmonaut Seas. In Interdisciplinary Investigations of Pelagic Ecosystem in the Commonwealth and Cosmonaut Seas; Lubimova, T.G., Makarov, R.R., Maslennikov, V.V., Samyshev, E.Z., Bibik, V.A., Tarverdieva, T.G., Eds.; VNIRO Publishers: Moscow, Russia, 1988; pp. 83–109. (In Russian) [Google Scholar]
- Bunichenko, E.V. Change in age composition of the abundant copepod species in the Cooperation Sea in 1977–1990. Oceanol. Russ. Acad. Sci. 1994, 34, 252–258. (In Russian) [Google Scholar]
- Yang, G.; Li, C.; Wang, Y.; Zhang, Y. Vertical profiles of zooplankton community structure in Prydz Bay, Antarctica, during the austral summer of 2012/2013. Polar Biol. 2017, 40, 1101–1114. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Perissinotto, R. Trophodynamics of the hyperiid amphipod Themisto gaudichaudii in the South Georgia region during late austral summer. Mar. Ecol. Prog. Ser. 1996, 134, 91–100. [Google Scholar] [CrossRef]
- Froneman, P.W.; Pakhomov, E.A.; Treasure, A. Trophic importance of the hyperiid amphipod, Themisto gaudichaudii, in the Prince Edward Archipelago (Southern Ocean) ecosystem. Polar Biol. 2000, 23, 429–436. [Google Scholar] [CrossRef]
- Kruse, S.; Pakhomov, E.A.; Hunt, B.; Chikaraishi, Y.; Ogawa, N.; Bathmann, U. Uncovering the trophic relationship between Themisto gaudichaudii and Salpa thompsoni in the Antarctic Polar Frontal Zone. Mar. Ecol. Prog. Ser. 2015, 529, 63–74. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station No. | Date | Time | Latitude (South) | Longitude (West) | Depth Sampled (m) | Volume Filtered, m3 | SurfaceT °C |
|---|---|---|---|---|---|---|---|
| K1 | 11/17/2017 | 17:00 | 68.187 | 112.308 | 103 | 11.0 | 0.0 |
| K2 | 11/21/2017 | 17:40 | 69.520 | 111.607 | 100 | 13.0 | −1.0 |
| K3 | 11/26/2017 | 17:00 | 70.672 | 111.422 | 104 | 10.5 | −1.7 |
| K4 | 12/3/2017 | 13:50 | 70.717 | 111.620 | 111 | 13.5 | −1.8 |
| K5 | 12/9/2017 | 17:20 | 70.917 | 113.965 | 125 | 13.0 | −1.7 |
| K6 | 12/14/2017 | 17:20 | 72.430 | 117.173 | 118 | 12.0 | −1.8 |
| K7 | 12/16/2017 | 12:35 | 73.932 | 117.172 | 141 | 20.0 | −1.6 |
| K8 | 1/28/2018 | 18:30 | 71.910 | 120.702 | 109 | 12.0 | −1.6 |
| M1 | 11/28/2017 | 11:25 | 65.503 | 177.599 | 101 | 10.5 | nr |
| M2 | 1/16/2018 | 10:00 | 72.088 | 176.695 | 100 | 11.0 | nr |
| M3 | 1/16/2018 | 17:00 | 72.872 | 179.983 | 100 | 11.0 | nr |
| M4 | 1/17/2018 | 0:30 | 73.177 | 177.594 | 100 | 11.6 | nr |
| M5 | 1/18/2018 | 9:10 | 75.008 | 163.665 | 100 | 10.0 | nr |
| M6 | 1/18/2018 | 18:20 | 75.034 | 157.864 | 100 | 10.5 | nr |
| M7 | 1/19/2018 | 10:50 | 74.638 | 147.513 | 100 | 11.0 | nr |
| M8 | 1/21/2018 | 20:10 | 74.068 | 135.990 | 100 | 10.5 | nr |
| M9 | 1/24/2018 | 19:50 | 74.068 | 131.488 | 100 | 10.5 | nr |
| C1 | 12/12/2017 | 22:45 | 65.380 | 178.397 | 100 | 11.6 | nr |
| C2 | 1/14/2018 | 16:20 | 72.648 | 176.247 | 100 | 10.2 | nr |
| C3 | 1/18/2018 | 9:40 | 74.142 | 139.456 | 100 | 11.6 | nr |
| S1 | 11/29/2017 | 18:00 | 64.573 | 171.133 | 113 | 12.0 | nr |
| S2 | 12/1/2017 | 15:00 | 64.593 | 171.072 | 113 | 13.0 | nr |
| S3 | 1/11/2018 | 21:35 | 74.112 | 136.119 | 118 | 13.0 | nr |
| S4 | 1/28/2018 | 1:40 | 72.583 | 121.128 | 99 | 14.0 | nr |
| S_1 | 2/27/2018 | 10:00 | 61.905 | 37.583 | 118 | 12.0 | 0.2 |
| S_2 | 2/27/2018 | 14:30 | 62.000 | 36.833 | 122 | 13.0 | 0.0 |
| S_3 | 2/27/2018 | 15:30 | 61.999 | 36.502 | 121 | 14.0 | 0.0 |
| S_4 | 2/27/2018 | 19:30 | 61.583 | 35.667 | 122 | 13.0 | 0.0 |
| S_5 | 2/27/2018 | 21:30 | 61.430 | 34.247 | 114 | 14.0 | 0.0 |
| S_6 | 2/27/2018 | 23:00 | 61.350 | 34.673 | 112 | 13.0 | 0.0 |
| S_7 | 2/28/2018 | 9:15 | 60.932 | 35.067 | 121 | 14.0 | 0.7 |
| S_8 | 2/28/2018 | 11:15 | 60.833 | 36.167 | 118 | 12.0 | 1.5 |
| S_9 | 2/28/2018 | 13:45 | 61.169 | 36.180 | 113 | 12.0 | 1.4 |
| S_10 | 2/28/2018 | 15:30 | 61.257 | 36.669 | 113 | 12.0 | 1.6 |
| S_11 | 2/28/2018 | 18:00 | 61.333 | 37.250 | 113 | 12.5 | 1.6 |
| S_12 | 2/28/2018 | 20:30 | 61.500 | 36.833 | 104 | 12.0 | 1.5 |
| S_13 | 2/28/2018 | 21:30 | 61.500 | 36.417 | 98 | 12.0 | 1.5 |
| S_14 | 3/2/2018 | 12:40 | 61.667 | 39.167 | 109 | 11.0 | 1.4 |
| S_15 | 3/2/2018 | 14:00 | 61.750 | 38.667 | 109 | 12.0 | 1.4 |
| S_16 | 3/2/2018 | 16:20 | 61.998 | 39.168 | 106 | 11.0 | 1.4 |
| S_17 | 3/2/2018 | 18:00 | 61.998 | 38.667 | 103 | 11.0 | 1.4 |
| S_18 | 3/2/2018 | 21:10 | 62.017 | 40.634 | 103 | 11.0 | 1.4 |
| S_19 | 3/31/2018 | 17:30 | 60.002 | 34.007 | 116 | 12.0 | 0.7 |
| S_20 | 3/31/2018 | 19:30 | 60.333 | 34.417 | 108 | 11.0 | 0.7 |
| S_21 | 3/31/2018 | 21:25 | 60.658 | 34.678 | 91 | 10.5 | 0.7 |
| S_22 | 4/2/2018 | 21:10 | 60.583 | 35.333 | 100 | 11.0 | 0.8 |
| S_23 | 4/8/2018 | 5:30 | 59.592 | 35.422 | 100 | 11.0 | 0.9 |
| S_24 | 4/8/2018 | 9:20 | 59.672 | 36.167 | 104 | 12.0 | 1.0 |
| S_25 | 4/10/2018 | 7:30 | 61.250 | 37.833 | 113 | 12.0 | 1.2 |
| S_26 | 4/10/2018 | 13:20 | 60.916 | 38.298 | 108 | 11.0 | 1.2 |
| S_27 | 4/10/2018 | 15:40 | 60.666 | 37.833 | 106 | 11.0 | 1.3 |
| S_28 | 4/10/2018 | 20:20 | 60.255 | 36.752 | 100 | 11.0 | 1.4 |
| S_29 | 4/10/2018 | 21:40 | 60.207 | 37.167 | 94 | 11.5 | 1.4 |
| Species | Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | Cluster 5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | B | A | B | A | B | A | B | A | B | |
| Calanoides acutus | 0.4 | 5.3 | 9.9 | 50.4 | 10.3 | 27.6 | 3.4 | 6.8 | 5.2 | 36.6 |
| Metridia gerlachei | 0.1 | 0.4 | 3.3 | 6.3 | 4.4 | 5.1 | 9.5 | 6.1 | 1.1 | 1.4 |
| Pleuromamma spp. | 0 | 0 | 0.4 | <0.1 | 0 | 0 | <0.1 | <0.1 | 0.1 | <0.1 |
| Ctenocalanus/Clausocalanus | 13.6 | 20.2 | 10 | 5.5 | 6.2 | 1.8 | 15.9 | 8.1 | 8.2 | 10.3 |
| Paraeuchaeta spp. | 0 | 0 | 0.7 | 0.5 | 0.9 | 0.5 | 0.1 | 1.7 | 0.1 | 0.1 |
| Euchirella rostromagna | 0 | 0 | 0 | 0 | 0 | 0 | <0.1 | 0.1 | <0.1 | 0.1 |
| Calanus propinquus | 0 | 1.5 | 4.6 | 18.8 | 0.4 | 2.5 | 15.4 | 51.3 | 3.8 | 12.4 |
| Calanus simillimus | 0 | 1.8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rhincalanus gigas | 0.4 | 14.2 | 0.1 | 0.9 | <0.1 | 0.2 | <0.1 | 0.1 | 0.1 | 0.7 |
| Oithona spp. | 62.4 | 34.9 | 50.9 | 10.6 | 30.2 | 3.2 | 28.9 | 5.5 | 43.9 | 20.8 |
| Oncaea spp. | 3.5 | 1.1 | 2.3 | 0.3 | 35.1 | 2.1 | 1.1 | 0.1 | 5.5 | 1.4 |
| Microsetella spp. | 1.4 | 0.5 | 0.1 | <0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Thysanoessa macrura | 0.6 | 4.1 | 0.9 | 1.9 | <0.1 | 0.1 | 0.5 | 5.7 | 0.1 | 2.2 |
| Euphausia crystallorophias | 0 | 0 | 0 | 0 | <0.1 | 1.8 | 0 | 0 | 0 | 0 |
| Euphausia superba | 0 | 0 | 0.5 | 0.1 | 0 | 0 | 21 | 10.8 | <0.1 | 0.8 |
| Crustacea eggs | <0.1 | <0.1 | 1.9 | <0.1 | <0.1 | <0.1 | 1 | <0.1 | 7.1 | <0.1 |
| Copepoda egg clusters | 0 | 0 | 0 | 0 | <0.1 | <0.1 | 0 | 0 | <0.1 | <0.1 |
| Copepoda nauplii | 17.3 | 0.2 | 11.9 | 0.1 | 9.5 | 0 | 2.5 | <0.1 | 22 | 0.2 |
| Themisto gaudichaudii | 0 | 0 | <0.1 | 1.8 | 0 | 0 | <0.1 | 2.3 | <0.1 | 9.4 |
| Vibilia antarctica | <0.1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Primno macropa | <0.1 | <0.1 | <0.1 | <0.1 | 0 | 0 | 0 | 0 | <0.1 | 0.1 |
| Hyperiella dilatata | 0 | 0.1 | 0 | 0 | <0.1 | 0.4 | 0 | 0 | 0 | 0 |
| Eusirus sp. | 0 | 0 | 0 | 0 | <0.1 | 0.2 | 0 | 0 | 0 | 0 |
| Nematocarcinus spp. | 0 | 0 | <0.1 | <0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ostracoda | <0.1 | 0.2 | <0.1 | 0.1 | 0.5 | 1 | 0.1 | 0.4 | 0.1 | 0.3 |
| Spongiobranchaea australis | 0 | 0 | <0.1 | <0.1 | <0.1 | <0.1 | 0 | 0 | 0 | 0 |
| Clione antarctica | <0.1 | 0.6 | <0.1 | 0.1 | <0.1 | <0.1 | 0 | 0 | 0.1 | <0.1 |
| Limacina helicina | <0.1 | 0.2 | 0.7 | 0.6 | 0.1 | 0.2 | 0.3 | 0.2 | 1.2 | 0.9 |
| Rhinchonerella bongraini | <0.1 | 0.2 | <0.1 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pelagobia longicerrata | 0.1 | 0.1 | 1.3 | 0.6 | 1.4 | 0.5 | 0.1 | 0.2 | 0.9 | 1.2 |
| Tomopteris spp. | <0.1 | <0.1 | <0.1 | <0.1 | 0.1 | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| Sagitta gazellae | <0.1 | <0.1 | <0.1 | 0.1 | <0.1 | 0.2 | 0 | 0 | <0.1 | <0.1 |
| Eukrohnia hamata | <0.1 | 0.3 | 0.1 | 0.7 | 0.2 | 0.6 | 0.1 | 0.3 | 0.1 | 0.5 |
| Medusae | <0.1 | 0.3 | <0.1 | 0.4 | 0 | 0 | 0 | 0 | <0.1 | 0.5 |
| Appendicularia | 0.1 | 0.1 | 0.1 | <0.1 | 0.5 | 0.2 | <0.1 | <0.1 | 0.4 | <0.1 |
| Salpa thompsoni | 0.1 | 7.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pyrostephos vanhoeffeni | <0.1 | 4.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calicopsis borchgervinki | 0 | 0 | 0 | 0 | <0.1 | 0.2 | 0 | 0 | 0 | 0 |
| Calianira sp. | 0 | 0 | 0 | 0 | <0.1 | 51.7 | <0.1 | 0.1 | 0 | 0 |
| Total abundance (mean ± 1SD) | 1620 ± 1124 | 121.0 ± 148.3 | 164.2 ± 86.6 | 317.5 ± 133.1 | 419.1 ± 218.8 | |||||
| Total biomass (mean ± 1SD) | 26.2 ± 20.1 | 5.2 ± 4.1 | 13.8 ± 22.6 | 14.9 ± 9.0 | 8.0 ± 8.3 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakhomov, E.A.; Pshenichnov, L.K.; Krot, A.; Paramonov, V.; Slypko, I.; Zabroda, P. Zooplankton Distribution and Community Structure in the Pacific and Atlantic Sectors of the Southern Ocean during Austral Summer 2017–18: A Pilot Study Conducted from Ukrainian Long-Liners. J. Mar. Sci. Eng. 2020, 8, 488. https://doi.org/10.3390/jmse8070488
Pakhomov EA, Pshenichnov LK, Krot A, Paramonov V, Slypko I, Zabroda P. Zooplankton Distribution and Community Structure in the Pacific and Atlantic Sectors of the Southern Ocean during Austral Summer 2017–18: A Pilot Study Conducted from Ukrainian Long-Liners. Journal of Marine Science and Engineering. 2020; 8(7):488. https://doi.org/10.3390/jmse8070488
Chicago/Turabian StylePakhomov, Evgeny A., Leonid K. Pshenichnov, Anatoly Krot, Valery Paramonov, Ilia Slypko, and Pavel Zabroda. 2020. "Zooplankton Distribution and Community Structure in the Pacific and Atlantic Sectors of the Southern Ocean during Austral Summer 2017–18: A Pilot Study Conducted from Ukrainian Long-Liners" Journal of Marine Science and Engineering 8, no. 7: 488. https://doi.org/10.3390/jmse8070488
APA StylePakhomov, E. A., Pshenichnov, L. K., Krot, A., Paramonov, V., Slypko, I., & Zabroda, P. (2020). Zooplankton Distribution and Community Structure in the Pacific and Atlantic Sectors of the Southern Ocean during Austral Summer 2017–18: A Pilot Study Conducted from Ukrainian Long-Liners. Journal of Marine Science and Engineering, 8(7), 488. https://doi.org/10.3390/jmse8070488