Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures





Abstract
1. Introduction
2. Materials and Methods
2.1. Design of Experiment
2.2. Animal Identification and Rearing
2.3. Preparation of Feeds
2.4. Feeding Tests
2.5. Statistical Analyses
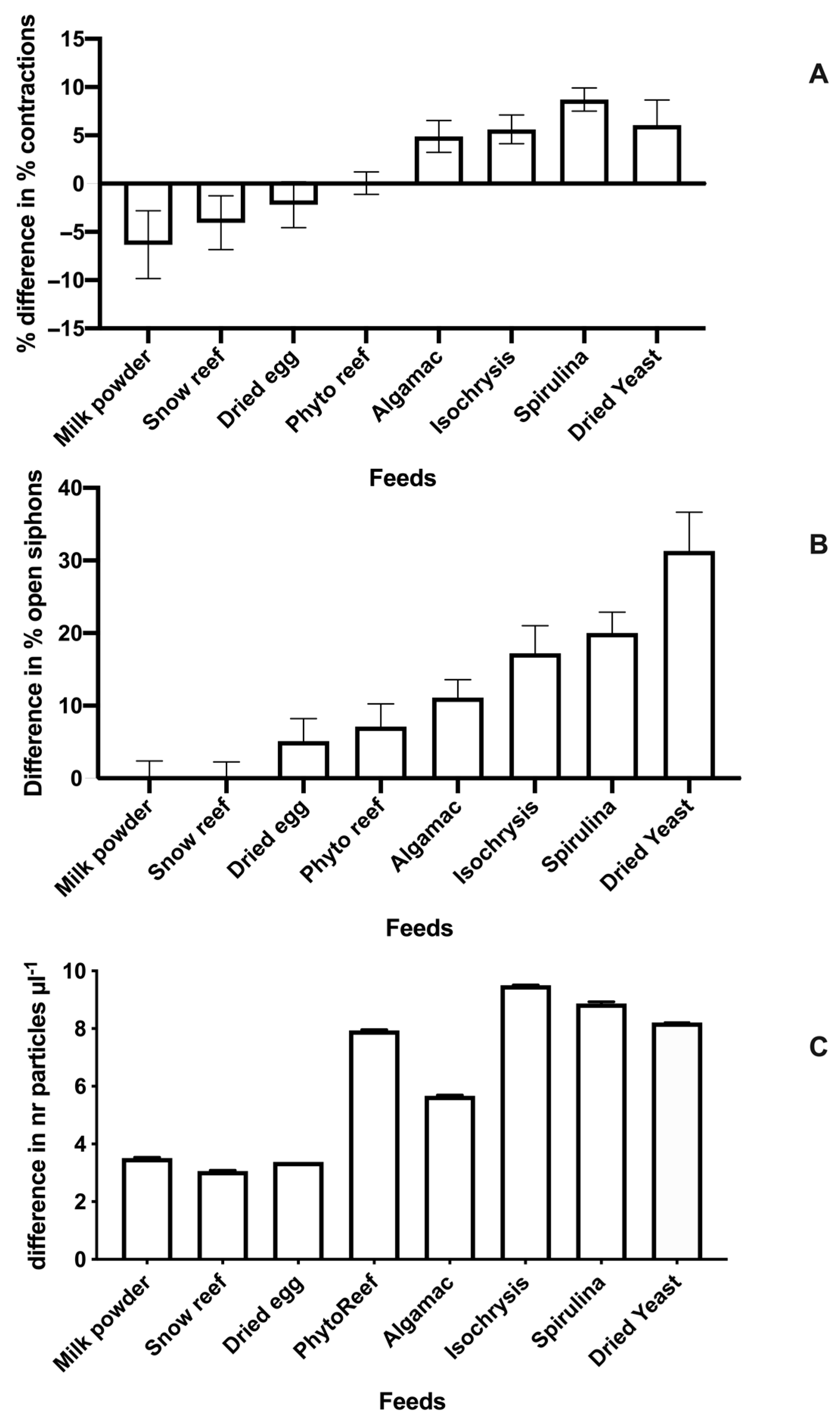
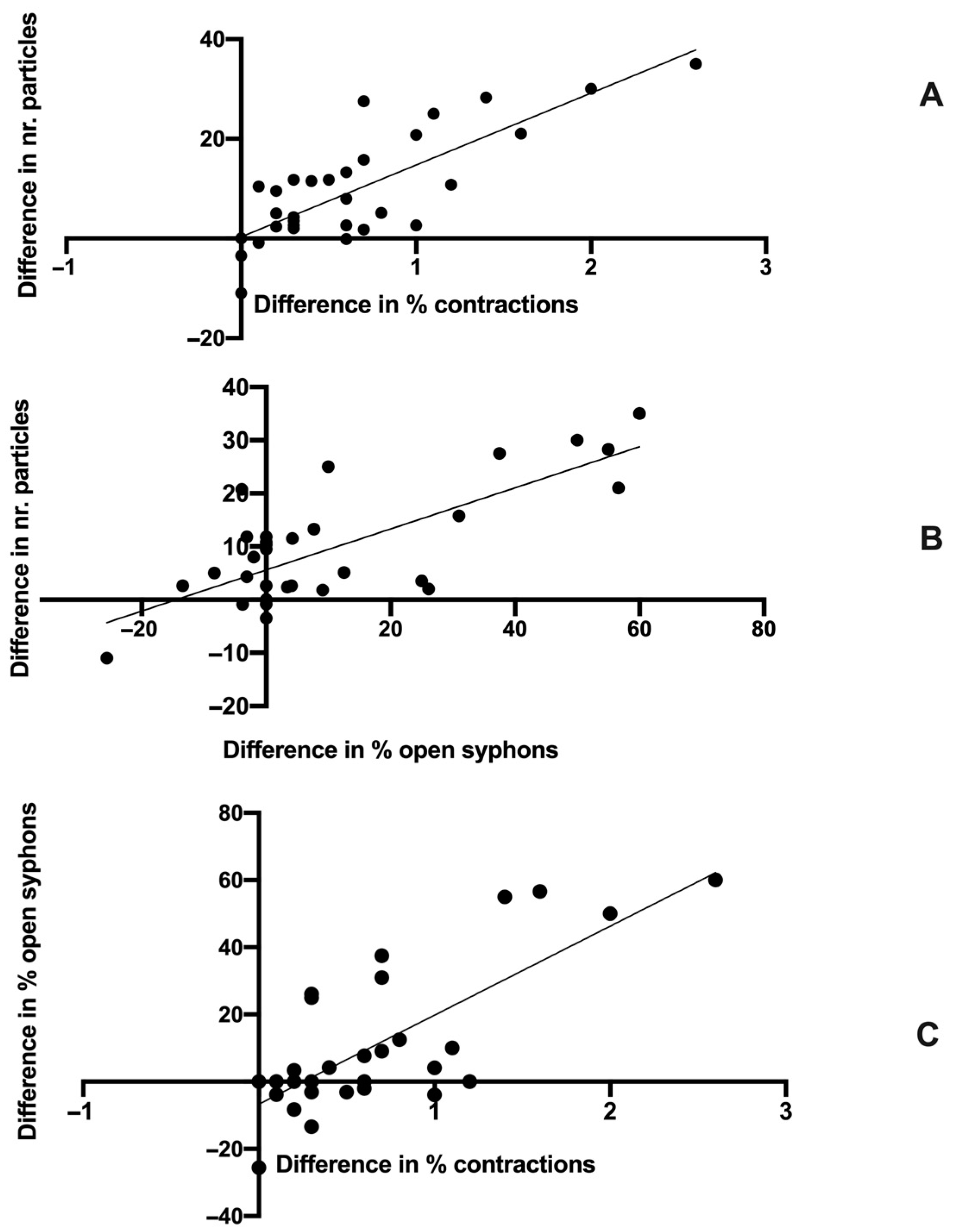
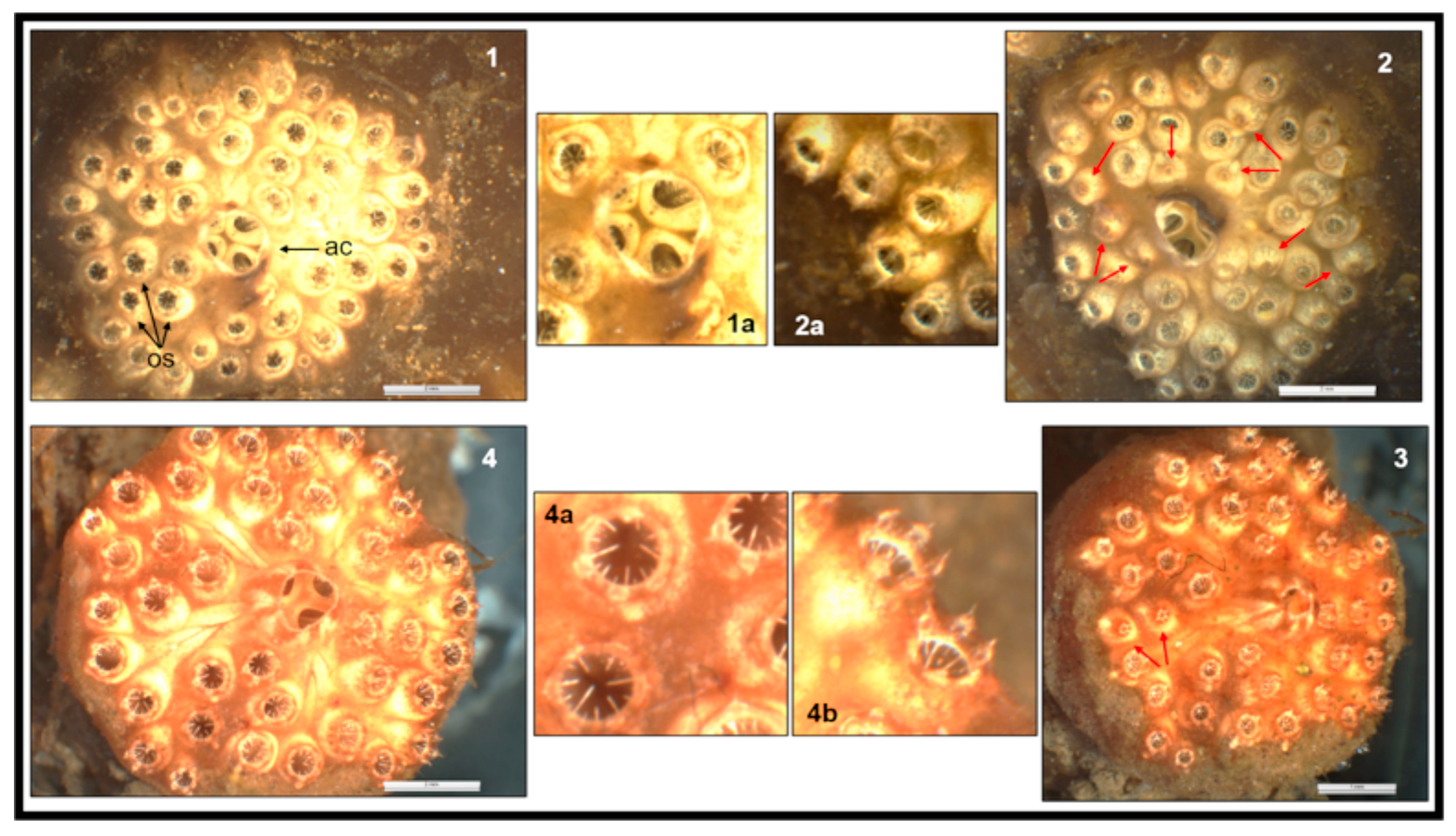
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Chadwick-Furman, N.E.; Weissman, I.L. Life histories and senescence of Botryllus schlosseri (Chordata, Ascidiacea) in Monterey Bay. Biol. Bull. 1995, 189, 36–41. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Voskoboynik, A.; Weissman, I.L. Botryllus schlosseri, an emerging model for the study of ageing, stem cells, and mechanisms of regeneration. Invertebr. Reprod. Dev. 2015, 59, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Austad, S. Recent advances in vertebrate ageing research. Ageing Cell 2009, 9, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Kott, P. The Australian ascidians. I. Phlebobranchia and Stolidobranchia. Mem. Queensl. Mus. 1985, 23, 1–440. [Google Scholar]
- Rinkevich, B.; Rabinowitz, C. In vitro culture of blood cells from the colonial protochordate Botryllus schlosseri. In Vitro Cell. Dev. Biol. 1993, 29, 79–85. [Google Scholar] [CrossRef]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Hervé, P. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef]
- Zupo, V.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Procaccini, G. Temporal variations in the spatial distribution of shoot density in a Posidonia oceanica meadow and patterns of genetic diversity. Mar. Ecol. 2006, 27, 328–338. [Google Scholar] [CrossRef]
- Grosholz, E. Small spatial-scale differentiation among populations of an introduced colonial invertebrate. Oecologia 2001, 129, 58–64. [Google Scholar] [CrossRef]
- Simoncini, M.; Miller, R.J. Feeding preference of Strongylocentrotus droebachiensis (Echinoidea) for a dominant native ascidian, Aplidium glabrum, relative to the invasive ascidian Botrylloides violaceus. J. Exp. Mar. Biol. Ecol. 2007, 342, 93–98. [Google Scholar] [CrossRef]
- Hirose, E. Colonial allorecognition, haemolytic rejection, and viviparity in Botryllid ascidians. Zool. Sci. 2003, 20, 387–394. [Google Scholar] [CrossRef]
- Berrill, N.J. The development and growth of Ciona. J. Mar. Biol. Assoc. 1947, 26, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Epelbaum, A.; Therriault, T.W.; Paulson, A.; Pearce, C.M. Botryllid tunicates: Culture techniques and experimental procedures. Aquat. Invasions 2009, 4, 111–120. [Google Scholar] [CrossRef]
- Zega, G.; Pennati, R.; Candiani, S.; Pestarino, M.; De Bernardi, F. Solitary ascidians embryos (Chordata, Tunicata) as model organisms for testing coastal pollutant toxicity. Invertebr. Surviv. J. 2009, 6, 29–34. [Google Scholar]
- Hyams, Y.; Paza, G.; Rabinowitza, C.; Rinkevich, B. Insights into the unique torpor of Botrylloides leachi, a colonial urochordate. Dev. Biol. 2007, 428, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Manni, L.; Gasparini, F.; Hotta, K.; Ishizuka, K.J.; Ricci, L.; Tiozzo, S.; Voskoboynik, A.; Dauga, D. Ontology for the asexual development and anatomy of the colonial chordate Botryllus schlosseri. PLoS ONE 2014, 9, e96434. [Google Scholar] [CrossRef]
- Ballarin, L.; Burighel, P.; Cima, F. A tale of death and life: Natural apoptosis in the colonial ascidian Botryllus schlosseri (Urochordata, Ascidiacea). Curr. Pharm. Des. 2008, 14, 138–147. [Google Scholar] [CrossRef]
- Rinkevich, B.; Shapira, M. An improved diet for inland broodstock and the establishment of an inbred line from a colonial sea squirt (Ascidiacea). Aquat. Living Resourc. 1998, 11, 163–171. [Google Scholar] [CrossRef]
- Brunetti, R.; Mastrototaro, F. Botrylloides pizoni, a new species of Botryllinae (Ascidiacea) from the Mediterranean Sea. Zootaxa 2012, 3258, 28–36. [Google Scholar] [CrossRef]
- Brunetti, R.; Beghi, L.; Bressan, M.; Marin, M.G. Combined effects of temperature and salinity on colonies of Botryllus schlosseri and Botrylloides leachi (Ascidiacea) from the Venetian lagoon. Mar. Ecol. Prog. Ser. 1980, 2, 303–314. [Google Scholar] [CrossRef]
- Bone, Q.; Carré, C.; Chang, P. Tunicate feeding filters. J. Mar. Biol. Assoc. 2003, 83, 907–919. [Google Scholar] [CrossRef]
- Brunetti, R. Botryllid species (Tunicata, Ascidiacea) from the Mediterranean coast of Israel, with some consideration on the systematic of Botryllinae. Zootaxa 2009, 2289, 18–32. [Google Scholar] [CrossRef]
- Sawada, H.; Yokosawa, H.; Lambert, C.C. (Eds.) The Biology of Ascidians; Springer: Tokyo, Japan, 2001; p. 470. [Google Scholar]
- Kloc, M.; Kubiak, J.K. Marine Organisms as Model Systems in Biology and Medicine; Springer: Houston, TX, USA, 2018; p. 65. [Google Scholar]
- Liu, L.; Xiang, J.; Dong, B.; Natarajan, P.; Yu, K.; Cai, N. Ciona intestinalis as an emerging model organism: Its regeneration under controlled conditions and methodology for egg dechorionation. J. Zhejiang Univ. Sci. 2006, 7, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Bay-Nouailhat, A.; Bay-Nouailhat, W.; Gasparini, F.; Brunetti, R. Botrylloides crystallinus n. sp., a new Botryllinae Adams & Adams, 1858 (Ascidiacea) from Mediterranean Sea. Zoosystema 2020, 42, 131–138. [Google Scholar]
- Degasperi, V.; Gasparini, F.; Shimeld, S.M.; Sinigaglia, C.; Burighel, P.; Manni, L. Muscle differentiation in a colonial ascidian: Organisation, gene expression and evolutionary considerations. BMC Dev. Biol. 2009, 9, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Mackie, G.O.; Burighel, P.; Caicci, F.; Manni, L. Innervation of ascidian siphons and their responses to stimulation. Can. J. Zool. 2006, 84, 1146–1162. [Google Scholar] [CrossRef]
- Lambert, G. New records of ascidians from the NE Pacific: A new species of Trididemnum, range extension and redescription of Aplidiopsis pannosum (Ritter, 1899) including its larva, and several nonindigenous species. Zoosystema 2003, 25, 665–679. [Google Scholar]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking climate change and biological invasions: Ocean warming facilitates nonindigenous species invasions. Proc. Natl. Acad. Sci. USA 2002, 99, 15497–15500. [Google Scholar] [CrossRef]
- Young, C.M. Selection of predator-free settlement sites by larval ascidians. Ophelia 1989, 30, 131–140. [Google Scholar] [CrossRef]
- Pisut, D.P.; Pawlik, J.R. Antipredatory chemical defenses of ascidians: Secondary metabolites or inorganic acids. J. Exp. Mar. Biol. Ecol. 2002, 270, 203–214. [Google Scholar] [CrossRef]
- Burighel, P.; Lane, N.J.; Gasparini, F.; Tiozzo, S.; Zaniolo, G.; Candia Carnevali, M.D.; Manni, L. Novel secondary sensory cell organ in Ascidians: In search of the ancestor of the vertebrate lateral line. J. Comp. Neurol. 2003, 461, 236–249. [Google Scholar] [CrossRef]
- Mackie, G.O.; Burighel, P. The nervous system in adult tunicates: Current research directions. Can. J. Zool. 2005, 83, 151–183. [Google Scholar] [CrossRef]
- Manni, L.; Mackie, G.O.; Caicci, F.; Zaniolo, G.; Burighel, P. Coronal organ of ascidians and the evolutionary significance of secondary sensory cells in chordates. J. Comp. Neurol. 2006, 495, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Bagwell, C.E.; Abernathy, A.; Barnwell, R.; Milliken, C.E.; Noble, P.A.; Dale, T.; Beauchesne, K.R.; Moeller, P.D.R. Discovery of bioactive metabolites in biofuel microalgae that offer protection against predatory bacteria. Front. Microbiology 2016, 7, 516–525. [Google Scholar]
- Varrella, S.; Romano, G.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. First morphological and molecular evidence of the negative impact of diatom-derived hydroxyacids on the sea urchin Paracentrotus lividus. Toxicol. Sci. 2016, 151, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Armsworthy, S.L.; MacDonald, B.A.; Ward, J.A. Feeding activity, absorption efficiency and suspension feeding processes in the ascidian, Halocynthia pyriformis (Stolidobranchia: Ascidiacea): Responses to variations in diet quantity and quality. J. Exp. Mar. Biol. Ecol. 2001, 260, 41–69. [Google Scholar] [CrossRef]
- Nappo, M.; Berkov, S.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Bastida, J. Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS. J. Appl. Phycol. 2009, 21, 295–306. [Google Scholar] [CrossRef]
- Raniello, R.; Iannicelli, M.M.; Nappo, M.; Avila, C.; Zupo, V. Production of Cocconeis neothumensis (Bacillariophyceae) biomass in batch cultures and bioreactors for biotechnological applications: Light and nutrient requirements. J. Appl. Phycol. 2007, 19, 383–391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Feed Component | Content | % Proteins | % Lipids | % Carbohydrates | % Fibres | % Ashes | % Moisture |
|---|---|---|---|---|---|---|---|
| Milk Powder (Ristora) | Dried skimmed milk | 34.00 | 1.20 | 56.00 | - | - | - |
| SnowReef® (SHG) | Yeast (Saccharomices cerevisiae) and torula yeast, microalgae, squid pulp, fish oil at high content of HUFA | 42.00 | 7.00 | - | - | 7.00 | - |
| Freeze-dried egg | Yolk and albumen | 12.56 | 9.51 | 0.72 | - | - | - |
| PhytoReef® (SHG) | Principal component: lyophilised Chlorella sp. | 58.00 | 11.00 | - | 19.00 | - | < 10.00 |
| AlgaMac® Protein Plus® (PTAqua) | Dried microalgae | 42.90 | 21.00 | 17.50 | - | 12.40 | 6.00 |
| Isochrysis galbana* | Live axenic culture of microalgae | 29.00 | 23.00 | 12.90 | - | - | - |
| Dry Spirulina® (SHG) | 100% freeze-dried microalgae | 60.00 | 4.00 | - | 7.00 | 7.00 | - |
| Dried Yeast (Selex) | Dried brewer yeast | 19.00 | 19.00 | 72.00 | 2.50 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregorin, C.; Musco, L.; Somma, E.; Zupo, V. Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures. J. Mar. Sci. Eng. 2020, 8, 1021. https://doi.org/10.3390/jmse8121021
Gregorin C, Musco L, Somma E, Zupo V. Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures. Journal of Marine Science and Engineering. 2020; 8(12):1021. https://doi.org/10.3390/jmse8121021
Chicago/Turabian StyleGregorin, Chiara, Luigi Musco, Emanuele Somma, and Valerio Zupo. 2020. "Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures" Journal of Marine Science and Engineering 8, no. 12: 1021. https://doi.org/10.3390/jmse8121021
APA StyleGregorin, C., Musco, L., Somma, E., & Zupo, V. (2020). Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures. Journal of Marine Science and Engineering, 8(12), 1021. https://doi.org/10.3390/jmse8121021