Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter sp. TT1 by a Chemical Dispersant

 ,
, 
 ,
,  and
and 
Abstract
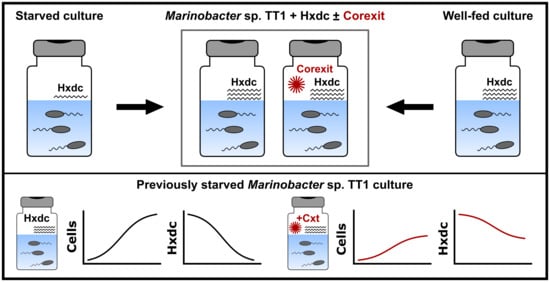
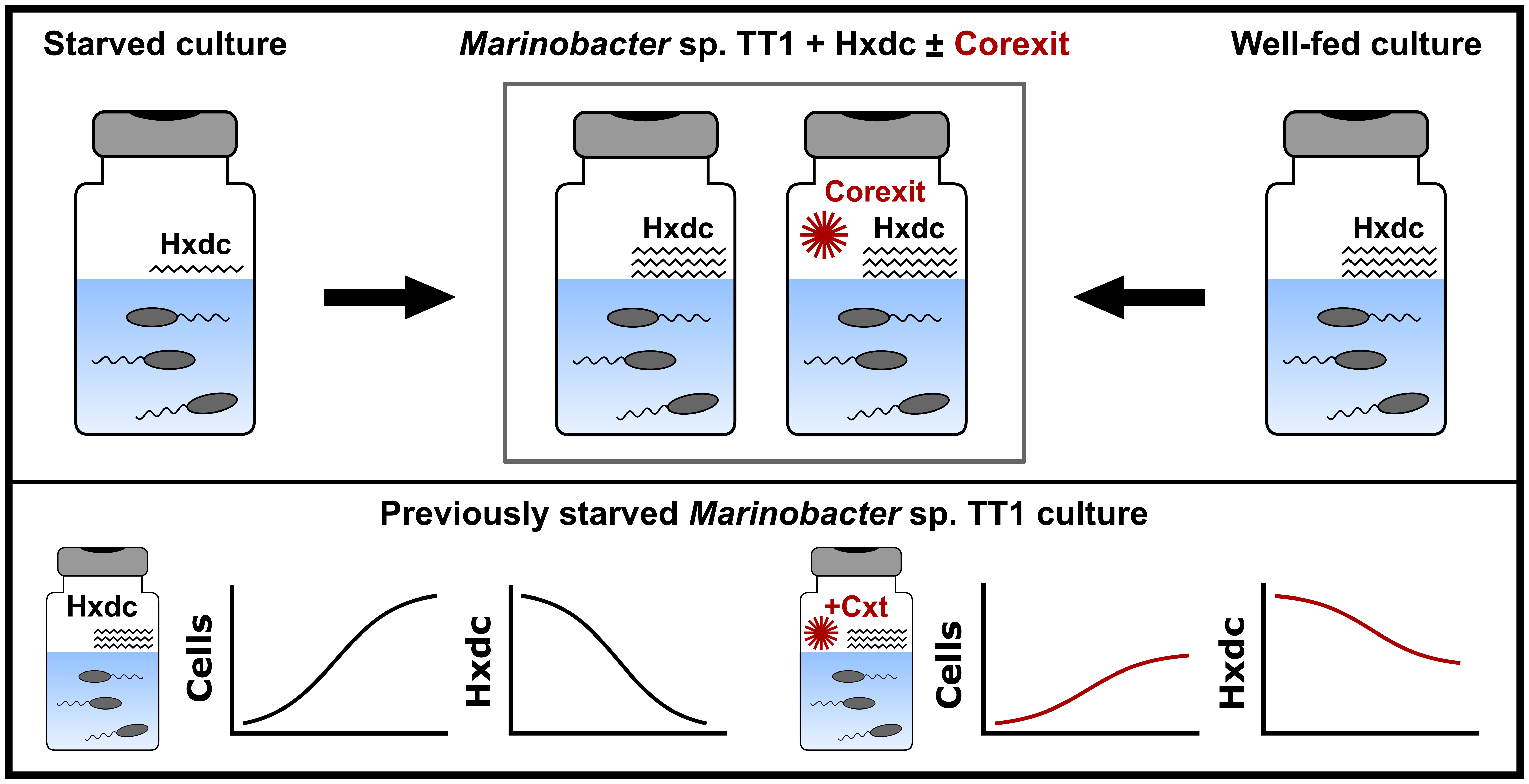{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Pre-Adaptation of Cultures
2.2. Experimental Setup
2.3. Cell Counts
2.4. Hydrocarbon Quantification
2.5. Data Analysis
3. Results and Discussion
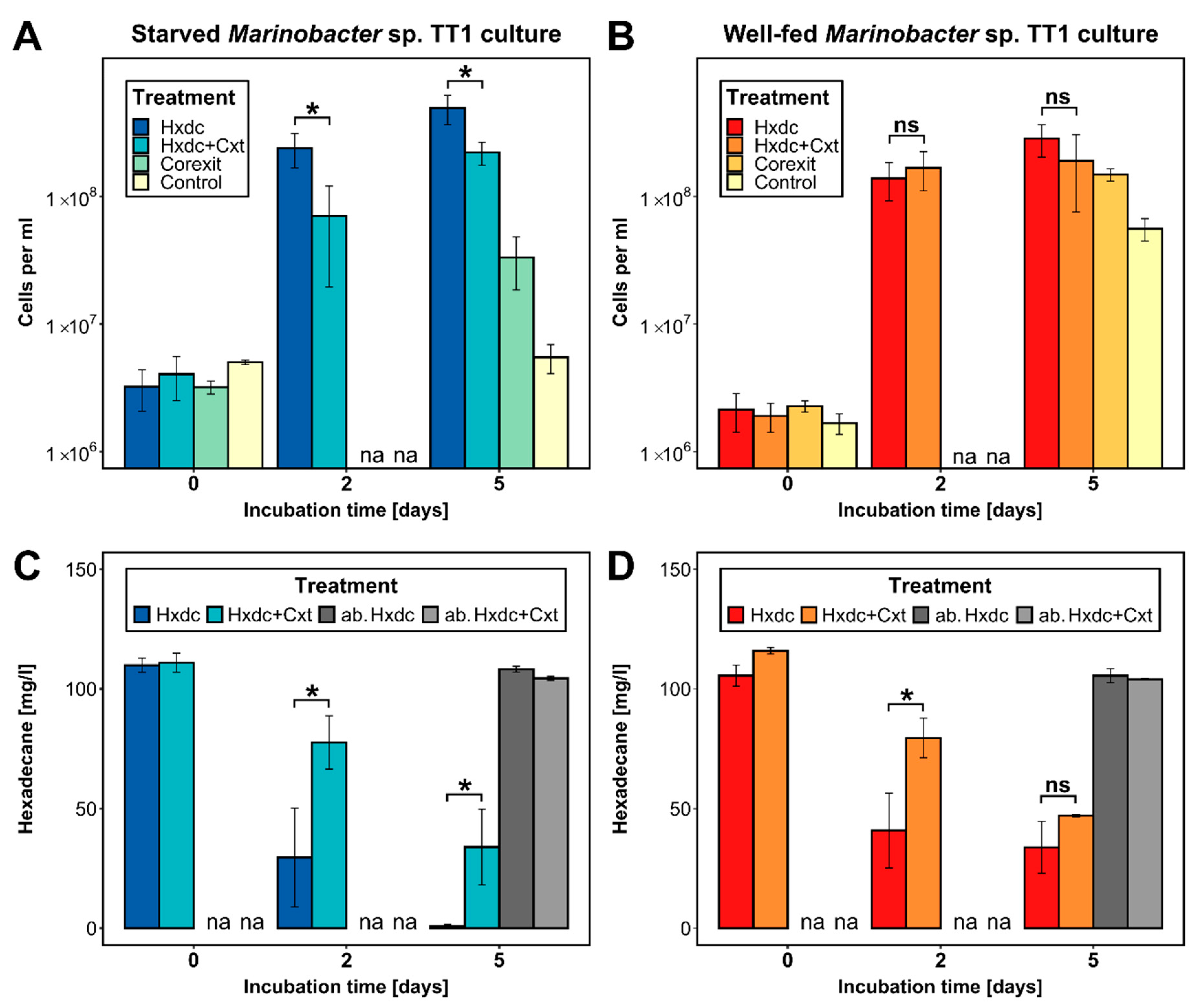
3.1. Rapid Response by Starved Marinobacter sp. TT1 to a High Hydrocarbon Pulse
3.2. Only Starved Marinobacter sp. TT1 Was Consistently Inhibited after Corexit Exposure
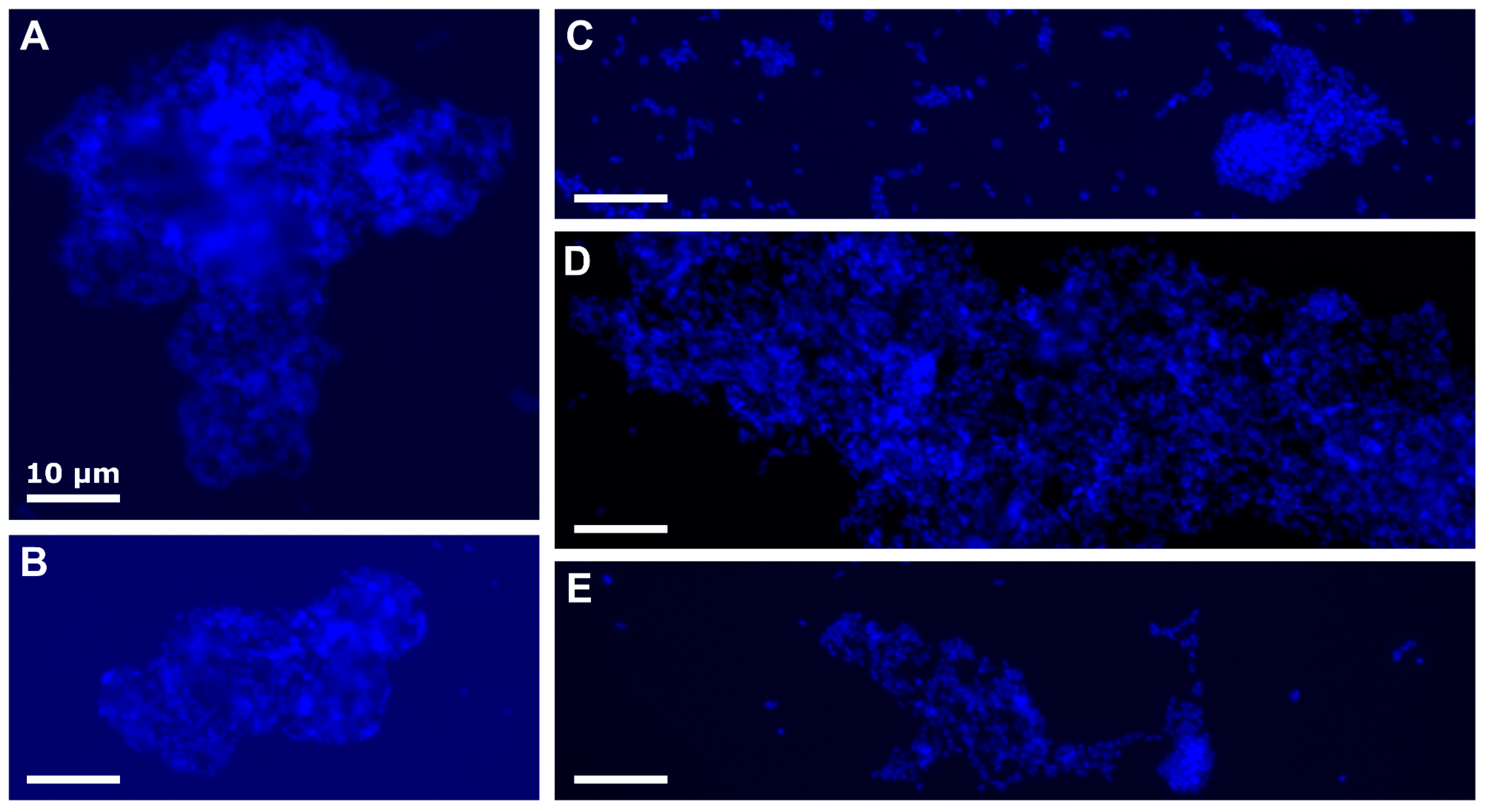
3.3. Potential Mechanisms Responsible for the Observed Effects of Starved Marinobacter sp. TT1
3.4. Environmental Implications
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- US National Commission DWH. The Use of Surface and Subsea Dispersants during the BP Deepwater Horizon Oil Spill; National Commission on the BP Deepwater Horizon Oil Spill and Offshore Drilling: Washington, DC, USA, 2011.
- McNutt, M.K.; Camilli, R.; Crone, T.J.; Guthrie, G.D.; Hsieh, P.A.; Ryerson, T.B.; Savas, O.; Shaffer, F. Review of flow rate estimates of the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20260–20267. [Google Scholar] [CrossRef] [PubMed]
- McGowan, C.J.; Kwok, R.K.; Engel, L.S.; Stenzel, M.R.; Stewart, P.A.; Sandler, D.P. Respiratory, Dermal, and Eye Irritation Symptoms Associated with Corexit EC9527A/EC9500A following the Deepwater Horizon Oil Spill: Findings from the GuLF STUDY. Environ. Health Perspect. 2017, 125, 097015. [Google Scholar] [CrossRef] [PubMed]
- DWH NRDA Trustees. Deepwater Horizon Oil Spill: Final Programmatic Damage Assessment and Restoration Plan and Final Programmatic Environmental Impact Statement; Deepwater Horizon Natural Resource Damage Assessment Trustees: Washington, DC, USA, 2016.
- US NASEM. The Use of Dispersants in Marine Oil Spill Response; National Academies of Sciences, Engineering, and Medicine: Washington, DC, USA, 2020. [Google Scholar]
- Bælum, J.; Borglin, S.; Chakraborty, R.; Fortney, J.L.; Lamendella, R.; Mason, O.U.; Auer, M.; Zemla, M.; Bill, M.; Conrad, M.E. Deep-sea bacteria enriched by oil and dispersant from the Deepwater Horizon spill. Environ. Microbiol. 2012, 14, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Prince, R.; Kelley, B.; Butler, J. Three Widely-Available Dispersants Substantially Increase the Biodegradation of Otherwise Undispersed Oil. J. Marine Sci. Res. Dev. 2016, 6, 183. [Google Scholar] [CrossRef]
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Lee, K.; Greer, C.W. Chemical dispersants enhance the activity of oil- and gas condensate-degrading marine bacteria. ISME J. 2017, 11, 2793–2808. [Google Scholar] [CrossRef]
- Hamdan, L.J.; Fulmer, P.A. Effects of Corexit EC9500A on bacteria from a beach oiled by the Deepwater Horizon spill. Aquat. Microb. Ecol. 2011, 63, 101–109. [Google Scholar] [CrossRef]
- Kleindienst, S.; Seidel, M.; Ziervogel, K.; Grim, S.; Loftis, K.; Harrison, S.; Malkin, S.Y.; Perkins, M.J.; Field, J.; Sogin, M.L. Chemical dispersants can suppress the activity of natural oil-degrading microorganisms. Proc. Natl. Acad. Sci. USA 2015, 112, 14900–14905. [Google Scholar] [CrossRef]
- Rahsepar, S.; Smit, M.P.; Murk, A.J.; Rijnaarts, H.H.; Langenhoff, A.A. Chemical dispersants: Oil biodegradation friend or foe? Mar. Pollut. Bull. 2016, 108, 113–119. [Google Scholar] [CrossRef]
- Hackbusch, S.; Noirungsee, N.; Viamonte, J.; Sun, X.; Bubenheim, P.; Kostka, J.E.; Muller, R.; Liese, A. Influence of pressure and dispersant on oil biodegradation by a newly isolated Rhodococcus strain from deep-sea sediments of the Gulf of Mexico. Mar. Pollut. Bull. 2020, 150, 110683. [Google Scholar] [CrossRef]
- Overholt, W.A.; Marks, K.P.; Romero, I.C.; Hollander, D.J.; Snell, T.W.; Kostka, J.E. Hydrocarbon-degrading bacteria exhibit a species-specific response to dispersed oil while moderating ecotoxicity. Appl. Environ. Microbiol. 2016, 82, 518–527. [Google Scholar] [CrossRef]
- Techtmann, S.M.; Zhuang, M.; Campo, P.; Holder, E.; Elk, M.; Hazen, T.C.; Conmy, R.; Santo Domingo, J.W. Corexit 9500 Enhances Oil Biodegradation and Changes Active Bacterial Community Structure of Oil-Enriched Microcosms. Appl. Environ. Microbiol. 2017, 83, e03416–e03462. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.M.; Whitaker, E.A.; De Pascuale, V.; Wade, T.L.; Knap, A.H.; Santschi, P.H.; Quigg, A.; Sylvan, J.B. Rapid Formation of Microbe-Oil Aggregates and Changes in Community Composition in Coastal Surface Water Following Exposure to Oil and the Dispersant Corexit. Front. Microbiol. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.T.; Moore, M.; Lee, H.; Trevors, J.T. Effect of carbon starvation on p-nitrophenol degradation by a Moraxella strain in buffer and river water. FEMS Microbiol. Ecol. 2005, 51, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Khleifat, K.M. Effect of substrate adaptation, carbon starvation and cell density on the biodegradation of phenol by Actinobacillus sp. Fresenius Environ. Bull. 2007, 16, 726–730. [Google Scholar]
- Santisi, S.; Genovese, M.; Crisafi, F.; Gentile, G.; Volta, A.; Bonsignore, M.; Cappello, S. Effects of Growth Conditions on Hydrophobicity in Marine Obligate Hydrocarbonoclastic Bacteria. Intern. J. Microbiol. Appl. 2015, 2, 50–55. [Google Scholar]
- Putthividhya, A.; Kukor, J.J.; Abriola, L.M. The effects of substrate exposure history and carbon starvation-induced stress on the EPS synthesis of TCE degrading toluene oxidizing soil bacteria. Environ. Earth Sci. 2016, 75. [Google Scholar] [CrossRef]
- Foght, J.; Westlake, D. Effect of the dispersant Corexit 9527 on the microbial degradation of Prudhoe Bay oil. Can. J. Microbiol. 1982, 28, 117–122. [Google Scholar] [CrossRef]
- Lindstrom, J.E.; Braddock, J.F. Biodegradation of petroleum hydrocarbons at low temperature in the presence of the dispersant Corexit 9500. Mar. Pollut. Bull. 2002, 44, 739–747. [Google Scholar] [CrossRef]
- Gutierrez, T.; Singleton, D.R.; Berry, D.; Yang, T.; Aitken, M.D.; Teske, A. Hydrocarbon-degrading bacteria enriched by the Deepwater Horizon oil spill identified by cultivation and DNA-SIP. ISME J. 2013, 7, 2091–2104. [Google Scholar] [CrossRef]
- Duran, R. Marinobacter. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Mounier, J.; Camus, A.; Mitteau, I.; Vaysse, P.J.; Goulas, P.; Grimaud, R.; Sivadon, P. The marine bacterium Marinobacter hydrocarbonoclasticus SP17 degrades a wide range of lipids and hydrocarbons through the formation of oleolytic biofilms with distinct gene expression profiles. FEMS Microbiol. Ecol. 2014, 90, 816–831. [Google Scholar] [CrossRef]
- Dyksterhouse, S.E.; Gray, J.P.; Herwig, R.P.; Lara, J.C.; Staley, J.T. Cycloclasticus pugetii gen. nov., sp. nov., an Aromatic Hydrocarbon-Degrading Bacterium from Marine Sediments. Int. J. Syst. Bacteriol. 1995, 45, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of Complex Microbial Populations by Denaturing Gradient Gel Electrophoresis Analysis of Polymerase Chain Reaction Amplified Genes Coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- ExxonMobil. Oil Spill Response Field Manual; ExxonMobil: Owen, TX, USA, 2014. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Straškrabová, V. The effect of substrate shock on populations of starving aquatic bacteria. J. Appl. Bacteriol. 1983, 54, 217–224. [Google Scholar] [CrossRef]
- Sebastian, M.; Estrany, M.; Ruiz-Gonzalez, C.; Forn, I.; Sala, M.M.; Gasol, J.M.; Marrase, C. High Growth Potential of Long-Term Starved Deep Ocean Opportunistic Heterotrophic Bacteria. Front. Microbiol. 2019, 10, 760. [Google Scholar] [CrossRef] [PubMed]
- Singer, E.; Webb, E.A.; Nelson, W.C.; Heidelberg, J.F.; Ivanova, N.; Pati, A.; Edwards, K.J. Genomic potential of Marinobacter aquaeolei, a biogeochemical “opportunitroph”. Appl. Environ. Microbiol. 2011, 77, 2763–2771. [Google Scholar] [CrossRef]
- Kleindienst, S.; Grim, S.; Sogin, M.; Bracco, A.; Crespo-Medina, M.; Joye, S.B. Diverse, rare microbial taxa responded to the Deepwater Horizon deep-sea hydrocarbon plume. ISME J 2016, 10, 400–415. [Google Scholar] [CrossRef]
- Sun, X.; Kostka, J.E. Hydrocarbon-Degrading Microbial Communities Are Site Specific, and Their Activity Is Limited by Synergies in Temperature and Nutrient Availability in Surface Ocean Waters. Appl. Environ. Microbiol. 2019, 85, e00419–e00443. [Google Scholar] [CrossRef]
- Chakraborty, R.; Borglin, S.E.; Dubinsky, E.A.; Andersen, G.L.; Hazen, T.C. Microbial Response to the MC-252 Oil and Corexit 9500 in the Gulf of Mexico. Front. Microbiol. 2012, 3, 357. [Google Scholar] [CrossRef]
- Yam, E.M.; Tang, K.W. Effects of starvation on aggregate colonization and motility of marine bacteria. Aquat. Microb. Ecol. 2007, 48, 207–215. [Google Scholar] [CrossRef][Green Version]
- Kjelleberg, S.; Hermansson, M. Starvation-induced effects on bacterial surface characteristics. Appl. Environ. Microbiol. 1984, 48, 497–503. [Google Scholar] [CrossRef]
- Joye, S.; Kleindienst, S.; Pena-Montenegro, T.D. SnapShot: Microbial Hydrocarbon Bioremediation. Cell 2018, 172, 1336. [Google Scholar] [CrossRef] [PubMed]
- Bergkessel, M.; Basta, D.W.; Newman, D.K. The physiology of growth arrest: Uniting molecular and environmental microbiology. Nat. Rev. Microbiol. 2016, 14, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Fida, T.T.; Moreno-Forero, S.K.; Heipieper, H.J.; Springael, D. Physiology and transcriptome of the polycyclic aromatic hydrocarbon-degrading Sphingomonas sp. LH128 after long-term starvation. Microbiology 2013, 159, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Kaberdin, V.R.; Montanchez, I.; Parada, C.; Orruno, M.; Arana, I.; Barcina, I. Unveiling the Metabolic Pathways Associated with the Adaptive Reduction of Cell Size During Vibrio harveyi Persistence in Seawater Microcosms. Microb. Ecol. 2015, 70, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Mounier, J.; Hakil, F.; Branchu, P.; Naïtali, M.; Goulas, P.; Sivadon, P.; Grimaud, R. AupA and AupB Are Outer and Inner Membrane Proteins Involved in Alkane Uptake in Marinobacter hydrocarbonoclasticus SP17. mBio 2018, 9, e00518–e00520. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F. Degradation of alkanes by bacteria. Environ. Microbiol. 2009, 11, 2477–2490. [Google Scholar] [CrossRef]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef]
- Folt, C.L.; Chen, C.Y.; Moore, M.V.; Burnaford, J. Synergism and antagonism among multiple stressors. Limnol. Oceanogr. 1999, 44, 864–877. [Google Scholar] [CrossRef]
- Ozhan, K.; Bargu, S. Distinct responses of Gulf of Mexico phytoplankton communities to crude oil and the dispersant Corexit EC9500A under different nutrient regimes. Ecotoxicology 2014, 23, 370–384. [Google Scholar] [CrossRef]
- Signorini, S.R.; Franz, B.A.; McClain, C.R. Chlorophyll variability in the oligotrophic gyres: Mechanisms, seasonality and trends. Front. Mar. Sci. 2015, 2, 1–11. [Google Scholar] [CrossRef]
- Polovina, J.J.; Howell, E.A.; Abecassis, M. Ocean’s least productive waters are expanding. Geophys. Res. Lett. 2008, 35, L03618. [Google Scholar] [CrossRef]
- Vaysse, P.J.; Sivadon, P.; Goulas, P.; Grimaud, R. Cells dispersed from Marinobacter hydrocarbonoclasticus SP17 biofilm exhibit a specific protein profile associated with a higher ability to reinitiate biofilm development at the hexadecane–Water interface. Environ. Microbiol. 2011, 13, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.; Grossi, V.; Bouriat, P.; Goulas, P.; Grimaud, R. Cytoplasmic wax ester accumulation during biofilm-driven substrate assimilation at the alkane–water interface by Marinobacter hydrocarbonoclasticus SP17. Res. Microbiol. 2008, 159, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Rughöft, S.; Jehmlich, N.; Gutierrez, T.; Kleindienst, S. Comparative proteomics of Marinobacter sp. TT1 reveals Corexit impacts on hydrocarbon metabolism, chemotactic motility and biofilm formation. Microorganisms 2020. under review. [Google Scholar]
- Joye, S.B.; Bracco, A.; Özgökmen, T.M.; Chanton, J.P.; Grosell, M.; MacDonald, I.R.; Cordes, E.E.; Montoya, J.P.; Passow, U. The Gulf of Mexico ecosystem, six years after the Macondo oil well blowout. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 129, 4–19. [Google Scholar] [CrossRef]
- Passow, U.; Ziervogel, K. Marine snow sedimented oil released during the Deepwater Horizon spill. Oceanography 2016, 29, 118–125. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rughöft, S.; Vogel, A.L.; Joye, S.B.; Gutierrez, T.; Kleindienst, S. Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter sp. TT1 by a Chemical Dispersant. J. Mar. Sci. Eng. 2020, 8, 925. https://doi.org/10.3390/jmse8110925
Rughöft S, Vogel AL, Joye SB, Gutierrez T, Kleindienst S. Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter sp. TT1 by a Chemical Dispersant. Journal of Marine Science and Engineering. 2020; 8(11):925. https://doi.org/10.3390/jmse8110925
Chicago/Turabian StyleRughöft, Saskia, Anjela L. Vogel, Samantha B. Joye, Tony Gutierrez, and Sara Kleindienst. 2020. "Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter sp. TT1 by a Chemical Dispersant" Journal of Marine Science and Engineering 8, no. 11: 925. https://doi.org/10.3390/jmse8110925
APA StyleRughöft, S., Vogel, A. L., Joye, S. B., Gutierrez, T., & Kleindienst, S. (2020). Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter sp. TT1 by a Chemical Dispersant. Journal of Marine Science and Engineering, 8(11), 925. https://doi.org/10.3390/jmse8110925